A Rice Immunophilin Gene, OsFKBP16-3, Confers Tolerance to Environmental Stress in Arabidopsis and Rice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
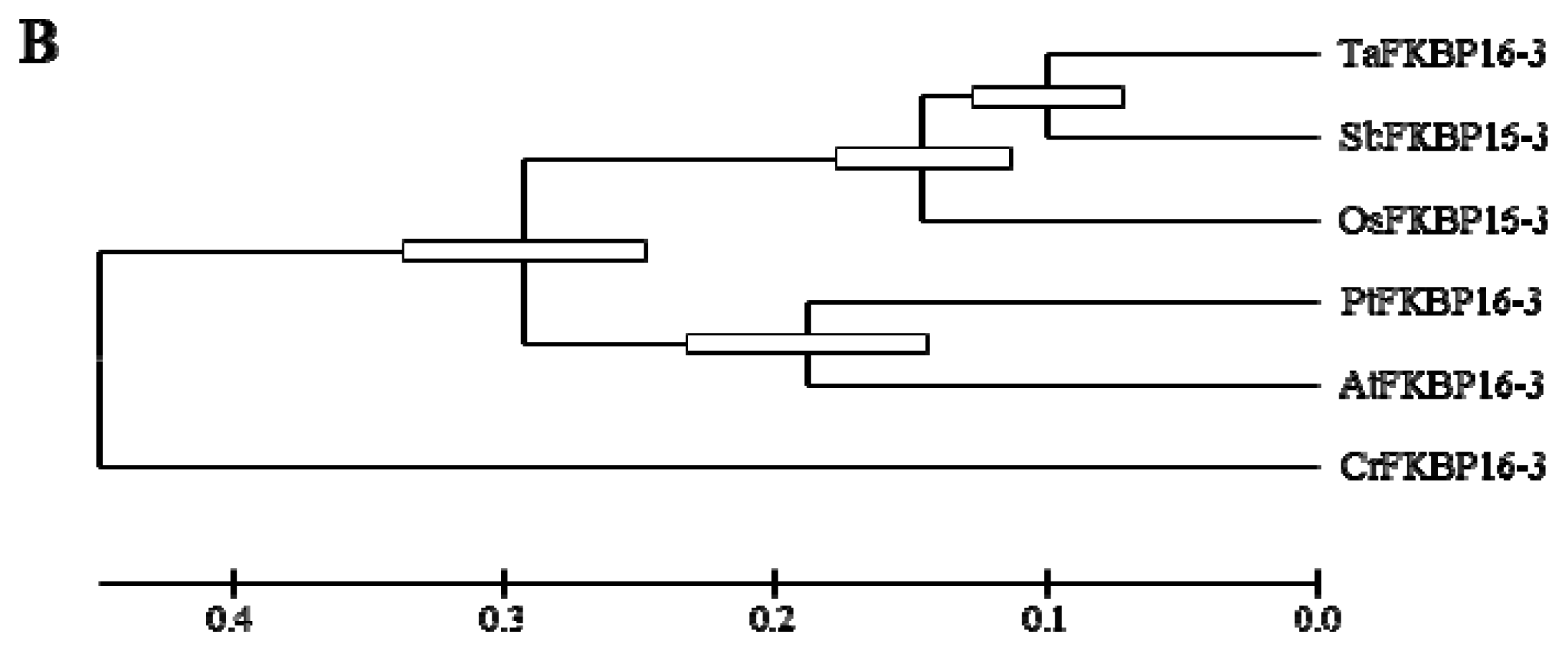
2.1. FKBP16-3s Are Well Conserved in Photosynthetic Organisms
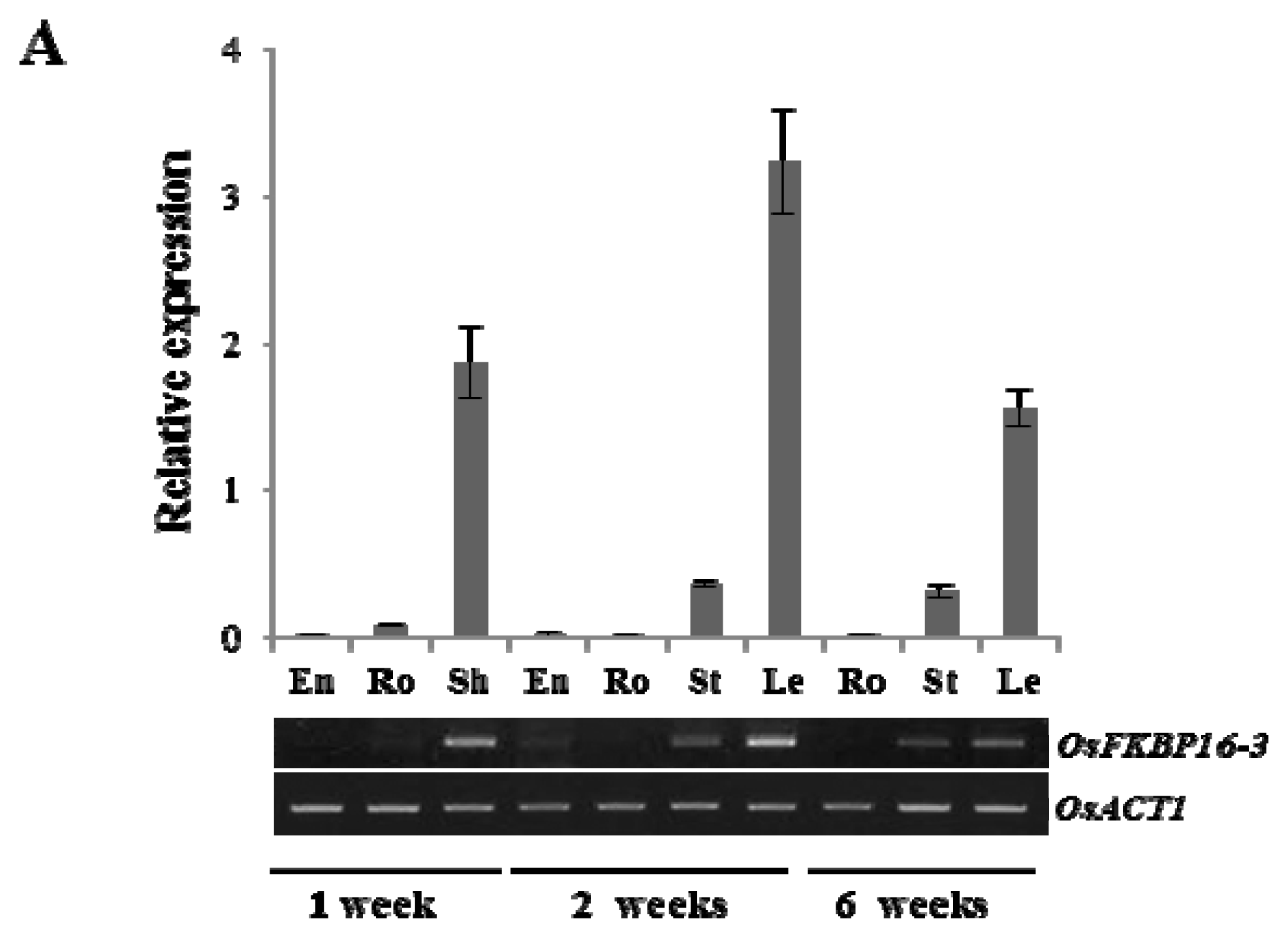
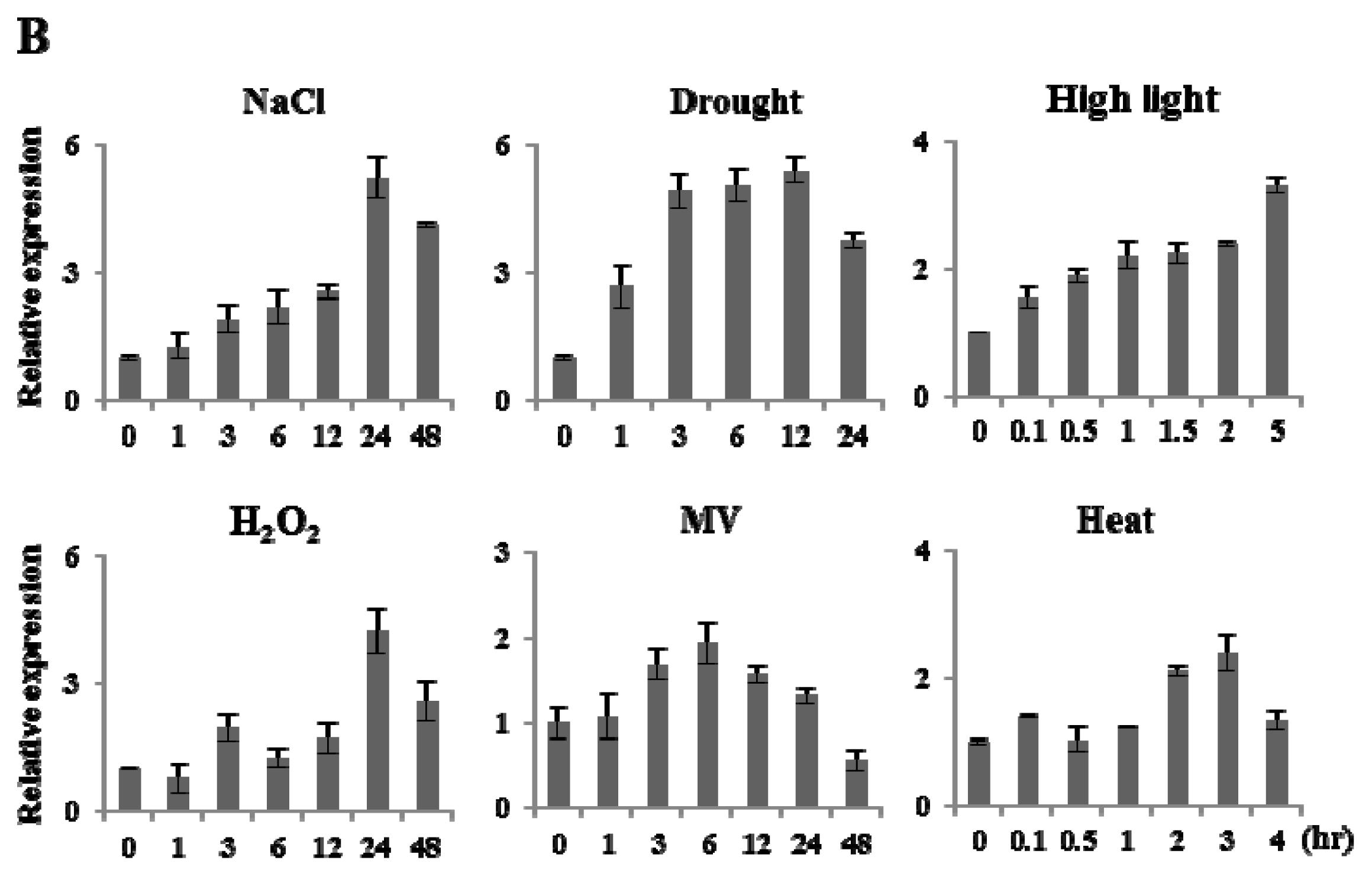
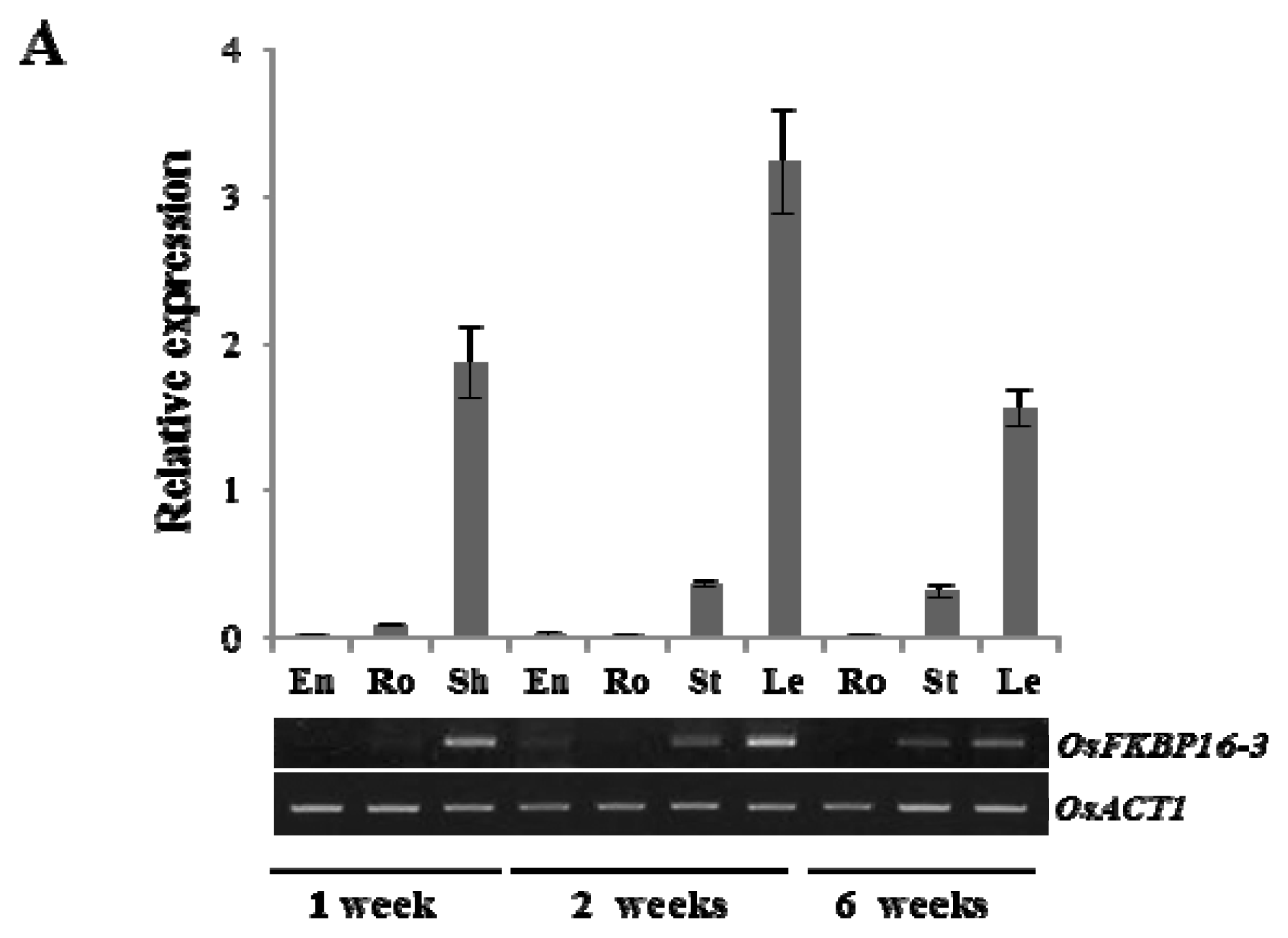
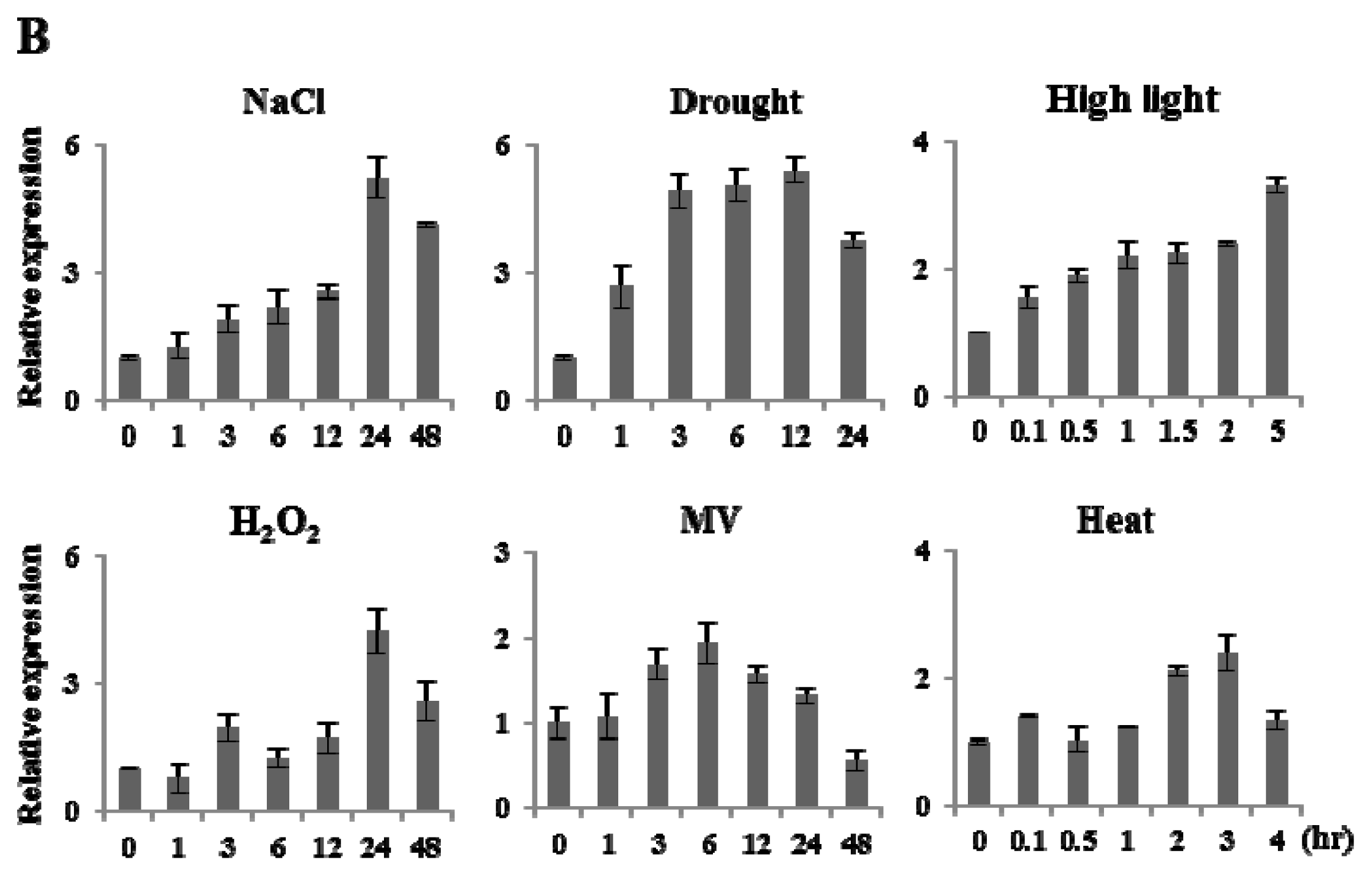
2.2. Expression Analysis of OsFKBP16-3
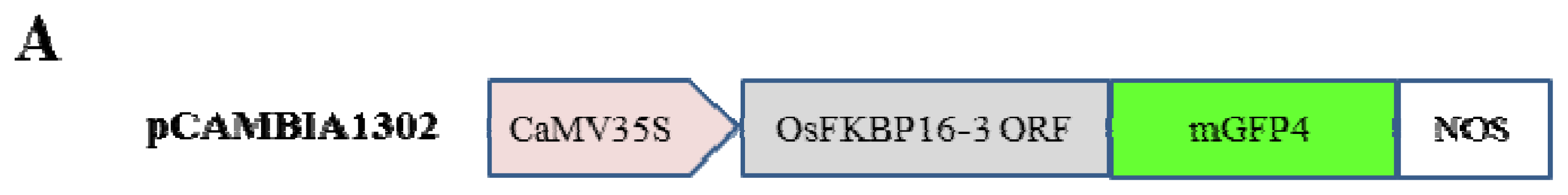
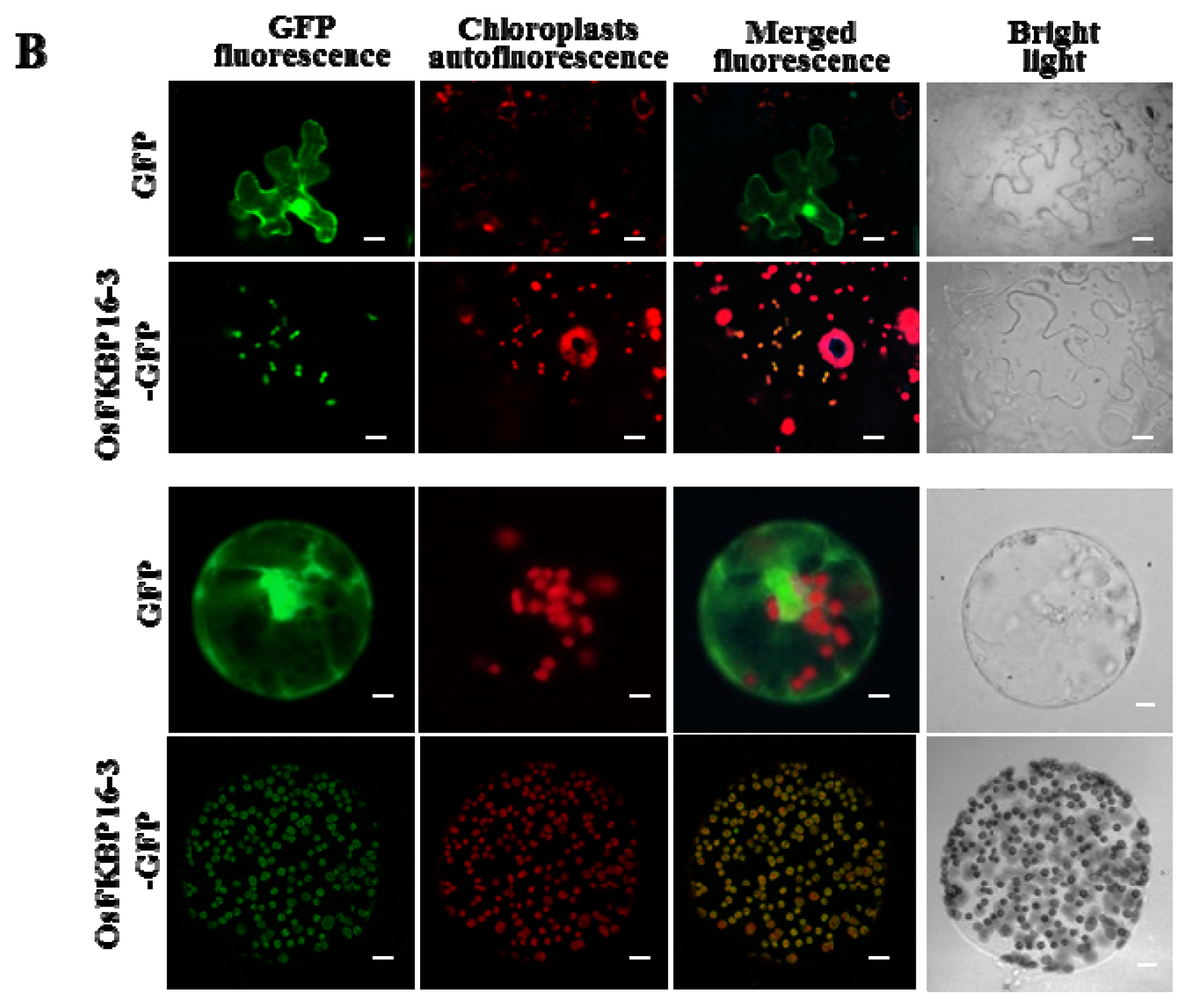
2.3. OsFKBP16-3 Is Localized to the Chloroplasts in Plant Cells
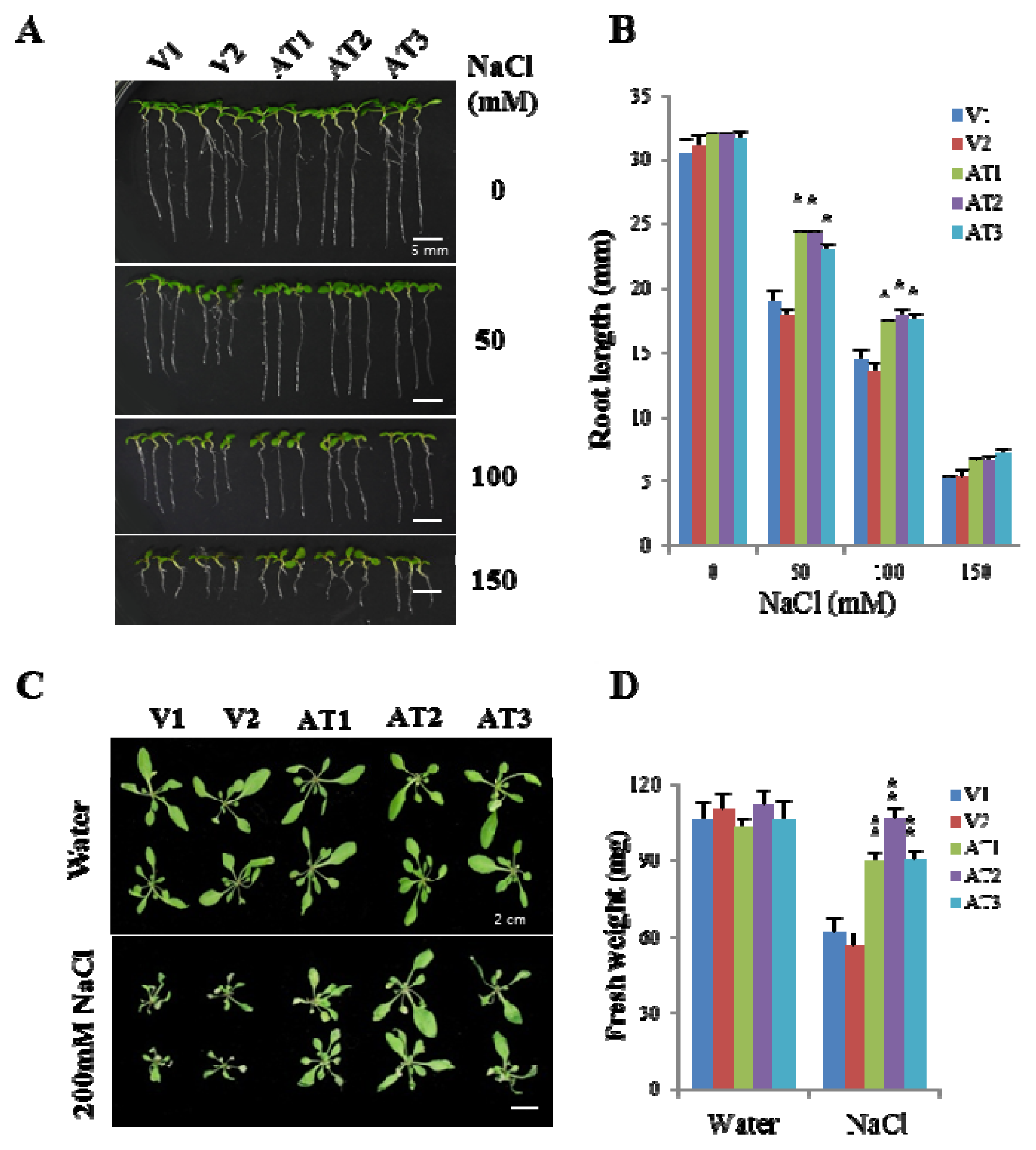
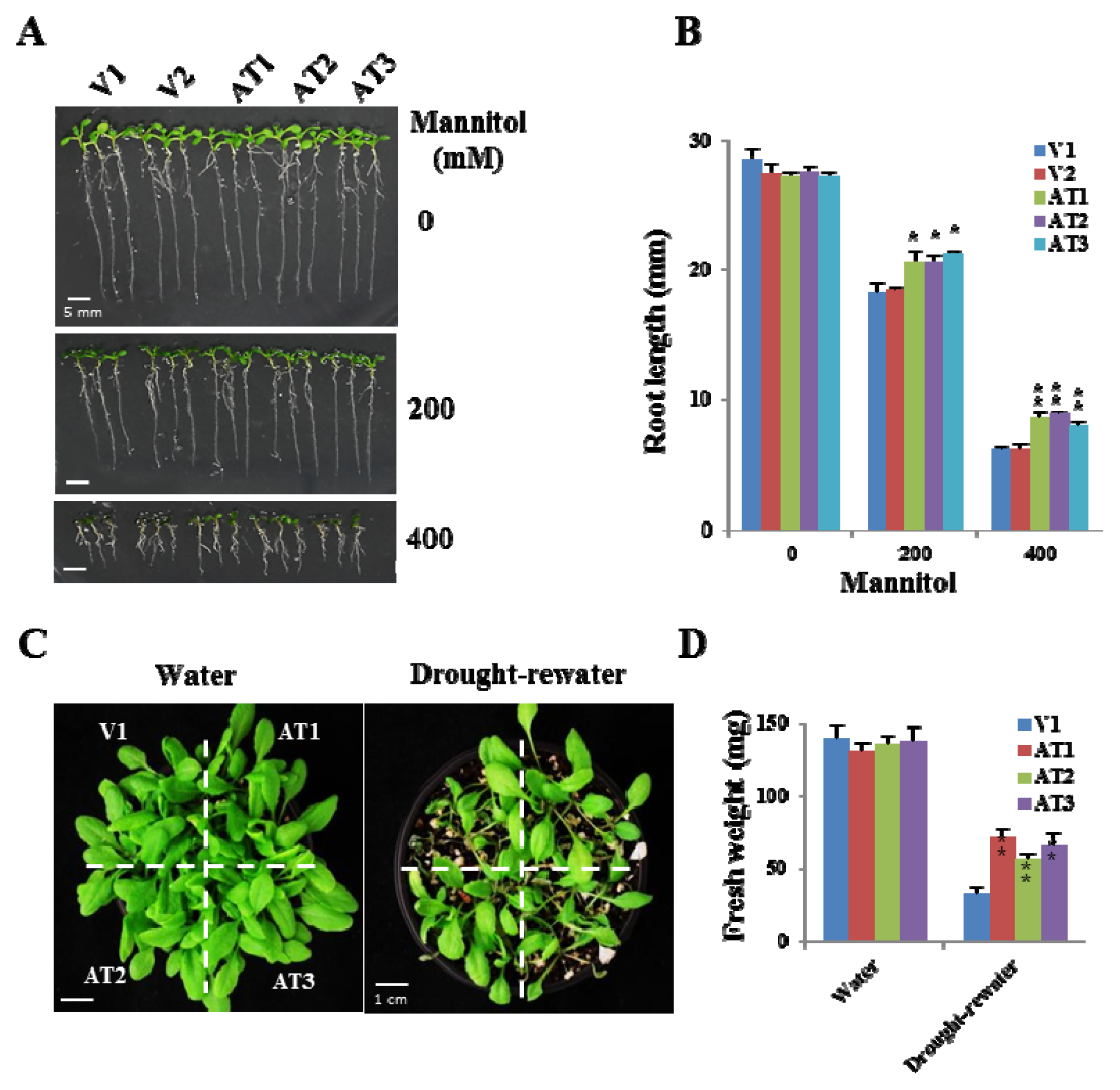
2.4. Ectopic Expression of OsFKBP16-3 Increased Tolerance to Salinity in Arabidopsis
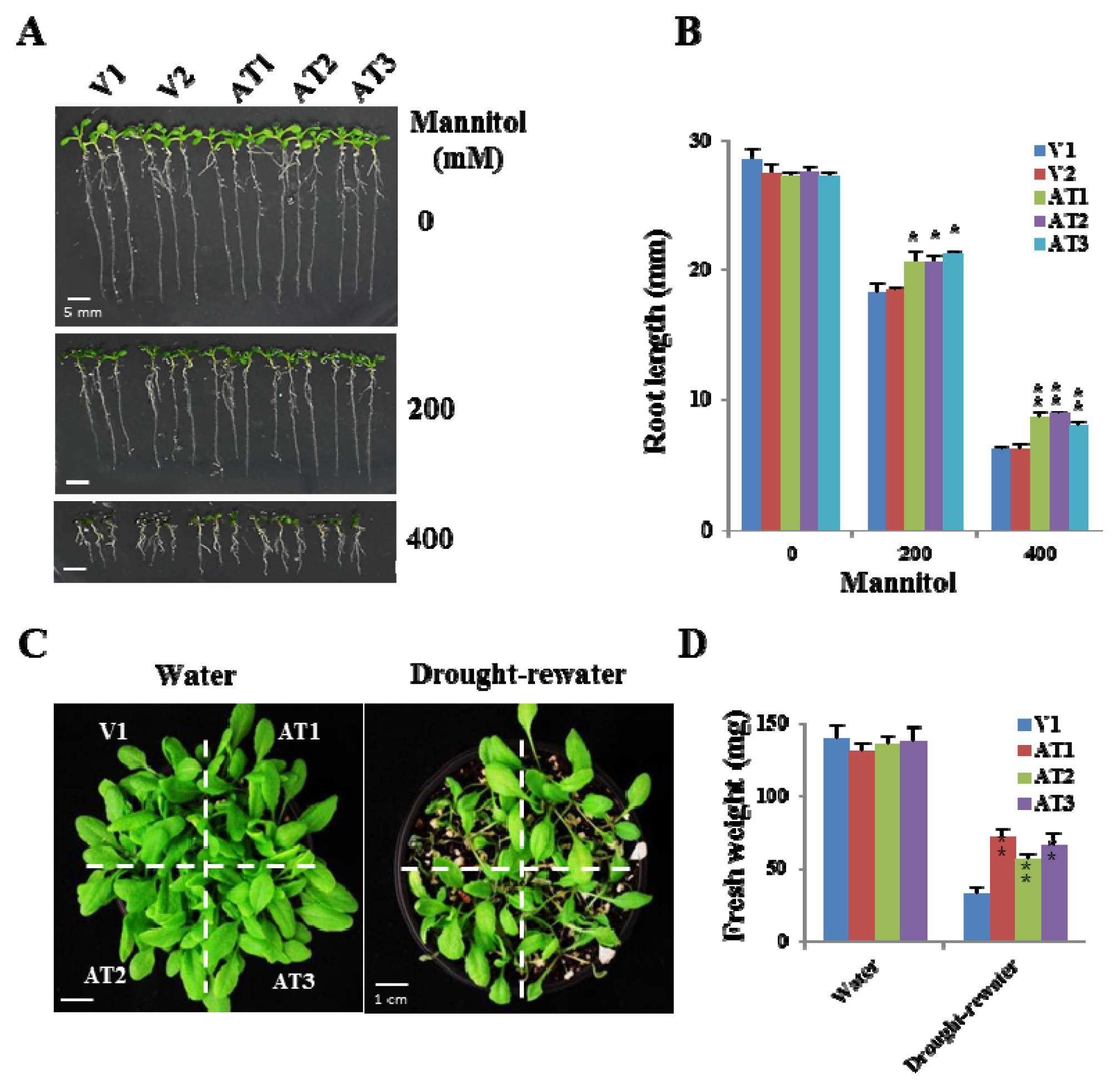
2.5. Ectopic Expression of OsFKBP16-3 Showed Increased Tolerance to Drought Stress in Arabidopsis
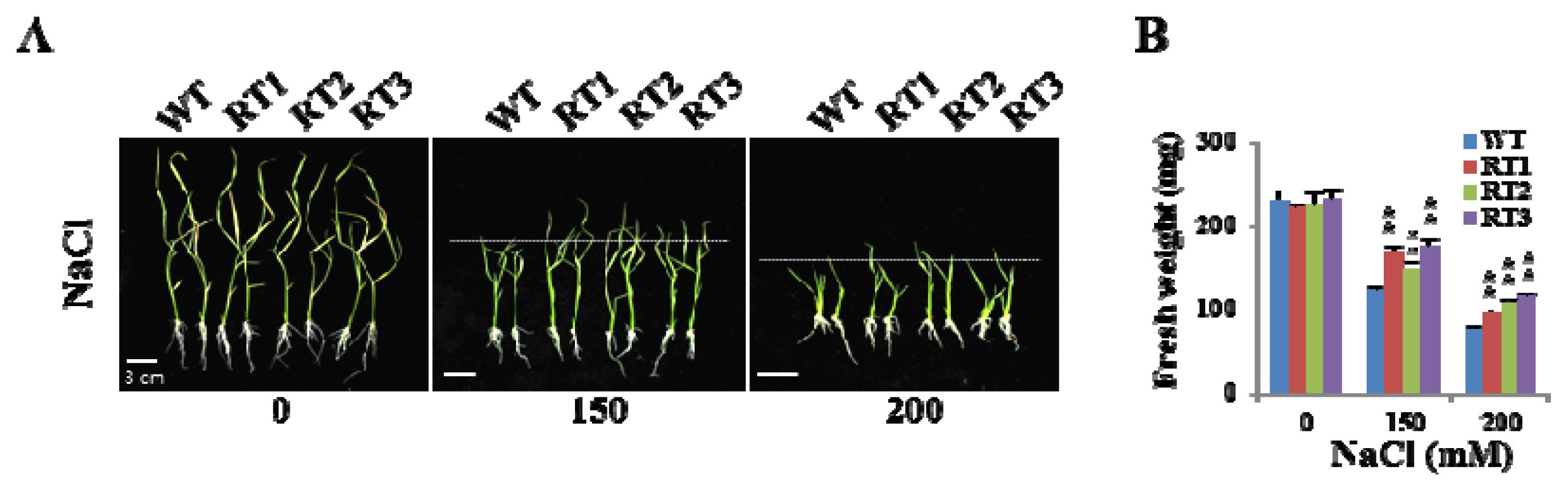
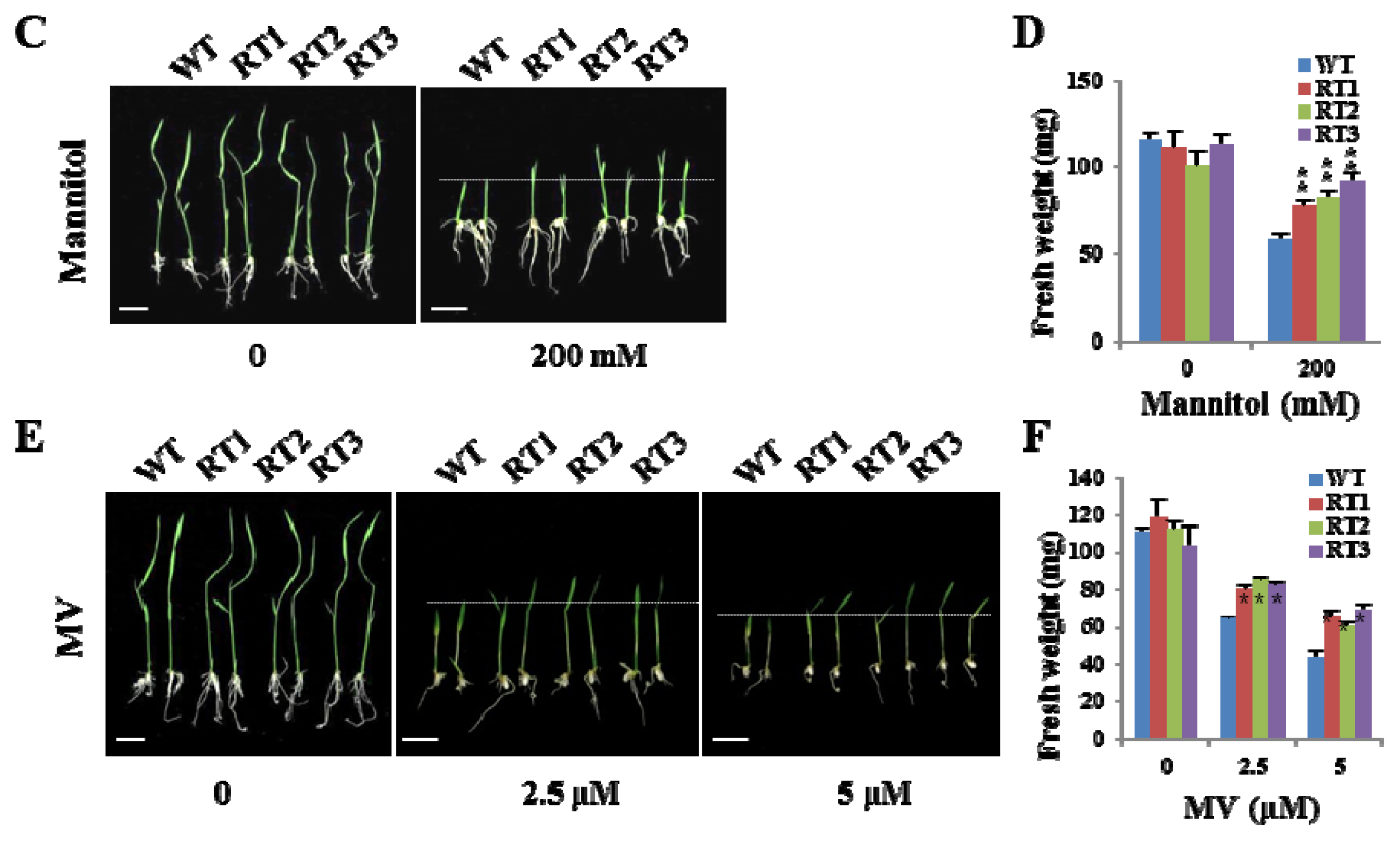
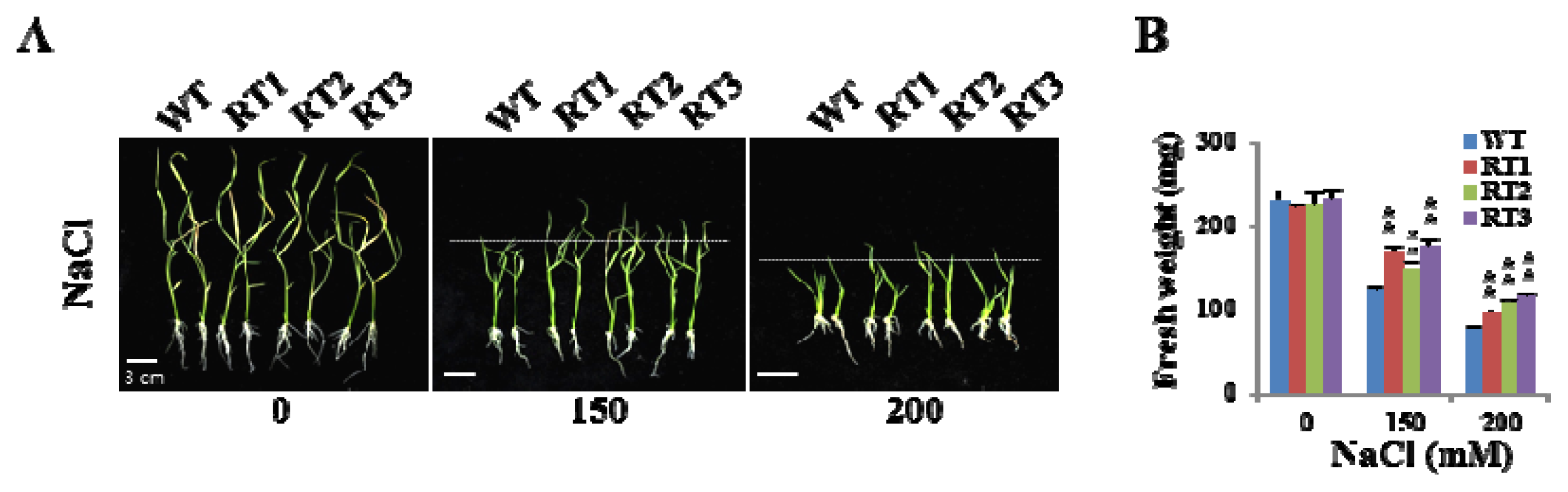
2.6. Overexpression of OsFKBP16-3 Improved Tolerance to Environmental Stress in Rice
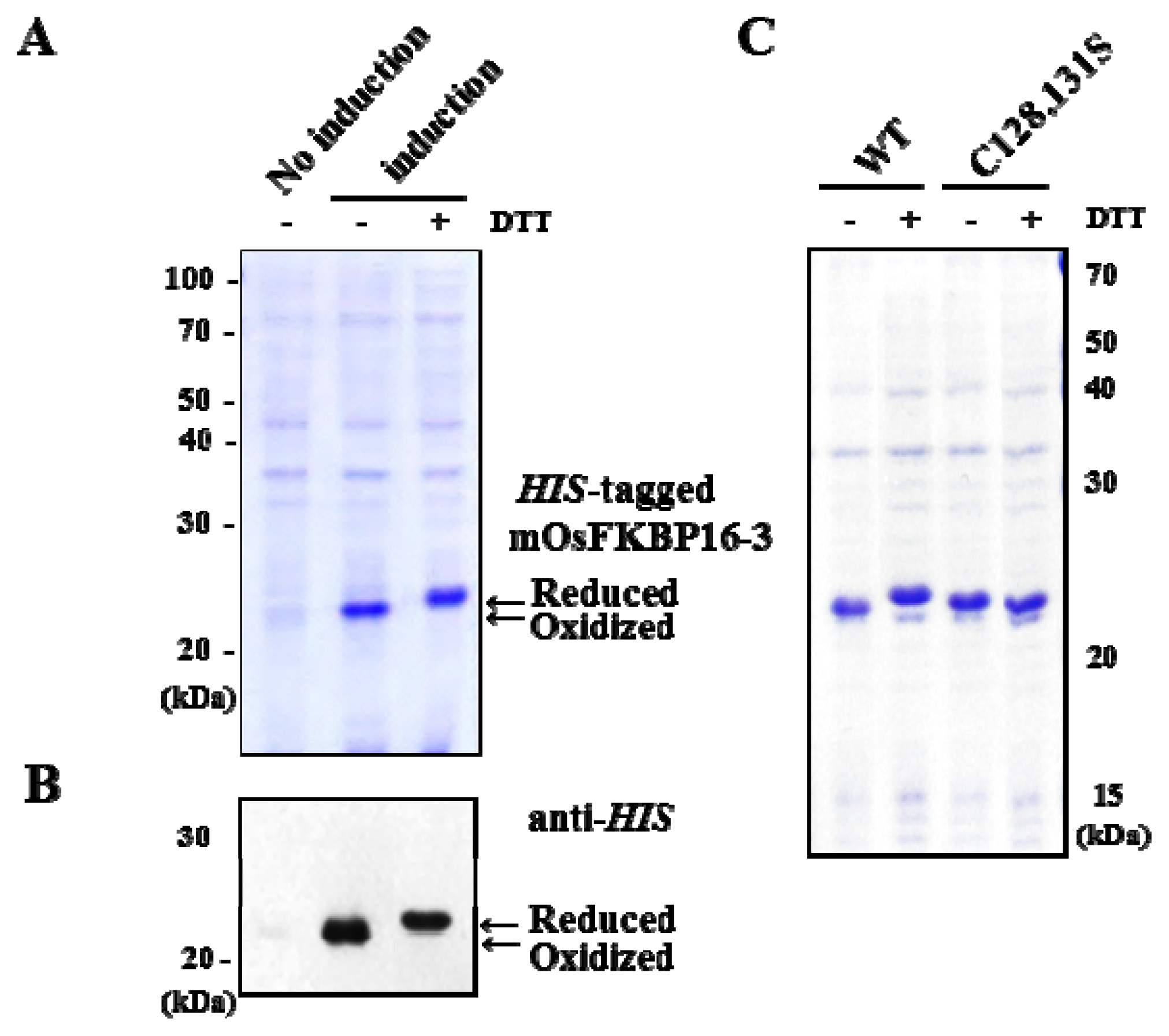
2.7. Redox State of Recombinant OsFKBP16-3 Protein
3. Discussion
4. Experimental Section
4.1. Plant Growth Conditions and Treatments
4.2. Gene Expression Analysis
4.3. Localization of OsFKBP16-3
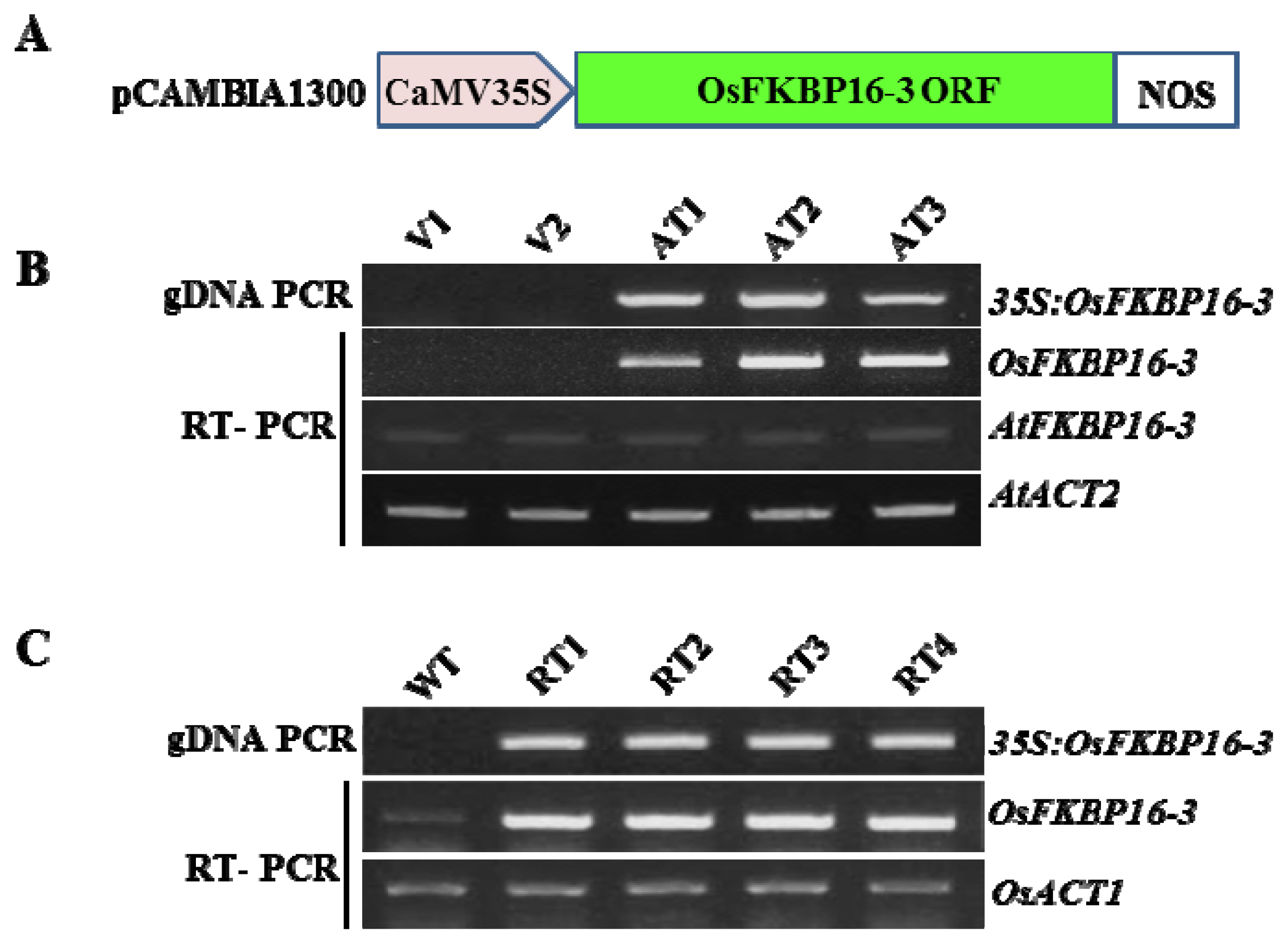
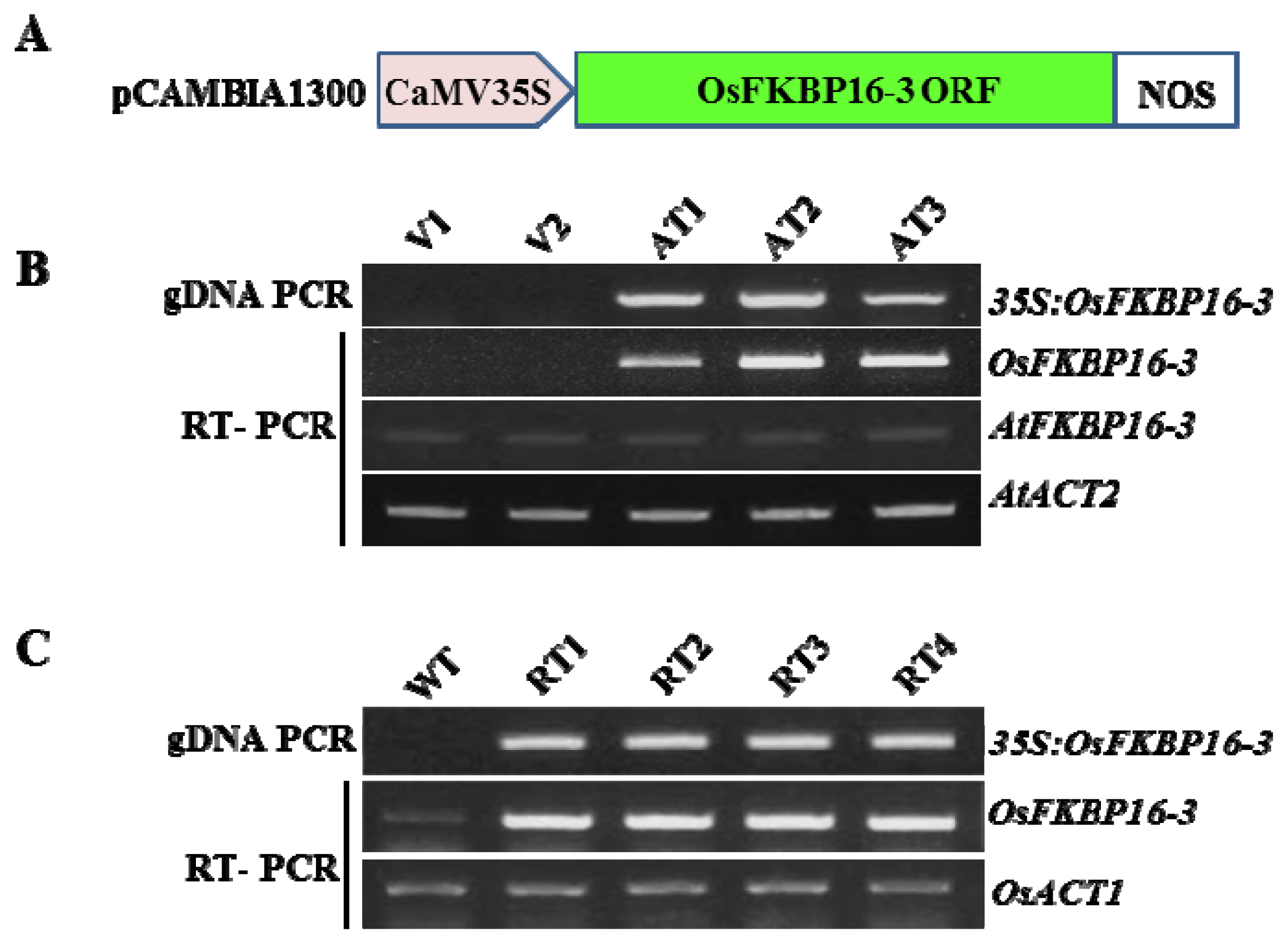
4.4. Gene Constructs and Plant Transformation
