Impact of Cold Ischemia on Mitochondrial Function in Porcine Hearts and Blood Vessels



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
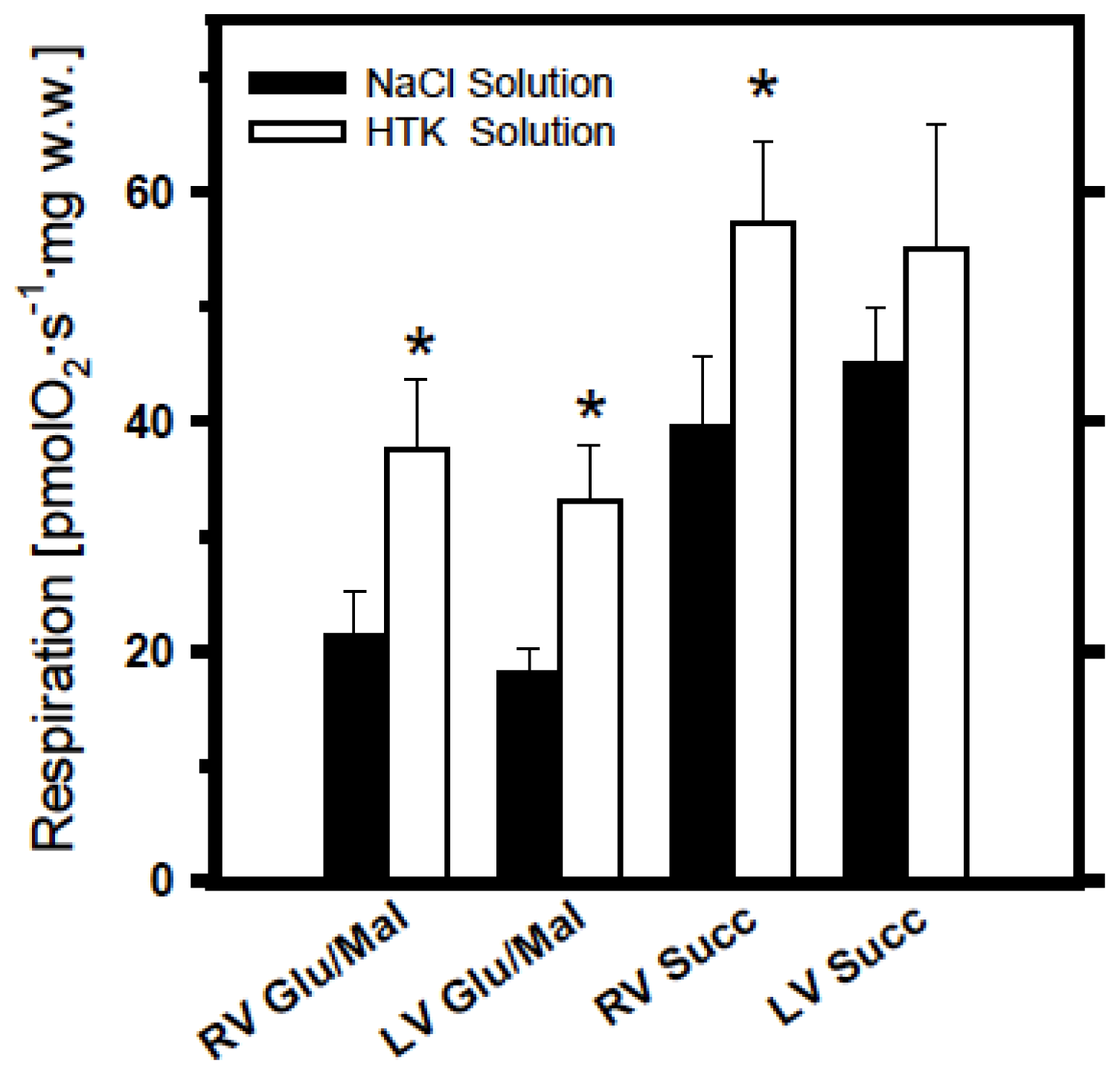
2.1. Analysis of Cold Ischemia Effects on Pig Myocardium
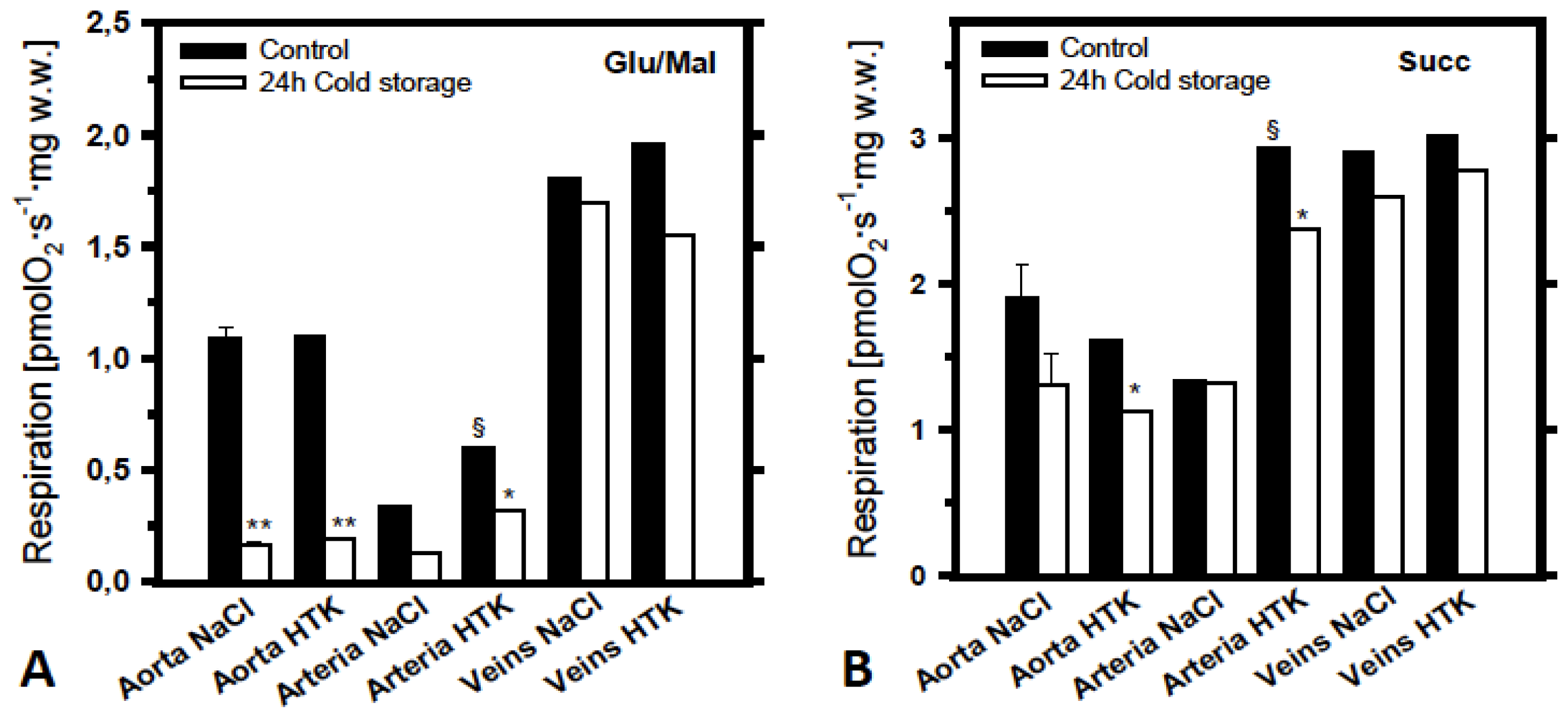
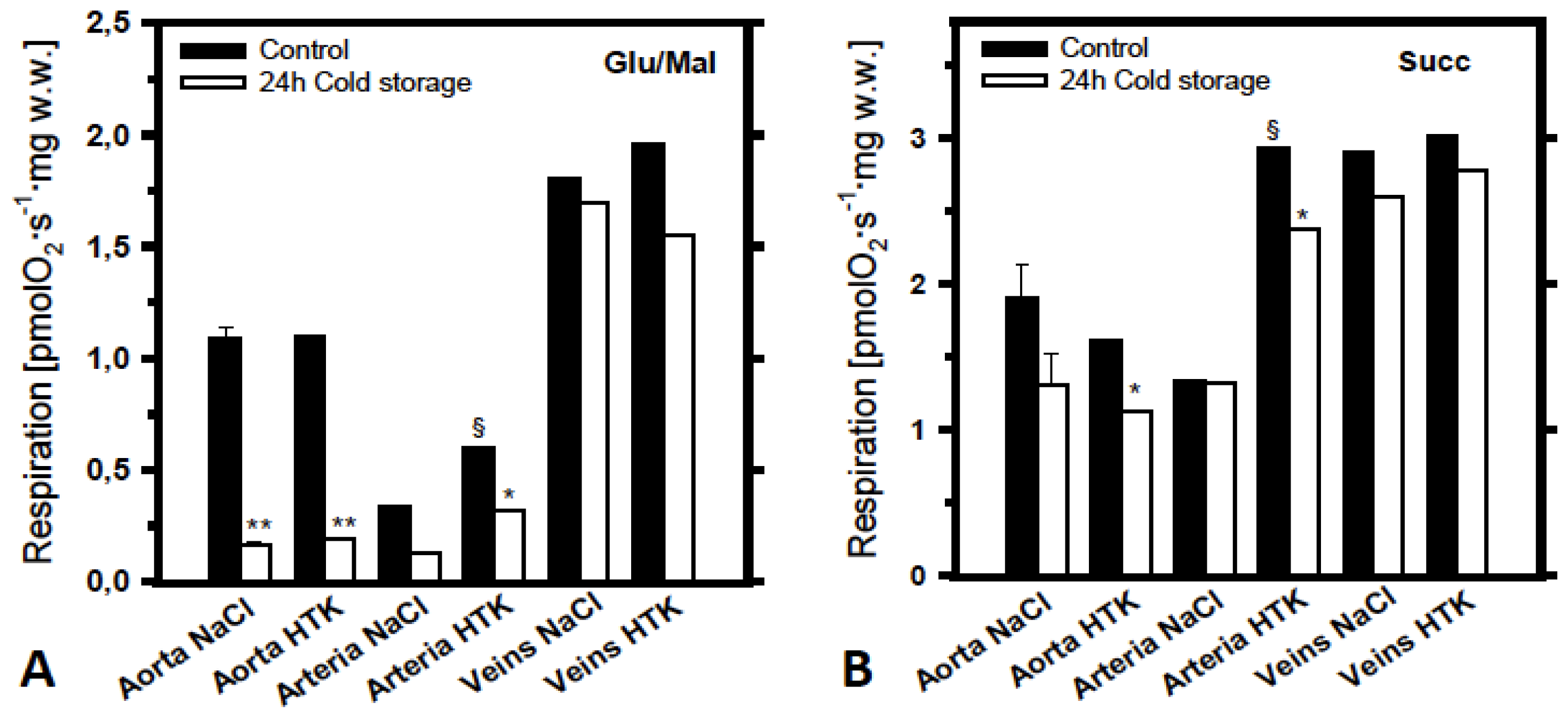
2.2. Analysis of the Effects of Cold Ischemia on Mitochondrial Function in Various Pig Blood Vessels
2.3. Discussion
Limitations
3. Experimental Section
3.1. Animals
3.2. Analysis of Mitochondrial Function
3.3. Storage Solutions and Chemicals
4. Conclusions

Acknowledgments
Conflicts of Interest
References
- Kuznetsov, A.V.; Veksler, V.; Gellerich, F.N.; Saks, V.; Margreiter, R.; Kunz, W.S. Analysis of mitochondrial function in situ in permeabilized muscle fibers, tissues and cells. Nat. Protoc 2008, 3, 965–976. [Google Scholar]
- Andreas, M.; Schmid, A.I.; Keilani, M.; Doberer, D.; Bartko, J.; Crevenna, R.; Moser, E.; Wolzt, M. Effect of ischemic preconditioning in skeletal muscle measured by functional magnetic resonance imaging and spectroscopy: A randomized crossover trial. J. Cardiovasc. Magn. Reson 2011, 13, 32:1–32:10. [Google Scholar]
- Meyerspeer, M.; Kemp, G.J.; Mlynarik, V.; Krssak, M.; Szendroedi, J.; Nowotny, P.; Roden, M.; Moser, E. Direct noninvasive quantification of lactate and high energy phosphates simultaneously in exercising human skeletal muscle by localized magnetic resonance spectroscopy. Magn. Reson. Med 2007, 57, 654–660. [Google Scholar]
