Differential Protein Expression in Response to Abiotic Stress in Two Potato Species: Solanum commersonii Dun and Solanum tuberosum L.

Abstract
:1. Introduction
2. Results and Discussion
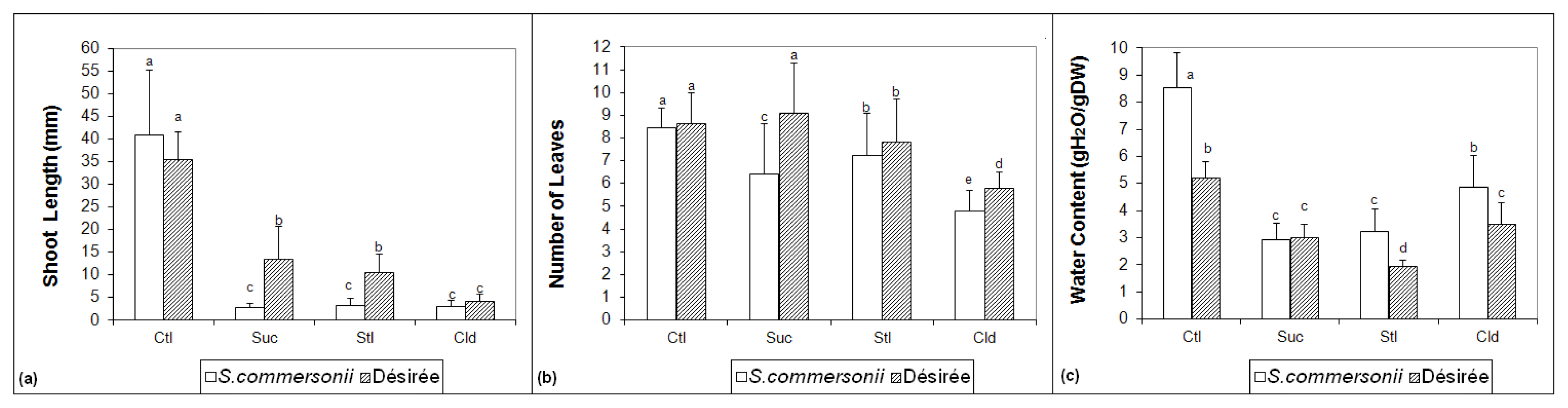
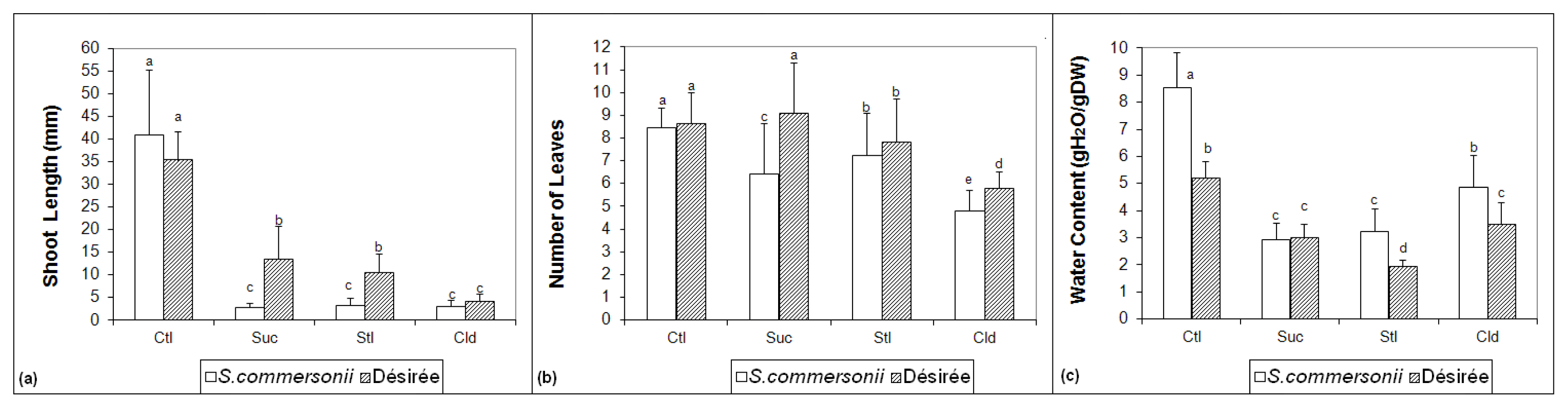
2.1. Morphological Measurements
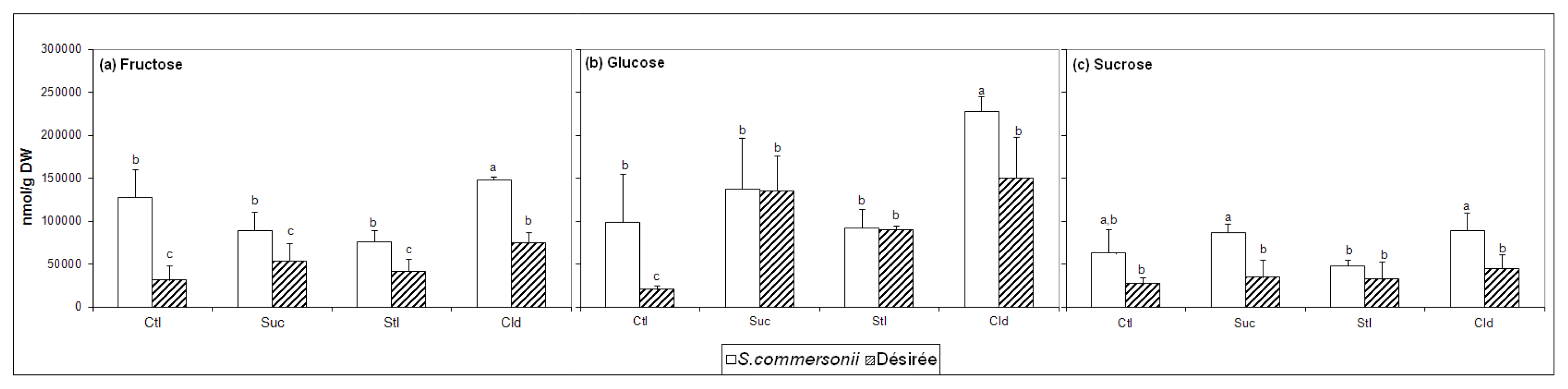