4.5. Molecular Analysis of Transgenic Plants
4.6. Stress Treatment of Transgenic Plants
4.7. Protein Expression and Reduction of OsFKBP16-3 in E. coli
5. Conclusions
Supplementary Information
ijms-14-05899-s001.pdfAcknowledgments
Conflict of Interest
References
- Schreiber, S.L. Chemistry and biology of the immuophilins and their immuno-suppressive ligands. Science 1991, 251, 283–287. [Google Scholar]
- Lang, K.; Schmid, F.X.; Fischer, G. Catalysis of protein folding by prolryl isomerase. Nature 1987, 329, 268–270. [Google Scholar]
- Edward, J.G.; Wysocka, J. Flipping MLL1’s switch one proline at a time. Cell 2010, 141, 1108–1110. [Google Scholar]
- Wang, Z.; Song, J.; Milne, T.A.; Wang, G.G.; Li, H.; Allis, C.D.; Patel, D.J. Pro isomerization in MLL1 PHD3-bromo cassette connects H3K4me readout to CyP33 and HDAC-mediated repression. Cell 2010, 25, 1183–1194. [Google Scholar]
- Galat, A. Peptidylprolyl cis/trans isomerase (immunophilins): Biological diversity-targets-functions. Curr. Top. Med. Chem 2003, 3, 1315–1347. [Google Scholar]
- He, Z.; Li, L.; Luan, S. Immunophilins and parvulins. Superfamily of peptidyl prolyl isomerases in Arabidopsis. Plant Physiol 2004, 134, 1248–1267. [Google Scholar]
- Ahn, J.C.; Kim, D.W.; You, Y.N.; Seok, M.S.; Park, J.M.; Hwang, H.; Kim, B.-G.; Luam, S.; Park, H-S.; Cho, H.S. Classification of rice (Oryza. sativa L. japonica Nipponbare) immunophilins (FKBPs, CYPs) and expression patterns under water stress. BMC Plant Biol. 2010, 10, 253. [Google Scholar]