- Phielix, E.; Szendroedi, J.; Roden, M. Mitochondrial function and insulin resistance during aging: A mini-review. Gerontology 2011, 57, 387–396. [Google Scholar]
- Schmid, A.I.; Szendroedi, J.; Chmelik, M.; Krssak, M.; Moser, E.; Roden, M. Liver ATP synthesis is lower and relates to insulin sensitivity in patients with type 2 diabetes. Diabetes Care 2011, 34, 448–453. [Google Scholar]
- Wallace, D.C. Diseases of the mitochondrial DNA. Annu. Rev. Biochem 1992, 61, 1175–1212. [Google Scholar]
- Orrenius, S.; Zhivotovsky, B.; Nicotera, P. Regulation of cell death: the calcium-apoptosis link. Nat. Rev. Mol. Cell Biol 2003, 4, 552–565. [Google Scholar]
- Di Lisa, F.; Bernardi, P. Mitochondria and ischemia-reperfusion injury of the heart: Fixing a hole. Cardiovasc. Res 2006, 70, 191–199. [Google Scholar]
- Javadov, S.; Karmazyn, M. Mitochondrial permeability transition pore opening as an endpoint to initiate cell death and as a putative target for cardioprotection. Cell Physiol. Biochem 2007, 20, 1–22. [Google Scholar]
- Belzer, F.O.; Southard, J.H. Principles of solid-organ preservation by cold storage. Transplantation 1988, 45, 673–676. [Google Scholar]
- Li, C.; Jackson, R.M. Reactive species mechanisms of cellular hypoxia-reoxygenation injury. Am. J. Physiol. Cell Physiol 2002, 282, C227–C241. [Google Scholar]
- Schneeberger, S.; Amberger, A.; Mandl, J.; Hautz, T.; Renz, O.; Obrist, P.; Meusburger, H.; Brandacher, G.; Mark, W.; Strobl, D.; et al. Cold ischemia contributes to the development of chronic rejection and mitochondrial injury after cardiac transplantation. Transpl. Int 2010, 23, 1282–1292. [Google Scholar]