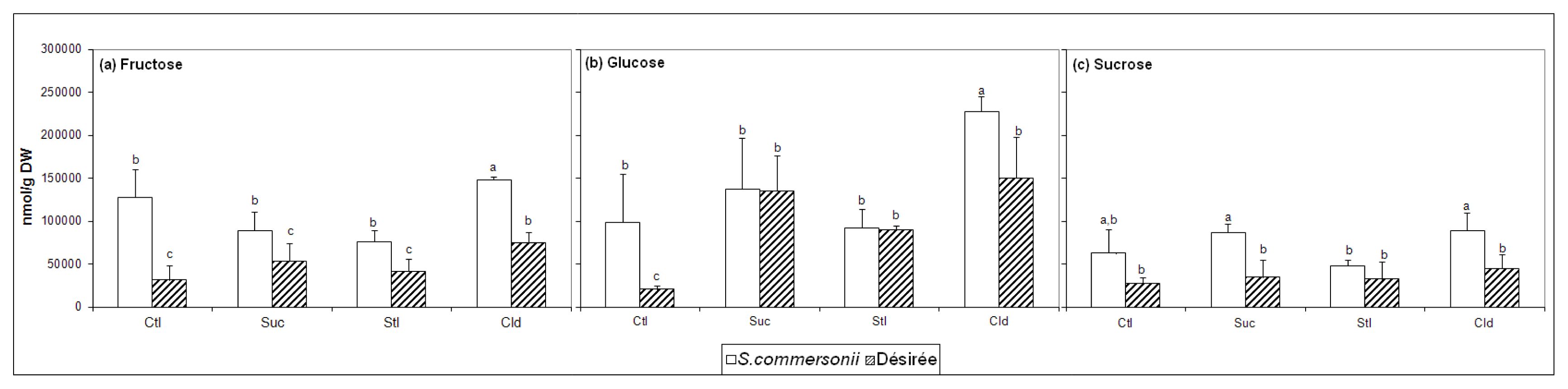
2.2. Soluble Sugars Accumulation
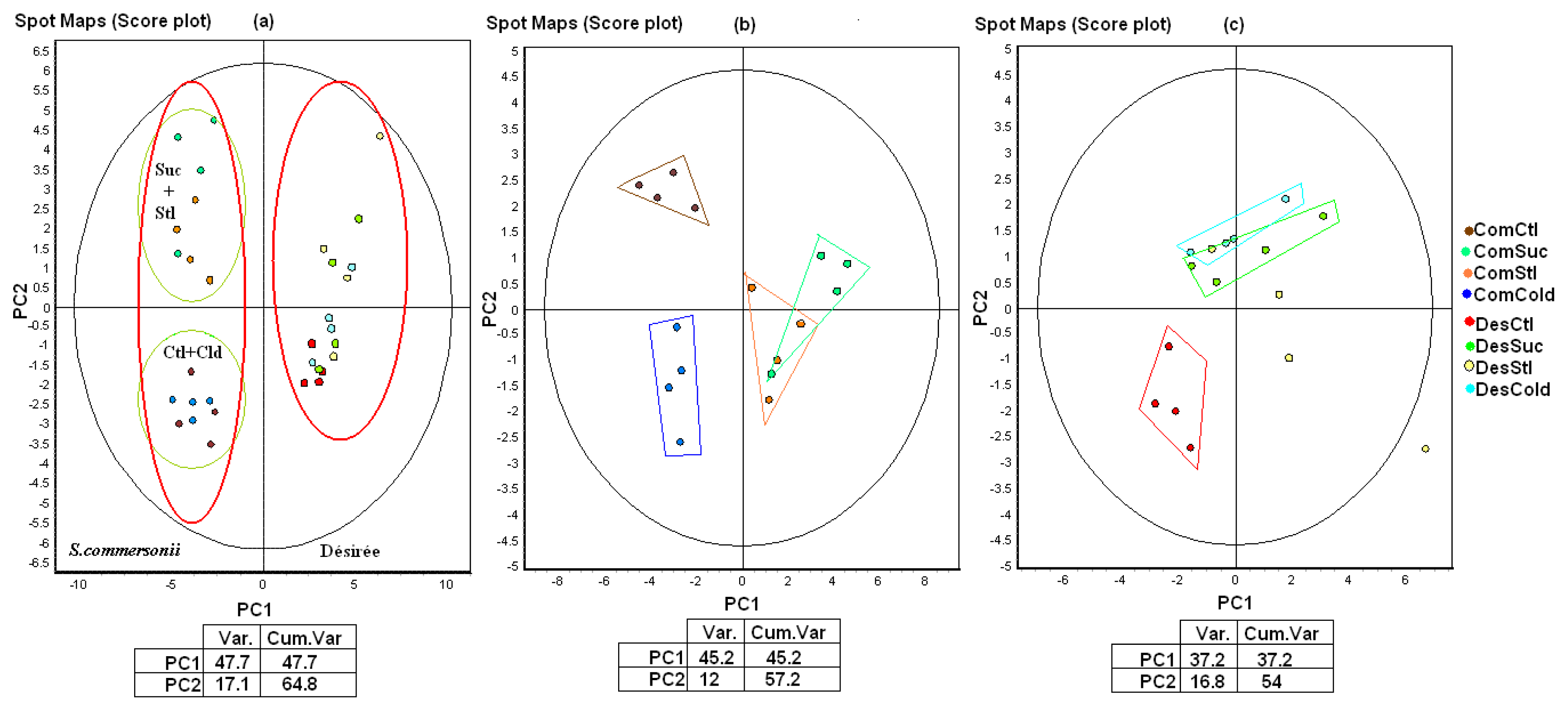
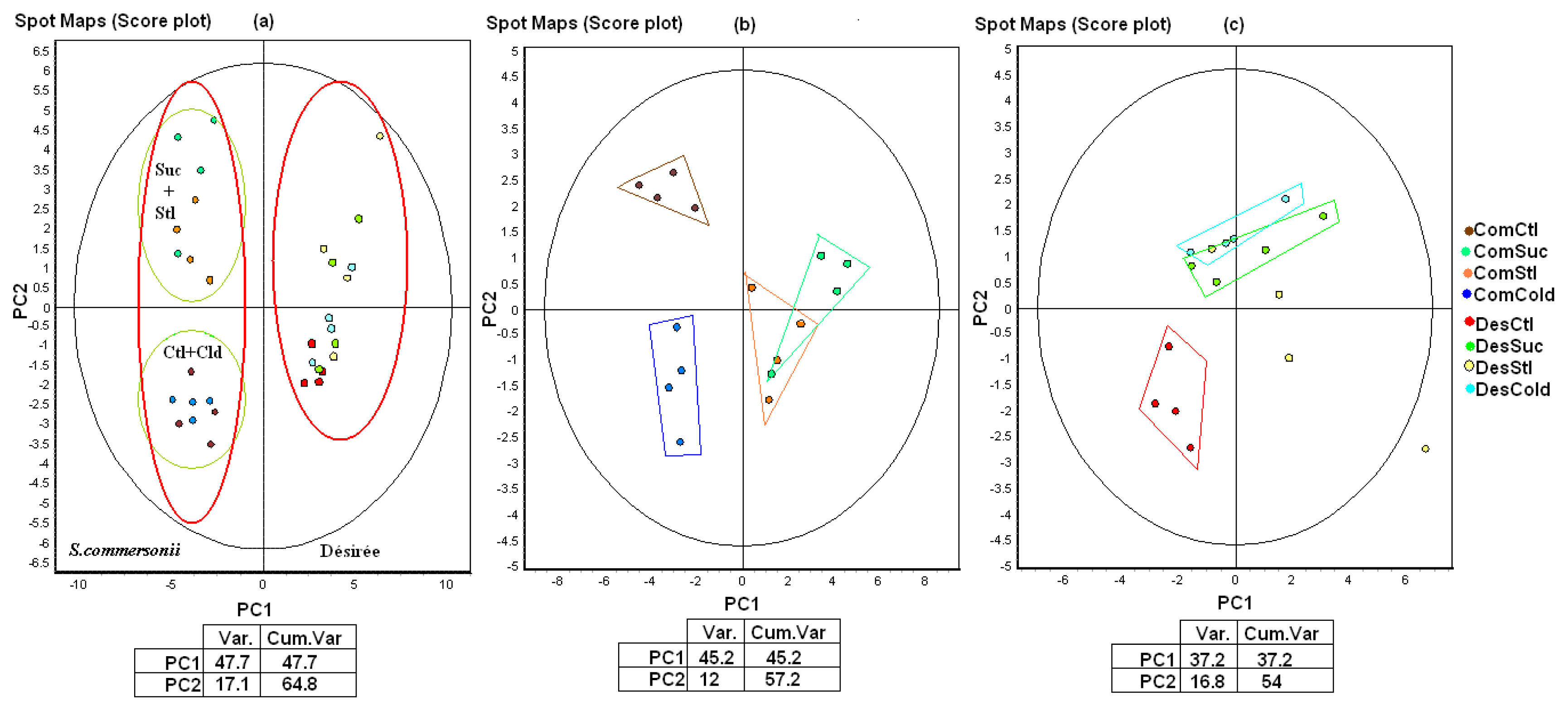
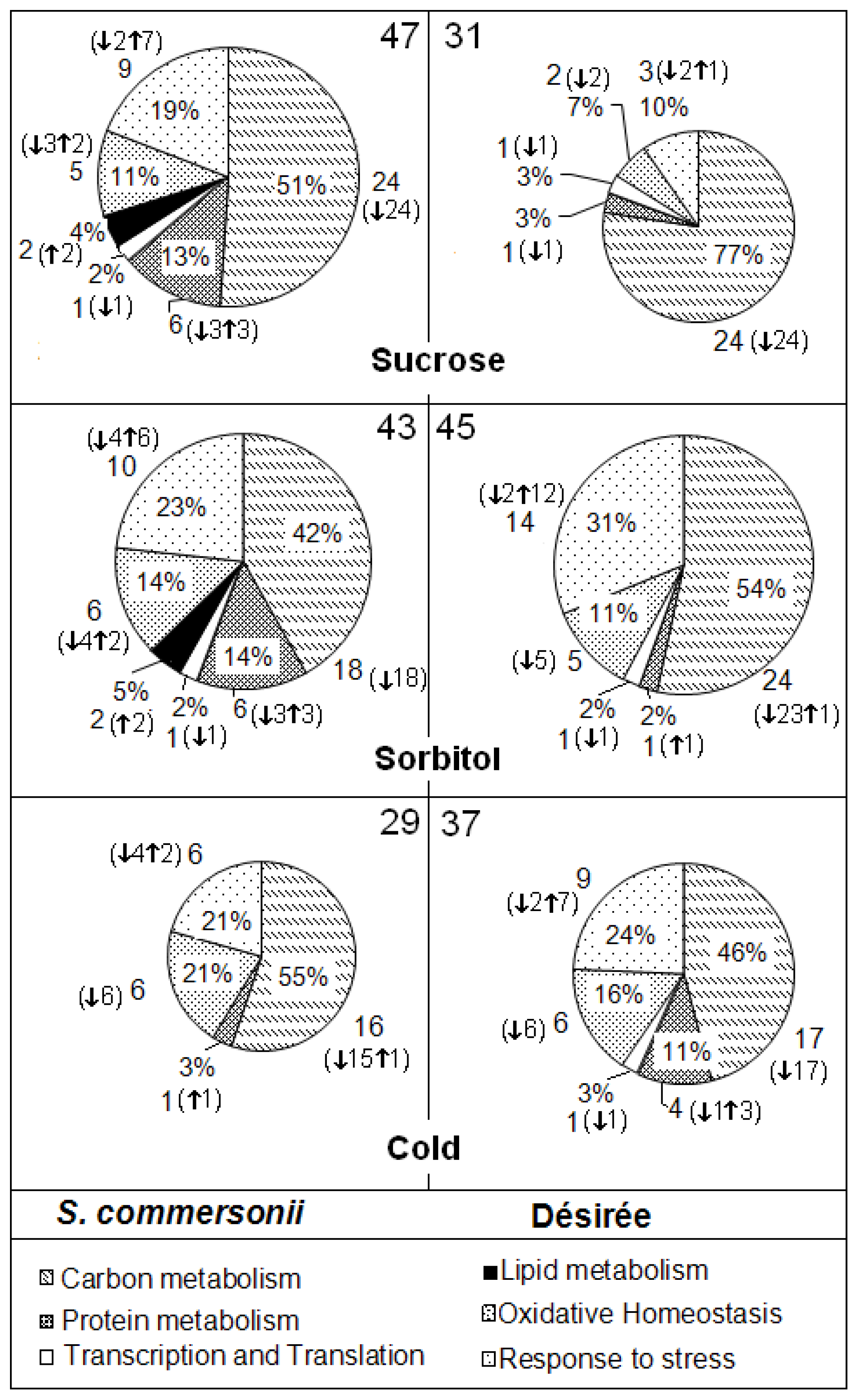
2.3. Proteome Study
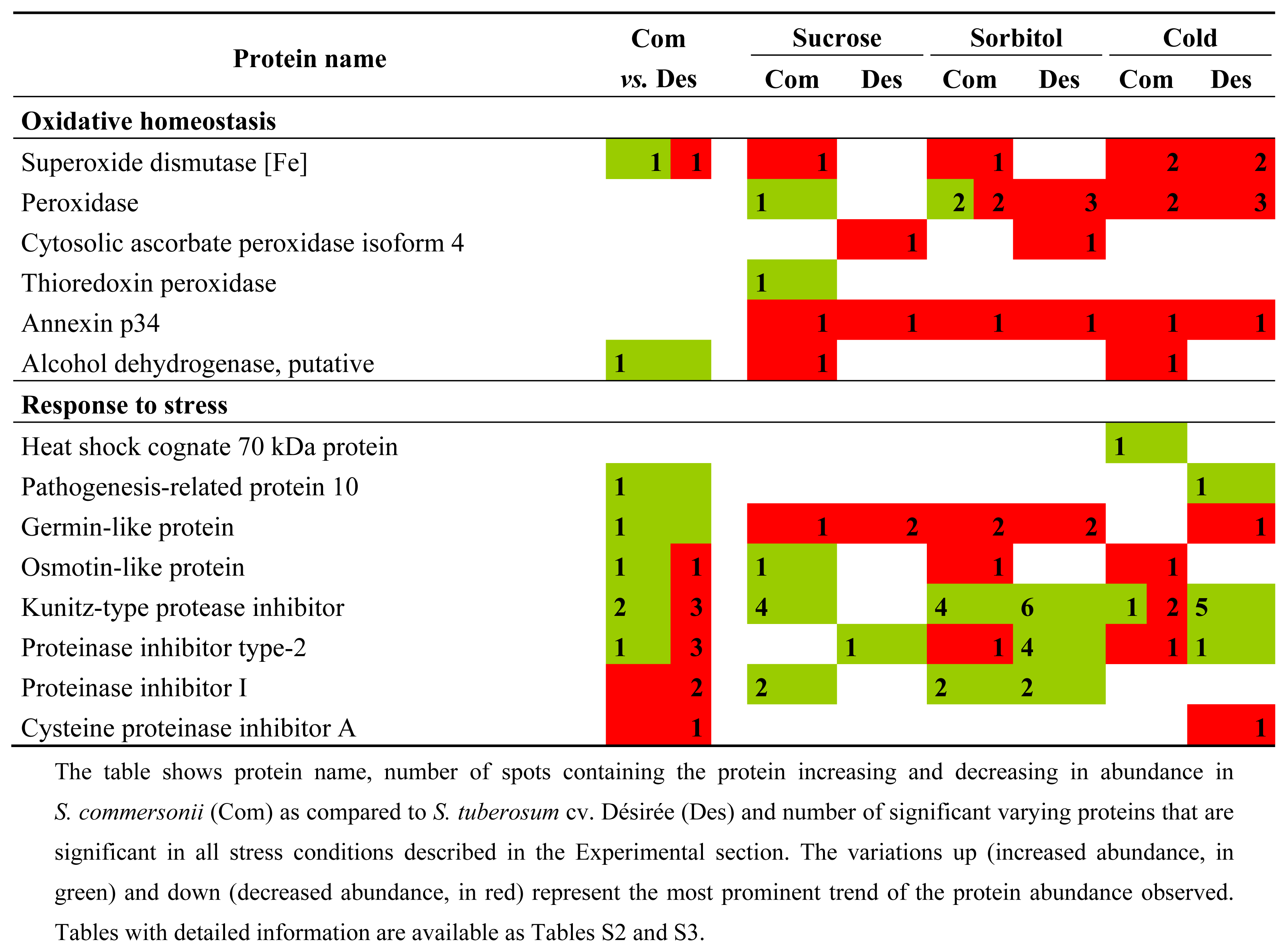
2.3.1. Differences between Species (Control Conditions)
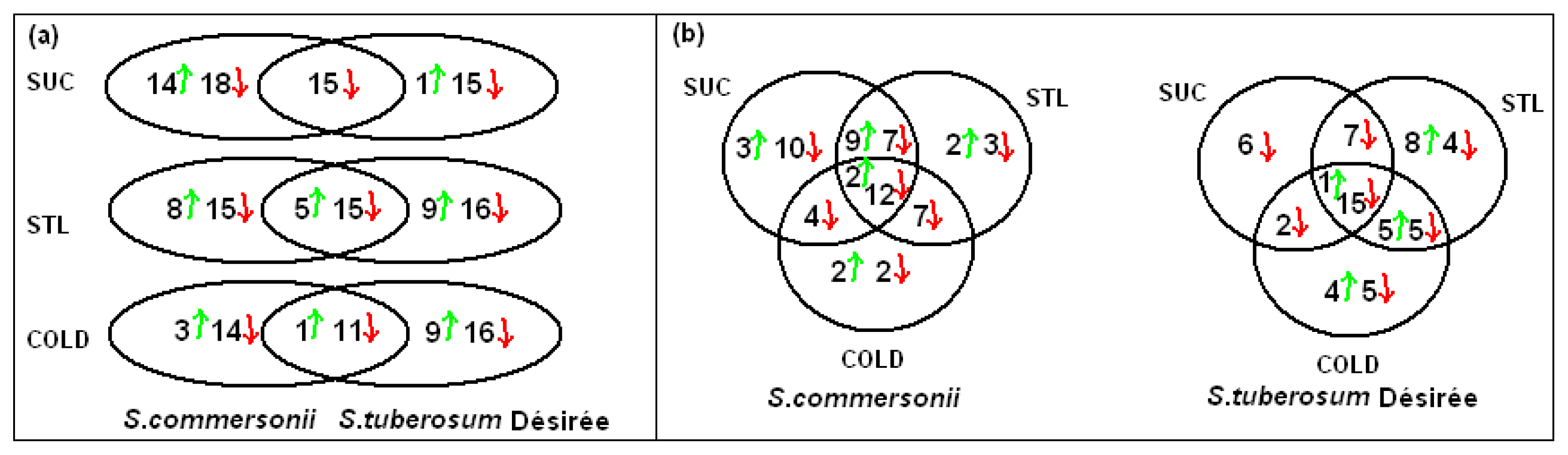