- Kieselbach, T.; Schröder, W.P. The proteome of the chloroplast lumen of higher plants. Photosynth. Res 2003, 787, 249–264. [Google Scholar]
- Peltier, J.B.; Emanuelsson, O.; Kalume, D.E.; Ytterber, J.; Friso, G.; Rudella, A.; Liberles, D.A.; Söderberg, L.; Roepstorff, P.; von Heijne, G.; et al. Central functions of the luminal and peripheral thylakoid proteome of Arabidopsis determined by experimentation and genome-wide prediction. Plant Cell 2002, 14, 211–236. [Google Scholar]
- Pottosin, I.I.; Schönknecht, G. Patch clamp study of the voltage-dependent anion channel in the thylakoid membrane. J. Membr. Biol 1995, 148, 143–156. [Google Scholar]
- Pottosin, I.I.; Schönknecht, G. Ion channel permeable for divalent and monovalent cations in native spinach thylakoid membranes. J. Membr. Biol 1996, 152, 223–233. [Google Scholar]
- Gupta, G.; Mould, R.M.; He, Z.; Luan, S. A chloroplast FKBP interacts with and affects the accumulation of Rieske subunit of cytochrome bf complex. Proc. Natl. Acad. Sci. USA 2002, 99, 15806–15811. [Google Scholar]
- Gopalan, G.; He, Z.; Balmer, Y.; Romano, P.G.N.; Gupta, R.; Héroux, A.; Buchanan, B.B.; Swaminathan, K.; Luan, S. Structural analysis uncovers a role for redox in regulating FKBP13, an immunophilin of the chloroplast thylakoid lumen. Proc. Natl. Acad. Sci. USA 2004, 101, 13945–13950. [Google Scholar]