- Wiedemann, D.; Schneeberger, S.; Friedl, P.; Zacharowski, K.; Wick, N.; Boesch, F.; Margreiter, R.; Laufer, G.; Petzelbauer, P.; Semsroth, S. The fibrin-derived peptide Bβ5–42 significantly attenuates ischemia-reperfusion injury in a cardiac transplant model. Transplantation 2010, 89, 824–829. [Google Scholar]
- McGregor, C.G.; McCallum, H.M.; Hannan, J.; Smith, A.F.; Muir, A.L. Long-term effects of cold cardioplegic myocardial protection in the rat. J. Thorac. Cardiovasc. Surg 1984, 87, 913–919. [Google Scholar]
- Aebert, H.; Cornelius, T.; Ehr, T.; Holmer, S.R.; Birnbaum, D.E.; Riegger, G.A.; Schunkert, H. Expression of immediate early genes after cardioplegic arrest and reperfusion. Ann. Thorac. Surg 1997, 63, 1669–1675. [Google Scholar]
- Stringham, J.C.; Southard, J.H.; Hegge, J.; Triemstra, L.; Fields, B.L.; Belzer, F.O. Limitations of heart preservation by cold storage. Transplantation 1992, 53, 287–294. [Google Scholar]
- Hearse, D.J. Myocardial protection during ischemia and reperfusion. Mol. Cell Biochem 1998, 186, 177–184. [Google Scholar]
- Reichenspurner, H.; Russ, C.; Uberfuhr, P.; Nollert, G.; Schluter, A.; Reichart, B.; Klövekorn, W.P.; Schüler, S.; Hetzer, R.; Brett, W.; et al. Myocardial preservation using HTK solution for heart transplantation. A multicenter study. Eur. J. Cardiothorac. Surg 1993, 7, 414–419. [Google Scholar]
