Lipid Transport and Metabolism in Healthy and Osteoarthritic Cartilage


Abstract
:1. Introduction
2. Transporting Lipids through the Cartilage
2.1. Osteoarthritis and Lipid Availability
3. Lipids in Normal Cartilage
4. Lipids in Osteoathritic Cartilage
5. Lipids and Chondrocyte Metabolism
6. Conclusions
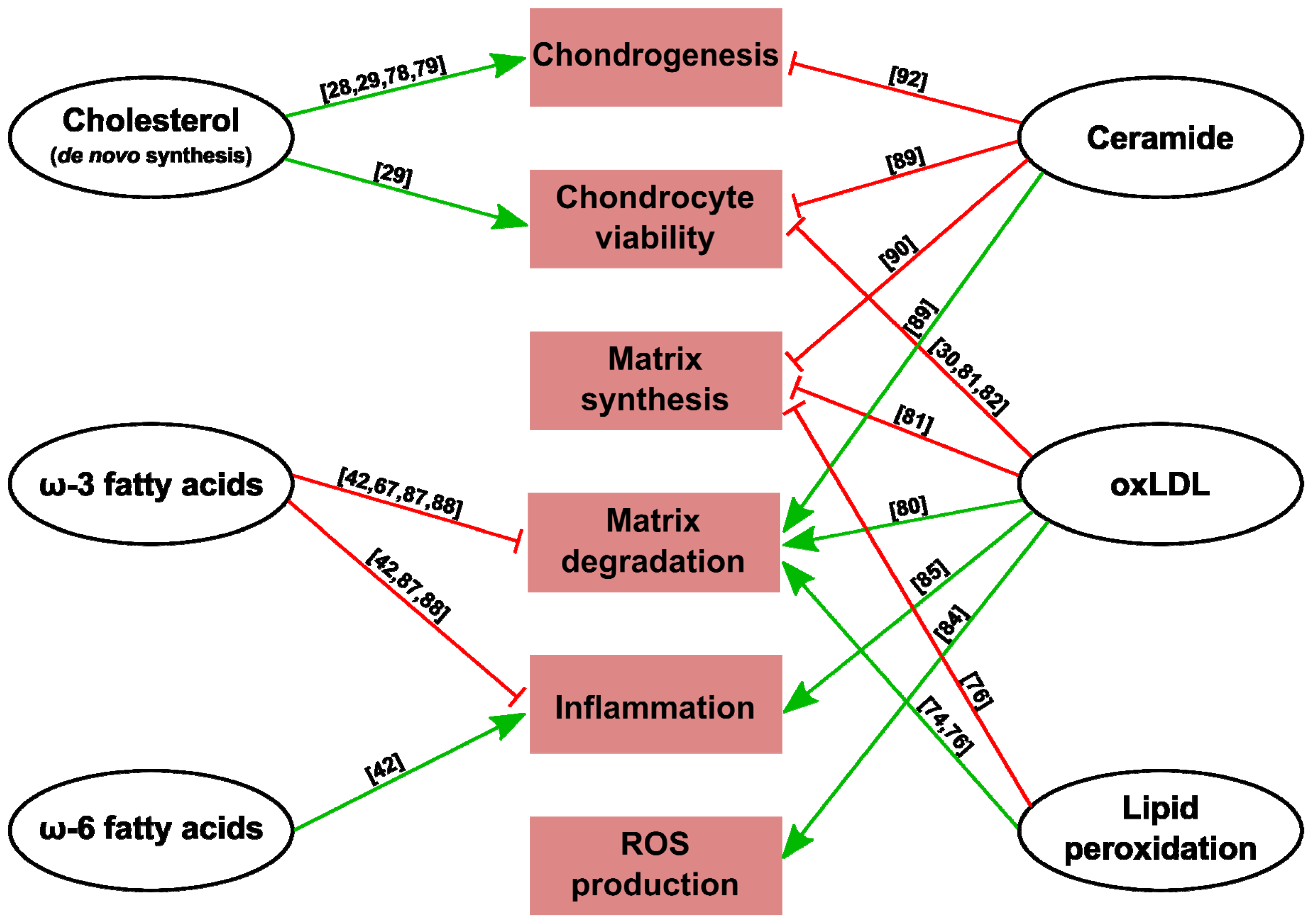

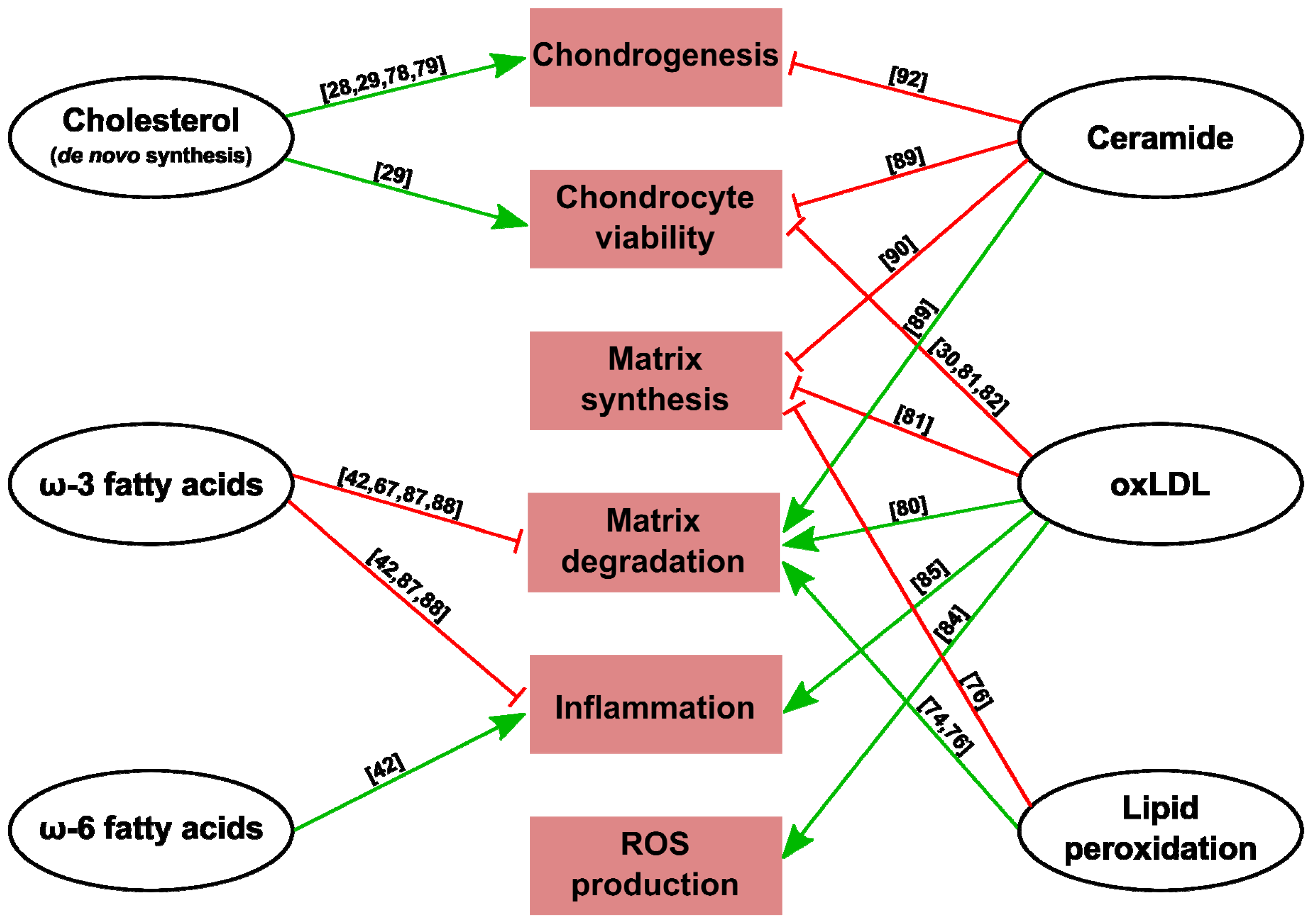{kind=link}
| Altered cartilage permeability | Events related to OA | References |
|---|---|---|
| Causes | Increased protease activity | [25] |
| Increased subchondral vessels invading calcified cartilage | [27] | |
| Joint immobilization | [22] | |
| Consequences | Loss of matrix components in the joint space | [10] |
| Access of deleterious agents (toxins, immunoglobulins) | [10] | |
| Access of proinflammatory plasma proteins | [26] | |
Acknowledgments
Conflicts of Interest
References
- Stockwell, R.A. Lipid content of human costal and articular cartilage. Ann. Rheum. Dis 1967, 26, 481–486. [Google Scholar]
- Otte, P. Basic cell metabolism of articular cartilage. Manometric studies. Z. Rheumatol 1991, 50, 304–312. [Google Scholar]
- Gkretsi, V.; Simopoulou, T.; Tsezou, A. Lipid metabolism and osteoarthritis: Lessons from atherosclerosis. Prog. Lipid Res 2011, 50, 133–140. [Google Scholar]
- Bernstein, P.; Sticht, C.; Jacobi, A.; Liebers, C.; Manthey, S.; Stiehler, M. Expression pattern differences between osteoarthritic chondrocytes and mesenchymal stem cells during chondrogenic differentiation. Osteoarthr. Cartil 2010, 18, 1596–1607. [Google Scholar]
- Ghadially, F.N.; Mehta, P.N.; Kirkaldy-Willis, W.H. Ultrastructure of articular cartilage in experimentally produced lipoarthrosis. J. Bone Jt. Surg. Am 1970, 52, 1147–1158. [Google Scholar]
- Roughley, P.J.; Lee, E.R. Cartilage proteoglycans: Structure and potential functions. Microsc. Res. Tech 1994, 28, 385–397. [Google Scholar]
- Cohen, N.P.; Foster, R.J.; Mow, V.C. Composition and dynamics of articular cartilage: Structure, function, and maintaining healthy state. J. Orthop. Sports Phys. Ther 1998, 28, 203–215. [Google Scholar]
- Maroudas, A. Transport of solutes through cartilage: Permeability to large molecules. J. Anat 1976, 122, 335–347. [Google Scholar]
- Gonsalves, M.; Macpherson, J.V.; O’Hare, D.; Winlove, C.P.; Unwin, P.R. High resolution imaging of the distribution and permeability of methyl viologen dication in bovine articular cartilage using scanning electrochemical microscopy. Biochim. Biophys. Acta 2001, 1524, 66–74. [Google Scholar]
- Lotke, P.A.; Granda, J.L. Alterations in the permeability of articular cartilage by proteolytic enzymes. Arthritis Rheum 1972, 15, 302–308. [Google Scholar]