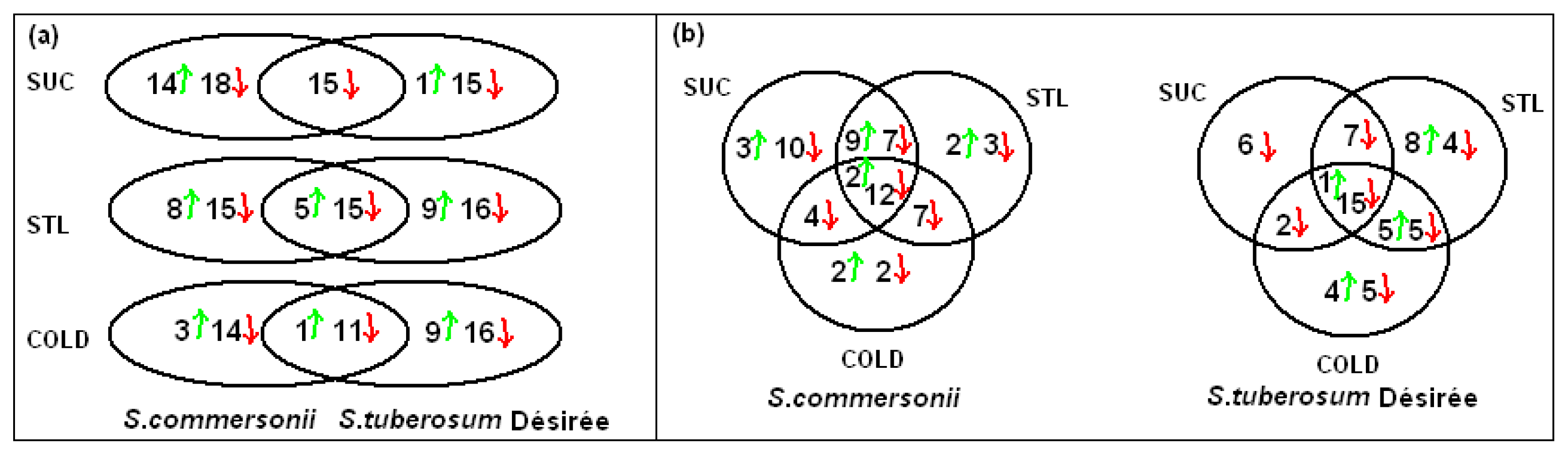
2.3.2. Common Changes to All Treatments in Both Species
2.3.3. Specific Responses during Osmotic Treatments
2.3.4. Changes Produced by Cold Treatment
3. Experimental Section
3.1. Plant Material and Stress Treatments
3.2. Morphological Study
3.3. Soluble Protein Extraction and Labeling
3.4. Electrophoresis
3.5. Image Capture and Analysis
3.6. Protein Identification
3.7. Carbohydrate Extraction and Quantification
4. Conclusions
Acknowledgments
Conflict of Interest
References
- FAOSTAT (Statistics division for the Food & Agriculture Organization). Available online: http://faostat3.fao.org/home/index.html accessed on 12 September 2012.