- Lima, A.; Lima, S.; Wong, J.H.; Phillips, R.S.; Buchanan, B.B.; Luan, S. A redox-active FKBP-type immunophilin functions in accumulation of the photosystem II supercomplex in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2006, 103, 12631–12636. [Google Scholar]
- Fu, A.; He, Z.; Cho, H.S.; Lima, A.; Buchanan, B.B.; Luan, S. A chloroplast cyclophilin functions in the assembly and maintenance of photosystem II in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2006, 103, 12631–1263. [Google Scholar]
- Sirpiö, S.; Khrouchtchova, A.; Allahverdiyeva, Y.; Hansson, M.; Fristedt, R.; Vener, A.V.; Scheller, H.V.; Jensen, P.E.; Haldrup, A.; Aro, E.M. AtCYP38 ensures early biogenesis, correct assembly and sustenance of photosystem II. Plant J 2008, 55, 639–651. [Google Scholar]
- Vasudevan, D.; Fu, A.; Luan, S.; Swaminathan, K. Crystal structure of Arabidopsis cyclophilin38 reveals a previously uncharacterized immunophilin fold and a possible autoinhibitory mechanism. Plant Cell 2012, 24, 2666–2674. [Google Scholar]
- Edvardsson, A.; Shapiquzov, A.; Petersson, U.A.; Schröder, W.P.; Vener, A.V. Immunophilin AtFKBP13 sustains all peptidyl-proryl isomerase activity in the thylakoid lumen from Arabidopsis thaliana deficient in AtCYP20–2. Biochemistry 2007, 46, 9432–9442. [Google Scholar]
- Romano, P.G.N.; Horton, P.; Gray, J.E. The Arabidopsis cyclophilin gene family. Plant Physiol 2004, 134, 1268–1282. [Google Scholar]
- Romano, P.G.N.; Gray, J.E.; Horton, P.; Luan, S. Plant immunophilins: Functional versatility beyond protein maturation. New Phytol 2005, 166, 753–769. [Google Scholar]