- Arkill, K.P.; Winlove, C.P. Fatty acid transport in articular cartilage. Arch. Biochem. Biophys 2006, 456, 71–78. [Google Scholar]
- Torzilli, P.A.; Grande, D.A.; Arduino, J.M. Diffusive properties of immature articular cartilage. J. Biomed. Mater. Res 1998, 40, 132–138. [Google Scholar]
- Honner, R.; Thompson, R.C. The nutritional pathways of articular cartilage. An autoradiographic study in rabbits using 35S injected intravenously. J. Bone Jt. Surg. Am 1971, 53, 742–748. [Google Scholar]
- Hodge, J.A.; McKibbin, B. The nutrition of mature and immature cartilage in rabbits. An autoradiographic study. J. Bone Jt. Surg. Br 1969, 51, 140–147. [Google Scholar]
- Ogata, K.; Whiteside, L.A.; Lesker, P.A. Subchondral route for nutrition to articular cartilage in the rabbit. Measurement of diffusion with hydrogen gas in vivo. J. Bone Jt. Surg. Am. 1978, 60, 905–910. [Google Scholar]
- Pan, J.; Zhou, X.; Li, W.; Novotny, J.E.; Doty, S.B.; Wang, L. In situ measurement of transport between subchondral bone and articular cartilage. J. Orthop. Res 2009, 27, 1347–1352. [Google Scholar]
- Arkill, K.P.; Winlove, C.P. Solute transport in the deep and calcified zones of articular cartilage. Osteoarthr. Cartil 2008, 16, 708–714. [Google Scholar]
- Greenwald, A.S.; Haynes, D.W. A pathway for nutrients from the medullary cavity to the articular cartilage of the human femoral head. J. Bone Jt. Surg. Br 1969, 51, 747–753. [Google Scholar]
- Lyons, T.J.; McClure, S.F.; Stoddart, R.W.; McClure, J. The normal human chondro-osseous junctional region: Evidence for contact of uncalcified cartilage with subchondral bone and marrow spaces. BMC Musculoskelet. Disord 2006. [Google Scholar] [CrossRef]
- Wang, Y.; Wei, L.; Zeng, L.; He, D.; Wei, X. Nutrition and degeneration of articular cartilage. Knee Surg. Sports Traumatol. Arthrosc 2012, 21, 1751–1762. [Google Scholar]
- Levick, J.R. Microvascular architecture and exchange in synovial joints. Microcirculation 1995, 2, 217–233. [Google Scholar]
- Maroudas, A.; Bullough, P.; Swanson, S.A.; Freeman, M.A. The permeability of articular cartilage. J. Bone Jt. Surg. Br 1968, 50, 166–177. [Google Scholar]
- Garcia, A.M.; Frank, E.H.; Grimshaw, P.E.; Grodzinsky, A.J. Contributions of fluid convection and electrical migration to transport in cartilage: Relevance to loading. Arch. Biochem. Biophys 1996, 333, 317–325. [Google Scholar]
- O’Hara, B.P.; Urban, J.P.; Maroudas, A. Influence of cyclic loading on the nutrition of articular cartilage. Ann. Rheum. Dis 1990, 49, 536–539. [Google Scholar]
- Appleton, C.T.G.; Pitelka, V.; Henry, J.; Beier, F. Global analyses of gene expression in early experimental osteoarthritis. Arthritis Rheum 2007, 56, 1854–1868. [Google Scholar]