- Ku, K.; Oku, H.; Alam, M.S.; Saitoh, Y.; Nosaka, S.; Nakayama, K. Prolonged hypothermic cardiac storage with histidine-tryptophan-ketoglutarate solution: Comparison with glucose-insulin-potassium and University of Wisconsin solutions. Transplantation 1997, 64, 971–975. [Google Scholar]
- Kuznetsov, A.V.; Kunz, W.S.; Saks, V.; Usson, Y.; Mazat, J.P.; Letellier, T.; Gellerich, F.N.; Margreiter, R. Cryopreservation of mitochondria and mitochondrial function in cardiac and skeletal muscle fibers. Anal. Biochem 2003, 319, 296–303. [Google Scholar]
- Kuznetsov, A.V.; Schneeberger, S.; Seiler, R.; Brandacher, G.; Mark, W.; Steurer, W.; Saks, V.; Usson, Y.; Margreiter, R.; Gnaiger, E. Mitochondrial defects and heterogeneous cytochrome c release after cardiac cold ischemia and reperfusion. Am. J. Physiol. Heart Circ. Physiol 2004, 286, H1633–H1641. [Google Scholar]
- Kay, L.; Daneshrad, Z.; Saks, V.A.; Rossi, A. Alteration in the control of mitochondrial respiration by outer mitochondrial membrane and creatine during heart preservation. Cardiovasc. Res 1997, 34, 547–556. [Google Scholar]
- Fyfe, B.; Loh, E.; Winters, G.L.; Couper, G.S.; Kartashov, A.I.; Schoen, F.J. Heart transplantation-associated perioperative ischemic myocardial injury. Morphological features and clinical significance. Circulation 1996, 93, 1133–1140. [Google Scholar]
- See, Y.P.; Weisel, R.D.; Mickle, D.A.; Teoh, K.H.; Wilson, G.J.; Tumiati, L.C.; Mohabeer, M.K.; Madonik, M.M.; Axford-Gatley, R.A.; Salter, D.R. Prolonged hypothermic cardiac storage for transplantation. The effects on myocardial metabolism and mitochondrial function. J. Thorac. Cardiovasc. Surg 1992, 104, 817–824. [Google Scholar]