- Sirpiö, S.; Holmström, M.; Battchikova, N.; Aro, E.M. AtCYP20–2 is an auxiliary protein of the chloroplast NAD(P)H dehydrogenase complex. FEBS Lett 2009, 583, 2355–2358. [Google Scholar]
- Kim, S.K.; You, Y.N.; Park, J.C.; Joung, Y.; Kim, B.G.; Ahn, J.C.; Cho, H.S. The rice thylakoid lumenal cyclophilin OsCYP20–2 confers enhanced environmental stress tolerance in tobacco and Arabidopsis. Plant Cell. Rep 2012, 31, 417–426. [Google Scholar]
- Li, B.; Xu, Y.; Zhang, Y.; Wang, T.; Bai, Y.; Han, C.; Zhang, A.; Xu, Z.; Chong, K. Integrative study on proteomics, molecular physiology, and genetics reveals an accumulation of cyclophilin-like protein, TaCYP20–2, leading to an increase of Rht protein and dwarf in a novel GA-insensitive mutant (gaid) in Wheat. J. Proteome Res 2010, 9, 4242–4253. [Google Scholar]
- Peng, L.; Fukao, Y.; Fujiwara, M.; Taakami, T.; Shikanai, T. Efficient operation of NAD(P)H dehydrogenase requires supercomplex formation with photosystem I via minor LHCI in Arabidopsis. Plant Cell 2009, 21, 3623–3640. [Google Scholar]
- Gollan, P.J.; Ziemann, M.; Bhave, M. PPIase activities and interaction partners of FK506-binding proteins in the wheat thylakoid. Physiol. Plant 2011, 143, 385–395. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol 2004, 55, 373–399. [Google Scholar]
- Martin, J.L. Thioredoxin: A fold for all reasons. Structure 1995, 3, 245–250. [Google Scholar]