- Sohn, D.H.; Sokolove, J.; Sharpe, O.; Erhart, J.C.; Chandra, P.E.; Lahey, L.J.; Lindstrom, T.M.; Hwang, I.; Boyer, K.A.; Andriacchi, T.P.; et al. Plasma proteins present in osteoarthritic synovial fluid can stimulate cytokine production via Toll-like receptor 4. Arthritis Res. Ther 2012. [Google Scholar] [CrossRef]
- Pan, J.; Wang, B.; Li, W.; Zhou, X.; Scherr, T.; Yang, Y.; Price, C.; Wang, L. Elevated cross-talk between subchondral bone and cartilage in osteoarthritic joints. Bone 2012, 51, 212–217. [Google Scholar]
- Wu, S.; de Luca, F. Role of cholesterol in the regulation of growth plate chondrogenesis and longitudinal bone growth. J. Biol. Chem 2004, 279, 4642–4647. [Google Scholar]
- Aguilar, A.; Wu, S.; de Luca, F. P450 oxidoreductase expressed in rat chondrocytes modulates chondrogenesis via cholesterol- and Indian hedgehog-dependent mechanisms. Endocrinology 2009, 150, 2732–2739. [Google Scholar]
- Simopoulou, T.; Malizos, K.N.; Tsezou, A. Lectin-like oxidized low density lipoprotein receptor 1 (LOX-1) expression in human articular chondrocytes. Clin. Exp. Rheumatol 2007, 25, 605–612. [Google Scholar]
- Tsezou, A.; Iliopoulos, D.; Malizos, K.N.; Simopoulou, T. Impaired expression of genes regulating cholesterol efflux in human osteoarthritic chondrocytes. J. Orthop. Res 2010, 28, 1033–1039. [Google Scholar]
- Prete, P.E.; Gurakar-Osborne, A.; Kashyap, M.L. Synovial fluid lipids and apolipoproteins: A contemporary perspective. Biorheology 1995, 32, 1–16. [Google Scholar]
- Oliviero, F.; Lo Nigro, A.; Bernardi, D.; Giunco, S.; Baldo, G.; Scanu, A.; Sfriso, P.; Ramonda, R.; Plebani, M.; Punzi, L. A comparative study of serum and synovial fluid lipoprotein levels in patients with various arthritides. Clin. Chim. Acta 2012, 413, 303–307. [Google Scholar]
- Cillero-Pastor, B.; Eijkel, G.; Kiss, A.; Blanco, F.J.; Heeren, R.M.A. Time-of-flight secondary ion mass spectrometry-based molecular distribution distinguishing healthy and osteoarthritic human cartilage. Anal. Chem 2012, 84, 8909–8916. [Google Scholar]
- Gabay, O.; Sanchez, C.; Salvat, C.; Chevy, F.; Breton, M.; Nourissat, G.; Wolf, C.; Jacques, C.; Berenbaum, F. Stigmasterol: A phytosterol with potential anti-osteoarthritic properties. Osteoarthr. Cartil 2010, 18, 106–116. [Google Scholar]
- Lippiello, L.; Walsh, T.; Fienhold, M. The association of lipid abnormalities with tissue pathology in human osteoarthritic articular cartilage. Metabolism 1991, 40, 571–576. [Google Scholar]
- Cleland, K.A.; James, M.J.; Neumann, M.A.; Gibson, R.A.; Cleland, L.G. Differences in fatty acid composition of immature and mature articular cartilage in humans and sheep. Lipids 1995, 30, 949–953. [Google Scholar]