- FAO (Food & Agriculture Organization). Potato World: Production and Consumption. Available online: http://www.potato.2008.org/en/world/index.html accessed on 12 September 2012.
- Roca, W.; Espinoza, C.; Panta, A. Potential of plant biotechnology for biodiversity research and utilization in Latin America and the Caribbean. AgBioForum J 2003, 7, 13–22. [Google Scholar]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Molecular responses to dehydration and low temperature: Differences and cross-talk between two stress signaling pathways. Curr. Opin. Plant Biol 2000, 3, 217–223. [Google Scholar]
- Buchanan, B.; Gruissemen, W.; Jones, R. Biochemistry and Molecular Biology of Plants; American Society of Plant Physiologists: Rockville, MD, USA, 2000. [Google Scholar]
- Chen, H.H.; Li, P.H. Biochemical changes in tuber-bearing solanum species in relation to frost hardiness during cold acclimation. Plant Physiol 1980, 66, 414–421. [Google Scholar]
- Keller, E.R.J.; Kaczmarczyk, A.; Senula, A. Cryopreservation for plant genebanks—A matter between high expectations and cautious reservation. Cryoletters 2008, 29, 53–62. [Google Scholar]
- Panis, B.; Lambardi, M. Status of cryopreservation technologies in plants (crops and forest trees). In The Role of Biotechnology in Exploring and Protecting Agricultural Genetic Resources; Ruane, J., Sonnino, A., Eds.; FAO (Food & Agriculture Organization): Rome, Italy, 2006. [Google Scholar]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; Gonzalez, J.A.; Hilal, M.; Prado, F.E. Soluble sugars–Metabolism, sensing and abiotic stress. Plant Signal. Behav 2009, 4, 388–393. [Google Scholar]
- Oono, Y.; Seki, M.; Nanjo, T.; Narusaka, M.; Fujita, M.; Satoh, R.; Satou, M.; Sakurai, T.; Ishida, J.; Akiyama, K.; et al. Monitoring expression profiles of Arabidopsis gene expression during rehydration process after dehydration using Ca 7000 full-length cDNA microarray. Plant J 2003, 34, 868–887. [Google Scholar]
- Rensink, W.A.; Buell, C.R. Microarray expression profiling resources for plant genomics. Trends Plant Sci 2005, 10, 603–609. [Google Scholar]
- Legay, S.; Lamoureux, D.; Hausman, J.F.; Hoffmann, L.; Evers, D. Monitoring gene expression of potato under salinity using cDNA microarrays. Plant Cell Rep 2009, 28, 1799–1816. [Google Scholar]
- Evers, D.; Lefevre, I.; Legay, S.; Lamoureux, D.; Hausman, J.F.; Rosales, R.O.; Marca, L.R.; Hoffmann, L.; Bonierbale, M.; Schafleitner, R. Identification of drought-responsive compounds in potato through a combined transcriptomic and targeted metabolite approach. J. Exp. Bot 2010, 61, 2327–2343. [Google Scholar]
- Carpentier, S.C.; Coemans, B.; Podevin, N.; Laukens, K.; Witters, E.; Matsumura, H.; Terauchi, R.; Swennen, R.; Panis, B. Functional genomics in a non-model crop: Transcriptomics or proteomics? Physiol. Plant 2008, 133, 117–130. [Google Scholar]
- Evers, D.; Legay, S.; Lamoureux, D.; Hausman, J.F.; Hoffmann, L.; Renaut, J. Towards a synthetic view of potato cold and salt stress response by transcriptomic and proteomic analyses. Plant Mol. Biol 2012, 78, 503–514. [Google Scholar]
- Canovas, F.M.; Dumas-Gaudot, E.; Recorbet, G.; Jorrin, J.; Mock, H.P.; Rossignol, M. Plant proteome analysis. Proteomics 2004, 4, 285–298. [Google Scholar]
- Vincent, D.; Zivy, M. Plant proteome responses to abiotic stress. In Plant Proteomics; Šamaj, J., Thelen, J.J., Eds.; Springer-Verlag: Berlin, Germany, 2007. [Google Scholar]
- Carpentier, S.C.; Witters, E.; Laukens, K.; van Onckelen, H.; Swennen, R.; Panis, B. Banana (Musa Spp.) as a model to study the meristem proteome: Acclimation to osmotic stress. Proteomics 2007, 7, 92–105. [Google Scholar]
- Renaut, J.; Bohler, S.; Hausman, J.F.; Hoffmann, L.; Sergeant, K.; Ahsan, N.; Jolivet, Y.; Dizengremel, P. The impact of atmospheric composition on plants: A case study of ozone and poplar. Mass Spectrom. Rev 2009, 28, 495–516. [Google Scholar]
- Bauw, G.; Nielsen, H.V.; Emmersen, J.; Nielsen, K.L.; Jorgensen, M.; Welinder, K.G. Patatins, Kunitz protease inhibitors and other major proteins in tuber of potato cv. Kuras. FEBS J 2006, 273, 3569–3584. [Google Scholar]
- Lehesranta, S.J.; Davies, H.V.; Shepherd, L.V.; Nunan, N.; McNicol, J.W.; Auriola, S.; Koistinen, K.M.; Suomalainen, S.; Kokko, H.I.; Karenlampi, S.O. Comparison of tuber proteomes of potato varieties, landraces, and genetically modified lines. Plant Physiol 2005, 138, 1690–1699. [Google Scholar]
- Delaplace, P.; Fauconnier, M.L.; Sergeant, K.; Dierick, J.F.; Oufir, M.; van der Wal, F.; America, A.H.; Renaut, J.; Hausman, J.F.; du Jardin, P. Potato (Solanum tuberosum L.) tuber ageing induces changes in the proteome and antioxidants associated with the sprouting pattern. J. Exp. Bot 2009, 60, 1273–1288. [Google Scholar]
- Coulthurst, S.J.; Lilley, K.S.; Salmond, G.P. Genetic and proteomic analysis of the role of luxs in the enteric phytopathogen, Erwinia carotovora. Mol. Plant Pathol 2006, 7, 31–45. [Google Scholar]
- An Nguyen, T.T.; Michaud, D.; Cloutier, C. Proteomic profiling of aphid Macrosiphum euphorbiae responses to host-plant-mediated stress induced by defoliation and water deficit. J. Insect Physiol 2007, 53, 601–611. [Google Scholar]
- Mattinen, L.; Nissinen, R.; Riipi, T.; Kalkkinen, N.; Pirhonen, M. Host-extract induced changes in the secretome of the plant pathogenic bacterium Pectobacterium atrosepticum. Proteomics 2007, 7, 3527–3537. [Google Scholar]
- Renaut, J.; Planchon, S.; Oufir, M.; Hausman, J.F.; Hoffmann, L.; Evers, D. Identification of proteins from potato leaves submitted to chilling temperature. In Plant Cold Hardiness: From the Laboratory to the Field; Gusta, L.V., Wisniewski, M.E., Tanino, K.K., Eds.; CAB International: Wallingford, UK, 2009. [Google Scholar]
- Espinoza, N.; Estrada, R.; Tovar, P.; Bryan, J.; Dodds, J.H. Tissue Culture Micropropagation, Conservation, and Export of Potato Germplasm; Specialized Technology Document 1; Lima, International Potato Center: Lima, Peru, 1986; pp. 1–20. [Google Scholar]
- Zhang, Y.; Donnelly, D.J. In vitro ranking for salinity tolerance of potato cultivars. Potato Res 1997, 40, 285–295. [Google Scholar]
- Gopal, J.; Iwama, K. In vitro screening of potato against water-stress mediated through Sorbitol and Polyethylene Glycol. Plant Cell Rep 2007, 26, 693–700. [Google Scholar]
- Gopal, J.; Iwama, K.; Jitsuyama, Y. Effect of water stress mediated through agar on in vitro growth of potato. In Vitro Cell. Dev. Biol. Plant 2008, 44, 221–228. [Google Scholar]
- Ruana, Y.-L.; Jinb, Y.; Yang, Y.-J.; Lid, G.-J.; Boyere, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol 2004, 7, 235–246. [Google Scholar]
- GOanna Tool, From AgBase Online Software. Available online: http://agbase.msstate.edu/cgi-bin/generateBlastPage.pl accessed on 2 August 2011.