- Marchand, C.; Le Marèchal, P.; Meyer, Y.; Miginiac-Maslow, M.; Issakidis-Bourguet, E.; Decottignies, P. New targets of Arabidopsis thioredoxins revealed by proteomic analysis. Proteomics 2004, 4, 2696–2706. [Google Scholar]
- Michelet, L.; Zaffagnini, M.; Vanacker, H.; Le Marèchal, P.; Marchand, C.; Schroda, M.; Lemmaire, S.D.; Decottignies, P. In vivo targets of S-thiolation in Chlamydomonas reinhardtii. J. Biol. Chem 2008, 283, 21571–21578. [Google Scholar]
- Nicholas, K.B.; Nicholas, H.B., Jr; Deerfield, D.W. Genedoc: Analysis and visualization of genetic variation. Embnet. News 1997, 4, 1–4. [Google Scholar]
- DeCenzo, M.T.; Park, S.T.; Jarrett, B.P.; Aldape, R.A.; Futer, O.; Murck, M.A.; Livingston, D.J. FK-506-binding protein mutational analysis: Defining the active-site residue contributions to catalysis and the stability of ligand complexes. Protein Eng 1996, 9, 173–180. [Google Scholar]
- Izawa, T.; Shimamoto, K. Becoming a model plant: The importance of rice to plant science. Trends Plant Sci 1996, 1, 95–99. [Google Scholar]
- Jackson, K.; Söll, D. Mutations in a new Arabidopsis cyclophilin disrupt its interaction with protein phosphatase 2A. Mol. Gen. Genet 1999, 262, 830–838. [Google Scholar]
- Robinson, C.; Bolhuis, A. Protein targeting by the twin-arginine translocation pathway. Nat. Rev. Mol. Cell Biol 2001, 2, 35–356. [Google Scholar]
- Edvardsson, A.; Eshaghi, S.; Vener, A.V.; Andersson, B. The major peptidyl-prolyl isomerase activity in thylakoid lumen of plant chloroplast belongs to a novel cyclophilin TLP20. FEBS Lett 2003, 542, 137–141. [Google Scholar]