- Le Lous, M.; Corvol, M.T.; Maroteaux, P. Lipid composition of two types of chondrocytes in primary culture. Calcif. Tissue Int 1981, 33, 403–407. [Google Scholar]
- Lippiello, L.; Fienhold, M.; Grandjean, C. Metabolic and ultrastructural changes in articular cartilage of rats fed dietary supplements of omega-3 fatty acids. Arthritis Rheum 1990, 33, 1029–1036. [Google Scholar]
- Xu, H.; Watkins, B.A.; Adkisson, H.D. Dietary lipids modify the fatty acid composition of cartilage, isolated chondrocytes and matrix vesicles. Lipids 1994, 29, 619–625. [Google Scholar]
- Nagao, M.; Ishii, S.; Murata, Y.; Akino, T. Effect of extracellular fatty acids on lipid metabolism in cultured rabbit articular chondrocytes. J. Orthop. Res 1991, 9, 341–347. [Google Scholar]
- Lopez, H.L. Nutritional interventions to prevent and treat osteoarthritis. Part I: Focus on fatty acids and macronutrients. PMR 2012, 4, S145–S154. [Google Scholar]
- Alvarez-Soria, M.A.; Largo, R.; Santillana, J.; Sánchez-Pernaute, O.; Calvo, E.; Hernández, M.; Egido, J.; Herrero-Beaumont, G. Long term NSAID treatment inhibits COX-2 synthesis in the knee synovial membrane of patients with osteoarthritis: Differential proinflammatory cytokine profile between celecoxib and aceclofenac. Ann. Rheum. Dis 2006, 65, 998–1005. [Google Scholar]
- Alvarez-Soria, M.A.; Herrero-Beaumont, G.; Moreno-Rubio, J.; Calvo, E.; Santillana, J.; Egido, J.; Largo, R. Long-term NSAID treatment directly decreases COX-2 and mPGES-1 production in the articular cartilage of patients with osteoarthritis. Osteoarthr. Cartil 2008, 16, 1484–1493. [Google Scholar]
- Simopoulos, A.P. The importance of the ω-6/ω-3 fatty acid ratio in cardiovascular disease and other chronic diseases. Exp. Biol. Med 2008, 233, 674–688. [Google Scholar]
- Sarma, A.V.; Powell, G.L.; LaBerge, M. Phospholipid composition of articular cartilage boundary lubricant. J. Orthop. Res 2001, 19, 671–676. [Google Scholar]
- Nagao, M.; Ishii, S.; Kitamura, K.; Akino, T. Arachidonic acid metabolism in articular chondrocytes. Clin. Orthop. Relat. Res 1991, 271, 288–295. [Google Scholar]
- Zhuo, Q.; Yang, W.; Chen, J.; Wang, Y. Metabolic syndrome meets osteoarthritis. Nat. Rev. Rheumatol 2012, 8, 729–737. [Google Scholar]
- Al-Arfaj, A.S. Radiographic osteoarthritis and serum cholesterol. Saudi Med. J 2003, 24, 745–747. [Google Scholar]
- Stürmer, T.; Sun, Y.; Sauerland, S.; Zeissig, I.; Günther, K.P.; Puhl, W.; Brenner, H. Serum cholesterol and osteoarthritis. The baseline examination of the Ulm Osteoarthritis Study. J. Rheumatol 1998, 25, 1827–1832. [Google Scholar]