- Li, P.H.; Palta, J.P. Frost hardening and freezing stress in tuber-bearing Solanum species. In Plant Cold Hardiness and Freezing Stress; Li, P.H., Sakai, A., Eds.; Academic Press: New York, NY, USA, 1978. [Google Scholar]
- Seppänen, M.M.; Majaharju, M.; Somersalo, S.; Pehu, E. Freezing tolerance, cold acclimation and oxidative stress in potato. Paraquat tolerance is related to acclimation but is a poor indicator of freezing tolerance. Physiol. Plantarum 1998, 102, 454–460. [Google Scholar]
- Chen, H.H.; Li, P.H. Characteristics of cold acclimation and deacclimation in tuber-bearing Solanum species. Plant Physiol 1980, 65, 1146–1148. [Google Scholar]
- Seppänen, M.M.; Nissinen, O.; Perälä, S. Freezing tolerance and low temperature photoinhibition tolerance in cultivated potato hybrids. Agric. Food Sci. Finl 2001, 10, 153–163. [Google Scholar]
- Spooner, D.; Castillo, R. Reexamination of series relationships of south american wild potatoes (Solanaceae: Solanum Sect. Petota): Evidence from chloroplast DNA restriction site variation. Am. J. Bot 1997, 84, 671. [Google Scholar]
- Hurter, N.P.A.; Palta, J.P.; Li, P.H.; Carter, J.V. Comparison of the structure and function of ribulose bisphosphate carboxylase-oxygenase from a frost-hardy and non-hardy potato species. Can. J. Biochem 1981, 59, 280–289. [Google Scholar]
- Shvaleva, A.L.; Costa, E.; Silva, F.; Breia, E.; Jouve, J.; Hausman, J.F.; Almeida, M.H.; Maroco, J.P.; Rodrigues, M.L.; Pereira, J.S.; Chaves, M.M. Metabolic responses to water deficit in two Eucalyptus globulus clones with contrasting drought sensitivity. Tree Physiol 2006, 26, 239–248. [Google Scholar]
- Chaves, M.M. Effects of water deficits on carbon assimilation. J. Exp. Bot 1991, 42, 1–16. [Google Scholar]
- Renaut, J.; Lutts, S.; Hoffmann, L.; Hausman, J.F. Responses of poplar to chilling temperatures: Proteomic and physiological aspects. Plant Biol. (Stuttg) 2004, 6, 81–90. [Google Scholar]
- Oufir, M.; Legay, S.; Nicot, N.; van Moer, K.; Hoffmann, L.; Renaut, J.; Hausman, J.-F.; Evers, D. Gene expression in potato during cold exposure: Changes in carbohydrate and polyamine metabolisms. Plant Sci 2008, 175, 839–852. [Google Scholar]
- Calvert, C.M.; Gant, S.J.; Bowles, D.J. Tomato Annexins P34 and P35 bind to F-Actin and display nucleotide phosphodiesterase activity inhibited by phospholipid binding. Plant Cell 1996, 8, 333–342. [Google Scholar]
- Caliskan, M. Germin, an oxalate oxidase, has a function in many aspects of plant life. Turk. J. Bot 2000, 24, 717–724. [Google Scholar]
- Lane, B.G. Oxalate, Germins, and higher-plant pathogens. IUBMB Life 2002, 53, 67–75. [Google Scholar]
- Berna, A.; Bernier, F. Regulation by biotic and abiotic stress of a wheat germin gene encoding oxalate oxidase, a H2O2-producing enzyme. Plant Mol. Biol 1999, 39, 539–549. [Google Scholar]
- Donaldson, P.A.; Anderson, T.; Lane, B.G.; Davidson, A.L.; Simmonds, D.H. Soybean plants expressing an active oligomeric oxalate oxidase from the wheat Gf-2.8 (Germin) gene are resistant to the oxalate-secreting pathogen Sclerotina sclerotiorum. Physiol. Mol. Plant Pathol 2001, 59, 297–307. [Google Scholar]
- Ono, M.; Sage-Ono, K.; Inoue, M.; Kamada, H.; Harada, H. Transient increase in the level of mRNA for a Germin-like protein in leaves of the short-day plant Pharbitis nil during the photoperiodic induction of flowering. Plant Cell Physiol 1996, 37, 855–861. [Google Scholar]
- Berna, A.; Bernier, F. Regulated expression of a wheat Germin gene in tobacco: Oxalate oxidase activity and apoplastic localization of the heterologous protein. Plant Mol. Biol 1997, 33, 417–429. [Google Scholar]
- Davidson, R.; Manosalva, P.; Snelling, J.; Bruce, M.; Leung, H.; Leach, J. Rice Germin-like proteins: Allelic diversity and relationships to early stress responses. Rice 2010, 3, 43–55. [Google Scholar]
- Bray, E.A. Genes commonly regulated by water-deficit stress in Arabidopsis thaliana. J. Exp. Bot 2004, 55, 2331–2341. [Google Scholar]
- Pinheiro, C.; Kehr, J.; Ricardo, C.P. Effect of water stress on lupin stem protein analysed by two-dimensional gel electrophoresis. Planta 2005, 221, 716–728. [Google Scholar]
- Heibges, A.; Salamini, F.; Gebhardt, C. Functional comparison of homologous members of three groups of Kunitz-type enzyme inhibitors from potato tubers (Solanum tuberosum L.). Mol. Genet. Genomics 2003, 269, 535–541. [Google Scholar]
- Molosov, V.V.; Valueva, T.A. Role of proteinase inhibitors and their function in plants. A review. Appl. Biochem. Microbiol 2005, 41, 227–247. [Google Scholar]
- Hendriks, T.; Vreugdenhil, D.; Stiekema, W.J. Patatin and four serine proteinase inhibitor genes are differentially expressed during potato tuber development. Plant Mol. Biol 1991, 17, 385–394. [Google Scholar]
- Sin, S.F.; Yeung, E.C.; Chye, M.L. Downregulation of Solanum americanum genes encoding proteinase inhibitor II causes defective seed development. Plant J 2006, 45, 58–70. [Google Scholar]
- Lopez, F.; Vansuyt, G.; Derancourt, J.; Fourcroy, P.; Casse-Delbart, F. Identification by 2D-PAGE analysis of salt-stress induced proteins in radish (Raphanus sativus). Cell Mol. Biol. (Noisy-le-grand) 1994, 40, 85–90. [Google Scholar]