- Finkel, T.; Holbrook, N.J. Oxidants, oxidative stress and the biology of ageing. Nature 2000, 408, 239–247. [Google Scholar]
- Kanman, K.; Jain, S. Oxidative stress and apotosis. Pathophysiology 2000, 7, 153–163. [Google Scholar]
- Halliwell, B. Reactive species and antioxidants. Redox biology is a fundamental theme of aerobic life. Plant Physiol 2006, 141, 312–322. [Google Scholar]
- Pfannschmidt, T. Chloroplast redox signals: How photosynthesis controls its own genes. Trends Plant Sci 2003, 8, 33–41. [Google Scholar]
- Notor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol 1998, 49, 249–279. [Google Scholar]
- Schürmann, P. Redox signaling in the chloroplast: The Ferredoxin/Thioredoxin system. Antioxid. Redox. Signal 2003, 5, 69–78. [Google Scholar]
- Kim, J.; Eichacker, L.A.; Rudiger, W.; Mullet, J.E. Chlorophyll regulates accumulation of the plastid-encoded chlorophyll proteins P700 and D1 by increasing apoprotein stability. Plant Physiol 1994, 104, 907–916. [Google Scholar]
- Lezhneva, L.; Amann, K.; Meurer, J. The universally conserved HCF101 protein is involved in assembly of [4Fe-4S]-cluster-containing complexes in Arabidopsis thaliana chloroplasts. Plant J 2004, 37, 174–185. [Google Scholar]
- Amann, K.; Lezhneva, L.; Wanner, G.; Hermann, R.G.; Meurer, J. Accumulation of photosystem one1, a member of a novel gene family, is required for accumulation of [4Fe-4S] Cluster–containing chloroplast complexes and antenna proteins. Plant Cell 2004, 16, 3084–3097. [Google Scholar]