- Hart, D.J.; Doyle, D.V.; Spector, T.D. Association between metabolic factors and knee osteoarthritis in women: The Chingford Study. J. Rheumatol 1995, 22, 1118–1123. [Google Scholar]
- Conaghan, P.G. The effects of statins on osteoarthritis structural progression: Another glimpse of the Holy Grail? Ann. Rheum. Dis 2012, 71, 633–634. [Google Scholar]
- Clockaerts, S.; van Osch, G.J.V.M.; Bastiaansen-Jenniskens, Y.M.; Verhaar, J.A.N.; van Glabbeek, F.; van Meurs, J.B.; Kerkhof, H.J.M.; Hofman, A.; Stricker, B.H.C.; Bierma-Zeinstra, S.M. Statin use is associated with reduced incidence and progression of knee osteoarthritis in the Rotterdam study. Ann. Rheum. Dis 2012, 71, 642–647. [Google Scholar]
- Kadam, U.T.; Blagojevic, M.; Belcher, J. Statin use and clinical osteoarthritis in the general population: A longitudinal study. J. Gen. Intern. Med 2013, 28, 943–949. [Google Scholar]
- Riddle, D.L.; Moxley, G.; Dumenci, L. Associations between statin use and changes in pain, function and structural progression: A longitudinal study of persons with knee osteoarthritis. Ann. Rheum. Dis 2013, 72, 196–203. [Google Scholar]
- Beattie, M.S.; Lane, N.E.; Hung, Y.-Y.; Nevitt, M.C. Association of statin use and development and progression of hip osteoarthritis in elderly women. J. Rheumatol 2005, 32, 106–110. [Google Scholar]
- Aktas, E.; Sener, E.; Gocun, P.U. Mechanically induced experimental knee osteoarthritis benefits from anti-inflammatory and immunomodulatory properties of simvastatin via inhibition of matrix metalloproteinase-3. J. Orthop. Traumatol 2011, 12, 145–151. [Google Scholar]
- Yudoh, K.; Karasawa, R. Statin prevents chondrocyte aging and degeneration of articular cartilage in osteoarthritis (OA). Aging 2010, 2, 990–998. [Google Scholar]
- Dombrecht, E.J.; van Offel, J.F.; Bridts, C.H.; Ebo, D.G.; Seynhaeve, V.; Schuerwegh, A.J.; Stevens, W.J.; de Clerck, L.S. Influence of simvastatin on the production of pro-inflammatory cytokines and nitric oxide by activated human chondrocytes. Clin. Exp. Rheumatol 2007, 25, 534–539. [Google Scholar]
- Barter, M.J.; Hui, W.; Lakey, R.L.; Catterall, J.B.; Cawston, T.E.; Young, D.A. Lipophilic statins prevent matrix metalloproteinase-mediated cartilage collagen breakdown by inhibiting protein geranylgeranylation. Ann. Rheum. Dis 2010, 69, 2189–2198. [Google Scholar]
- Lazzerini, P.E.; Capecchi, P.L.; Nerucci, F.; Fioravanti, A.; Chellini, F.; Piccini, M.; Bisogno, S.; Marcolongo, R.; Laghi Pasini, F. Simvastatin reduces MMP-3 level in interleukin 1β stimulated human chondrocyte culture. Ann. Rheum. Dis 2004, 63, 867–869. [Google Scholar]