- Schneeberger, S.; Kuznetsov, A.V.; Seiler, R.; Renz, O.; Meusburger, H.; Mark, W.; Brandacher, G.; Margreiter, R.; Gnaiger, E. Mitochondrial ischemia-reperfusion injury of the transplanted rat heart: Improved protection by preservation versus cardioplegic solutions. Shock 2008, 30, 365–371. [Google Scholar]
- Sommer, S.P.; Sommer, S.; Sinha, B.; Wiedemann, J.; Otto, C.; Aleksic, I.; Schimmer, C.; Leyh, R.G. Ischemia-reperfusion injury-induced pulmonary mitochondrial damage. J. Heart Lung Transplant 2011, 30, 811–818. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wiedemann, D.; Schachner, T.; Bonaros, N.; Dorn, M.; Andreas, M.; Kocher, A.; Kuznetsov, A.V. Impact of Cold Ischemia on Mitochondrial Function in Porcine Hearts and Blood Vessels. Int. J. Mol. Sci. 2013, 14, 22042-22051. https://doi.org/10.3390/ijms141122042
Wiedemann D, Schachner T, Bonaros N, Dorn M, Andreas M, Kocher A, Kuznetsov AV. Impact of Cold Ischemia on Mitochondrial Function in Porcine Hearts and Blood Vessels. International Journal of Molecular Sciences. 2013; 14(11):22042-22051. https://doi.org/10.3390/ijms141122042
Chicago/Turabian StyleWiedemann, Dominik, Thomas Schachner, Nikolaos Bonaros, Melissa Dorn, Martin Andreas, Alfred Kocher, and Andrey V. Kuznetsov. 2013. "Impact of Cold Ischemia on Mitochondrial Function in Porcine Hearts and Blood Vessels" International Journal of Molecular Sciences 14, no. 11: 22042-22051. https://doi.org/10.3390/ijms141122042