- Wang, Q.; Sullivan, R.W.; Kight, A.; Henry, R.L.; Huang, J.; Jones, A.M.; Korth, K.L. Deletion of the chloroplast-localized thylakoid formation1 gene product in Arabidopsis leads to deficient thylakoid formation and variegated leaves. Plant Physiol 2004, 136, 3594–3604. [Google Scholar]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza. sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J 1994, 6, 271–282. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 1998, 16, 735–743. [Google Scholar]





© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Park, H.J.; Lee, S.S.; You, Y.N.; Yoon, D.H.; Kim, B.-G.; Ahn, J.C.; Cho, H.S. A Rice Immunophilin Gene, OsFKBP16-3, Confers Tolerance to Environmental Stress in Arabidopsis and Rice. Int. J. Mol. Sci. 2013, 14, 5899-5919. https://doi.org/10.3390/ijms14035899
Park HJ, Lee SS, You YN, Yoon DH, Kim B-G, Ahn JC, Cho HS. A Rice Immunophilin Gene, OsFKBP16-3, Confers Tolerance to Environmental Stress in Arabidopsis and Rice. International Journal of Molecular Sciences. 2013; 14(3):5899-5919. https://doi.org/10.3390/ijms14035899
Chicago/Turabian StylePark, Hyun Ji, Sang Sook Lee, Young Nim You, Dae Hwa Yoon, Beom-Gi Kim, Jun Cheul Ahn, and Hye Sun Cho. 2013. "A Rice Immunophilin Gene, OsFKBP16-3, Confers Tolerance to Environmental Stress in Arabidopsis and Rice" International Journal of Molecular Sciences 14, no. 3: 5899-5919. https://doi.org/10.3390/ijms14035899