- Sverdrup, F.M.; Yates, M.P.; Vickery, L.E.; Klover, J.A.; Song, L.R.-H.; Anglin, C.P.; Misko, T.P. Protein geranylgeranylation controls collagenase expression in osteoarthritic cartilage. Osteoarthr. Cartil 2010, 18, 948–955. [Google Scholar]
- Stammers, T.; Sibbald, B.; Freeling, P. Efficacy of cod liver oil as an adjunct to non-steroidal anti-inflammatory drug treatment in the management of osteoarthritis in general practice. Ann. Rheum. Dis 1992, 51, 128–129. [Google Scholar]
- Iacono, A.; Gómez, R.; Sperry, J.; Conde, J.; Bianco, G.; Meli, R.; Gómez-Reino, J.J.; Smith, A.B.; Gualillo, O. Effect of oleocanthal and its derivatives on inflammatory response induced by lipopolysaccharide in a murine chondrocyte cell line. Arthritis Rheum 2010, 62, 1675–1682. [Google Scholar]
- Wang, Y.; Wluka, A.E.; Hodge, A.M.; English, D.R.; Giles, G.G.; O’Sullivan, R.; Cicuttini, F.M. Effect of fatty acids on bone marrow lesions and knee cartilage in healthy, middle-aged subjects without clinical knee osteoarthritis. Osteoarthr. Cartil 2008, 16, 579–583. [Google Scholar]
- Baker, K.R.; Matthan, N.R.; Lichtenstein, A.H.; Niu, J.; Guermazi, A.; Roemer, F.; Grainger, A.; Nevitt, M.C.; Clancy, M.; Lewis, C.E.; et al. Association of plasma n-6 and n-3 polyunsaturated fatty acids with synovitis in the knee: The MOST study. Osteoarthr. Cartil 2012, 20, 382–387. [Google Scholar]
- Knott, L.; Avery, N.C.; Hollander, A.P.; Tarlton, J.F. Regulation of osteoarthritis by ω-3 (n-3) polyunsaturated fatty acids in a naturally occurring model of disease. Osteoarthr. Cartil 2011, 19, 1150–1157. [Google Scholar]
- Huang, M.-J.; Wang, L.; Jin, D.; Zhang, Z.-M.; Chen, T.-Y.; Jia, C.-H.; Wang, Y.; Zhen, X.-C.; Huang, B.; Yan, B.; et al. Enhancement of the synthesis of n-3 PUFAs in fat-1 transgenic mice inhibits mTORC1 signalling and delays surgically induced osteoarthritis in comparison with wild-type mice. Ann. Rheum. Dis 2013. [Google Scholar] [CrossRef]
- Kosinska, M.K.; Liebisch, G.; Lochnit, G.; Wilhelm, J.; Klein, H.; Kaesser, U.; Lasczkowski, G.; Rickert, M.; Schmitz, G.; Steinmeyer, J. A lipidomic study of phospholipid classes and species in human synovial fluid. Arthritis Rheum 2013, 65, 2323–2333. [Google Scholar]
- Parks, T.P.; Lukas, S.; Hoffman, A.F. Purification and characterization of a phospholipase A2 from human osteoarthritic synovial fluid. Adv. Exp. Med. Biol 1990, 275, 55–81. [Google Scholar]
- Pruzanski, W.; Bogoch, E.; Stefanski, E.; Wloch, M.; Vadas, P. Enzymatic activity and distribution of phospholipase A2 in human cartilage. Life Sci 1991, 48, 2457–2462. [Google Scholar]