- Gosti, F.; Bertauche, N.; Vartanian, N.; Giraudat, J. Abscisic acid-dependent and -independent regulation of gene expression by progressive drought in Arabidopsis thaliana. Mol. Gen. Genet 1995, 246, 10–18. [Google Scholar]
- Legay, S.; Lefevre, I.; Lamoureux, D.; Barreda, C.; Luz, R.T.; Gutierrez, R.; Quiroz, R.; Hoffmann, L.; Hausman, J.F.; Bonierbale, M.; et al. Carbohydrate metabolism and cell protection mechanisms differentiate drought tolerance and sensitivity in advanced potato clones (Solanum tuberosum L.). Funct. Integr. Genomics 2011, 11, 275–291. [Google Scholar]
- Rabbani, M.A.; Maruyama, K.; Abe, H.; Khan, M.A.; Katsura, K.; Ito, Y.; Yoshiwara, K.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Monitoring expression profiles of rice genes under cold, drought, and high-salinity stresses and abscisic acid application using cDNA Microarray and RNA Gel-Blot analyses. Plant Physiol 2003, 133, 1755–1767. [Google Scholar]
- Schafleitner, R.; Gutierrez Rosales, R.O.; Gaudin, A.; varado Aliaga, C.A.; Martinez, G.N.; Tincopa Marca, L.R.; Bolivar, L.A.; Delgado, F.M.; Simon, R.; Bonierbale, M. Capturing candidate drought tolerance traits in two native andean potato clones by transcription profiling of field grown plants under water stress. Plant Physiol. Biochem 2007, 45, 673–690. [Google Scholar]
- Bohler, S.; Sergeant, K.; Lefevre, I.; Jolivet, Y.; Hoffmann, L.; Renaut, J.; Dizengremel, P.; Hausman, J.F. Differential impact of chronic ozone exposure on expanding and fully expanded poplar leaves. Tree Physiol 2010, 30, 1415–1432. [Google Scholar]
- Merewitz, E.B.; Gianfagna, T.; Huang, B. Protein accumulation in leaves and roots associated with improved drought tolerance in creeping bentgrass expressing an Ipt gene for cytokinin synthesis. J. Exp. Bot 2011, 62, 5311–5333. [Google Scholar]
- Ito, H.; Iwabuchi, M.; Ogawa, K. The sugar-metabolic enzymes aldolase and triose-phosphate isomerase are targets of glutathionylation in Arabidopsis thaliana: Detection using biotinylated glutathione. Plant Cell Physiol 2003, 44, 655–660. [Google Scholar]
- Rizhsky, L.; Liang, H.; Shuman, J.; Shulaev, V.; Davletova, S.; Mittler, R. When defense pathways collide. The response of arabidopsis to a combination of drought and heat stress. Plant Physiol 2004, 134, 1683–1696. [Google Scholar]
- Lu, Z.; Liu, D.; Liu, S. Two rice cytosolic ascorbate peroxidases differentially improve salt tolerance in transgenic Arabidopsis. Plant Cell Rep 2007, 26, 1909–1917. [Google Scholar]
- Miller, G.; Suzuki, N.; Rizhsky, L.; Hegie, A.; Koussevitzky, S.; Mittler, R. Double mutants deficient in cytosolic and thylakoid ascorbate peroxidase reveal a complex mode of interaction between reactive oxygen species, plant development, and response to abiotic stresses. Plant Physiol 2007, 144, 1777–1785. [Google Scholar]
- Koussevitzky, S.; Suzuki, N.; Huntington, S.; Armijo, L.; Sha, W.; Cortes, D.; Shulaev, V.; Mittler, R. Ascorbate peroxidase 1 plays a key role in the response of Arabidopsis thaliana to stress combination. J. Biol. Chem 2008, 283, 34197–34203. [Google Scholar]
- Maruta, T.; Tanouchi, A.; Tamoi, M.; Yabuta, Y.; Yoshimura, K.; Ishikawa, T.; Shigeoka, S. Arabidopsis chloroplastic ascorbate peroxidase isoenzymes play a dual role in photoprotection and gene regulation under photooxidative stress. Plant Cell Physiol 2010, 51, 190–200. [Google Scholar]
- Pnueli, L.; Liang, H.; Rozenberg, M.; Mittler, R. Growth suppression, altered stomatal responses, and augmented induction of Heat Shock Proteins in cytosolic ascorbate peroxidase (Apx1)-deficient Arabidopsis plants. Plant J 2003, 34, 187–203. [Google Scholar]
- Asai, N.; Matsuyama, T.; Tamaoki, M.; Nakajima, N.; Kubo, A.; Aono, M.; Kato, T.; Tabata, S.; Shirano, Y.; Shibata, D.; et al. Compensation for lack of a cytosolic ascorbate peroxidase in an Arabidopsis mutant by activation of multiple antioxidative systems. Plant Sci 2004, 166, 1547–1554. [Google Scholar]
- Bonifacio, A.; Martins, M.O.; Ribeiro, C.W.; Fontenele, A.V.; Carvalho, F.E.; Margis-Pinheiro, M.; Silveira, J.A. Role of peroxidases in the compensation of cytosolic ascorbate peroxidase knockdown in rice plants under abiotic stress. Plant Cell Environ 2011, 34, 1705–1722. [Google Scholar]
- Ribeiro, C.W.; Carvalho, F.E.; Rosa, S.B.; Alves-Ferreira, M.; Andrade, C.M.; Ribeiro-Alves, M.; Silveira, J.A.; Margis, R.; Margis-Pinheiro, M. Modulation of genes related to specific metabolic pathways in response to cytosolic ascorbate peroxidase knockdown in rice plants. Plant Biol 2012, 14, 944–955. [Google Scholar]
- Guo, G.; Ge, P.; Ma, C.; Li, X.; Lv, D.; Wang, S.; Ma, W.; Yan, Y. Comparative proteomic analysis of salt response proteins in seedling roots of two wheat varieties. J. Proteomics 2012, 75, 1867–1885. [Google Scholar]
- Abdalla, K.O.; Rafudeen, M.S. Analysis of the nuclear proteome of the resurrection plant Xerophyta viscosa in response to dehydration stress using iTRAQ with 2DLC and tandem mass spectrometry. J. Proteomics 2012, 75, 2361–2374. [Google Scholar]
- Boscariol-Camargo, R.L.; Berger, I.J.; Souza, A.A.; do Amaral, A.M.; Carlos, E.F.; Freitas-Astúa, J.; Takita, M.A.; Targon, M.L.P.N.; Medina, C.L.; Reis, M.S.; et al. In silico analysis of ESTs from roots of rangpur lime (Citrus limonia Osbeck) under water stress. Genetics Mol. Biol 2007, 30, 906–916. [Google Scholar]