- Chang, J.; Gilman, S.C.; Lewis, A.J. Interleukin 1 activates phospholipase A2 in rabbit chondrocytes: A possible signal for IL 1 action. J. Immunol 1986, 136, 1283–1287. [Google Scholar]
- Triantaphyllidou, I.-E.; Kalyvioti, E.; Karavia, E.; Lilis, I.; Kypreos, K.E.; Papachristou, D.J. Perturbations in the HDL metabolic pathway predispose to the development of osteoarthritis in mice following long-term exposure to western-type diet. Osteoarthr. Cartil 2013, 21, 322–330. [Google Scholar]
- Tiku, M.L.; Shah, R.; Allison, G.T. Evidence linking chondrocyte lipid peroxidation to cartilage matrix protein degradation. Possible role in cartilage aging and the pathogenesis of osteoarthritis. J. Biol. Chem 2000, 275, 20069–20076. [Google Scholar]
- Shah, R.; Raska, K.; Tiku, M.L. The presence of molecular markers of in vivo lipid peroxidation in osteoarthritic cartilage: A pathogenic role in osteoarthritis. Arthritis Rheum 2005, 52, 2799–2807. [Google Scholar]
- Morquette, B.; Shi, Q.; Lavigne, P.; Ranger, P.; Fernandes, J.C.; Benderdour, M. Production of lipid peroxidation products in osteoarthritic tissues: New evidence linking 4-hydroxynonenal to cartilage degradation. Arthritis Rheum 2006, 54, 271–281. [Google Scholar]
- Roman-Blas, J.A.; Contreras-Blasco, M.A.; Largo, R.; Alvarez-Soria, M.A.; Castañeda, S.; Herrero-Beaumont, G. Differential effects of the antioxidant N-acetylcysteine on the production of catabolic mediators in IL-1β-stimulated human osteoarthritic synoviocytes and chondrocytes. Eur. J. Pharmacol 2009, 623, 125–131. [Google Scholar]
- Gentili, C.; Tutolo, G.; Pianezzi, A.; Cancedda, R.; Descalzi Cancedda, F. Cholesterol secretion and homeostasis in chondrocytes: A liver X receptor and retinoid X receptor heterodimer mediates apolipoprotein A1 expression. Matrix Biol 2005, 24, 35–44. [Google Scholar]
- Woods, A.; James, C.G.; Wang, G.; Dupuis, H.; Beier, F. Control of chondrocyte gene expression by actin dynamics: A novel role of cholesterol/Ror-α signalling in endochondral bone growth. J. Cell. Mol. Med 2009, 13, 3497–3516. [Google Scholar]
- Kakinuma, T.; Yasuda, T.; Nakagawa, T.; Hiramitsu, T.; Akiyoshi, M.; Akagi, M.; Sawamura, T.; Nakamura, T. Lectin-like oxidized low-density lipoprotein receptor 1 mediates matrix metalloproteinase 3 synthesis enhanced by oxidized low-density lipoprotein in rheumatoid arthritis cartilage. Arthritis Rheum 2004, 50, 3495–3503. [Google Scholar]
- Akagi, M.; Kanata, S.; Mori, S.; Itabe, H.; Sawamura, T.; Hamanishi, C. Possible involvement of the oxidized low-density lipoprotein/lectin-like oxidized low-density lipoprotein receptor-1 system in pathogenesis and progression of human osteoarthritis. Osteoarthr. Cartil 2007, 15, 281–290. [Google Scholar]