- Kieffer, P.; Dommes, J.; Hoffmann, L.; Hausman, J.F.; Renaut, J. Quantitative changes in protein expression of cadmium-exposed poplar plants. Proteomics 2008, 8, 2514–2530. [Google Scholar]
- Baneyx, F.; Bertsch, U.; Kalbach, C.E.; van der Vies, S.M.; Soll, J.; Gatenby, A.A. Spinach chloroplast Cpn21 Co-chaperonin possesses two functional domains fused together in a toroidal structure and exhibits nucleotide-dependent binding to plastid chaperonin 60. J. Biol. Chem 1995, 270, 10695–10702. [Google Scholar]
- Gutteridge, S.; Gatenby, A.A. Rubisco synthesis, assembly, mechanism, and regulation. Plant Cell 1995, 7, 809–819. [Google Scholar]
- Yamaguchi, K.; Subramanian, A.R. Proteomic identification of all plastid-specific ribosomal proteins in higher plant chloroplast 30S ribosomal subunit. Eur. J. Biochem 2003, 270, 190–205. [Google Scholar]
- Feller, U. Proteolysis. In Plant Cell Death Processes; Noodén, L.D., Ed.; Academic Press: San Diego, CA, USA, 2004. [Google Scholar]
- Veierskov, B.; Ferguson, I.B. Conjugation of Ubiquitin to proteins from green plant tissues. Plant Physiol 1991, 96, 4–9. [Google Scholar]
- Waters, M.T.; Langdale, J.A. The making of a chloroplast. EMBO J 2009, 28, 2861–2873. [Google Scholar]
- Holk, A.; Rietz, S.; Zahn, M.; Quader, H.; Scherer, G.F. Molecular identification of cytosolic, Patatin-related phospholipases A from Arabidopsis with potential functions in plant signal transduction. Plant Physiol 2002, 130, 90–101. [Google Scholar]
- Banerji, S.; Flieger, A. Patatin-like proteins: A new family of lipolytic enzymes present in bacteria? Microbiology 2004, 150, 522–525. [Google Scholar]
- Bamberg, J.B.; Palta, J.P.; Vega, S.E. Solanum commersonii cytoplasm does not improve freezing tolerance in substitution backcross hybrids with frost sensitive potato species. Am. J. Potato Res 2005, 82, 251–254. [Google Scholar]
- Beere, H.M.; Wolf, B.B.; Cain, K.; Mosser, D.D.; Mahboubi, A.; Kuwana, T.; Tailor, P.; Morimoto, R.I.; Cohen, G.M.; Green, D.R. Heat-Shock Protein 70 inhibits apoptosis by preventing recruitment of Procaspase-9 to the Apaf-1 apoptosome. Nat. Cell Biol 2000, 2, 469–475. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissues cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar]
- Skynner, H.A.; Rosahl, T.W.; Knowles, M.R.; Salim, K.; Reid, L.; Cothliff, R.; McAllister, G.; Guest, P.C. Alterations of stress related proteins in genetically altered mice revealed by two-dimensional differential in-Gel Electrophoresis analysis. Proteomics 2002, 2, 1018–1025. [Google Scholar]
- Bohler, S.; Bagard, M.; Oufir, M.; Planchon, S.; Hoffmann, L.; Jolivet, Y.; Hausman, J.F.; Dizengremel, P.; Renaut, J. A DIGE analysis of developing poplar leaves subjected to ozone reveals major changes in carbon metabolism. Proteomics 2007, 7, 1584–1599. [Google Scholar]
- Sergeant, K.; Spiess, N.; Renaut, J.; Wilhelm, E.; Hausman, J.F. One dry summer: A leaf proteome study on the response of oak to drought exposure. J. Proteomics 2011, 74, 1385–1395. [Google Scholar]
- Guignard, C.; Jouve, L.; Bogeat-Triboulot, M.B.; Dreyer, E.; Hausman, J.F.; Hoffmann, L. Analysis of carbohydrates in plants by High-performance anion-exchange chromatography coupled with electrospray mass spectrometry. J. Chromatogr. A 2005, 1085, 137–142. [Google Scholar]
- Hummel, I.; Pantin, F.; Sulpice, R.; Piques, M.; Rolland, G.; Dauzat, M.; Christophe, A.; Pervent, M.; Bouteille, M.; Stitt, M.; et al. Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: An integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol 2010, 154, 357–372. [Google Scholar]
- Osuna, D.; Usadel, B.; Morcuende, R.; Gibon, Y.; Blasing, O.E.; Hohne, M.; Gunter, M.; Kamlage, B.; Trethewey, R.; Scheible, W.R.; et al. Temporal responses of transcripts, enzyme activities and metabolites after adding sucrose to carbon-deprived arabidopsis seedlings. Plant J 2007, 49, 463–491. [Google Scholar]
- Good, A.G.; Zaplachinski, S.T. The effects of drought stress on free amino acid accumulation and protein synthesis in Brassica napus. Physiol. Plantarum 1994, 90, 9–14. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Folgado, R.; Panis, B.; Sergeant, K.; Renaut, J.; Swennen, R.; Hausman, J.-F. Differential Protein Expression in Response to Abiotic Stress in Two Potato Species: Solanum commersonii Dun and Solanum tuberosum L. Int. J. Mol. Sci. 2013, 14, 4912-4933. https://doi.org/10.3390/ijms14034912
Folgado R, Panis B, Sergeant K, Renaut J, Swennen R, Hausman J-F. Differential Protein Expression in Response to Abiotic Stress in Two Potato Species: Solanum commersonii Dun and Solanum tuberosum L. International Journal of Molecular Sciences. 2013; 14(3):4912-4933. https://doi.org/10.3390/ijms14034912
Chicago/Turabian StyleFolgado, Raquel, Bart Panis, Kjell Sergeant, Jenny Renaut, Rony Swennen, and Jean-Francois Hausman. 2013. "Differential Protein Expression in Response to Abiotic Stress in Two Potato Species: Solanum commersonii Dun and Solanum tuberosum L." International Journal of Molecular Sciences 14, no. 3: 4912-4933. https://doi.org/10.3390/ijms14034912