- Zushi, S.; Akagi, M.; Kishimoto, H.; Teramura, T.; Sawamura, T.; Hamanishi, C. Induction of bovine articular chondrocyte senescence with oxidized low-density lipoprotein through lectin-like oxidized low-density lipoprotein receptor 1. Arthritis Rheum 2009, 60, 3007–3016. [Google Scholar]
- Kishimoto, H.; Akagi, M.; Zushi, S.; Teramura, T.; Onodera, Y.; Sawamura, T.; Hamanishi, C. Induction of hypertrophic chondrocyte-like phenotypes by oxidized LDL in cultured bovine articular chondrocytes through increase in oxidative stress. Osteoarthr. Cartil 2010, 18, 1284–1290. [Google Scholar]
- Nishimura, S.; Akagi, M.; Yoshida, K.; Hayakawa, S.; Sawamura, T.; Munakata, H.; Hamanishi, C. Oxidized low-density lipoprotein (ox-LDL) binding to lectin-like ox-LDL receptor-1 (LOX-1) in cultured bovine articular chondrocytes increases production of intracellular reactive oxygen species (ROS) resulting in the activation of NF-κB. Osteoarthr. Cartil 2004, 12, 568–576. [Google Scholar]
- Akagi, M.; Ueda, A.; Teramura, T.; Kanata, S.; Sawamura, T.; Hamanishi, C. Oxidized LDL binding to LOX-1 enhances MCP-1 expression in cultured human articular chondrocytes. Osteoarthr. Cartil 2009, 17, 271–275. [Google Scholar]
- Lippiello, L. Lipid and cell metabolic changes associated with essential fatty acid enrichment of articular chondrocytes. Proc. Soc. Exp. Biol. Med 1990, 195, 282–287. [Google Scholar]
- Curtis, C.L.; Hughes, C.E.; Flannery, C.R.; Little, C.B.; Harwood, J.L.; Caterson, B. N-3 fatty acids specifically modulate catabolic factors involved in articular cartilage degradation. J. Biol. Chem 2000, 275, 721–724. [Google Scholar]
- Zainal, Z.; Longman, A.J.; Hurst, S.; Duggan, K.; Caterson, B.; Hughes, C.E.; Harwood, J.L. Relative efficacies of ω-3 polyunsaturated fatty acids in reducing expression of key proteins in a model system for studying osteoarthritis. Osteoarthr. Cartil 2009, 17, 896–905. [Google Scholar]
- Sabatini, M.; Rolland, G.; Léonce, S.; Thomas, M.; Lesur, C.; Pérez, V.; de Nanteuil, G.; Bonnet, J. Effects of ceramide on apoptosis, proteoglycan degradation, and matrix metalloproteinase expression in rabbit articular cartilage. Biochem. Biophys. Res. Commun 2000, 267, 438–444. [Google Scholar]
- Gilbert, S.J.; Blain, E.J.; Duance, V.C.; Mason, D.J. Sphingomyelinase decreases type II collagen expression in bovine articular cartilage chondrocytes via the ERK signaling pathway. Arthritis Rheum 2008, 58, 209–220. [Google Scholar]
- Gilbert, S.J.; Blain, E.J.; Jones, P.; Duance, V.C.; Mason, D.J. Exogenous sphingomyelinase increases collagen and sulphated glycosaminoglycan production by primary articular chondrocytes: An in vitro study. Arthritis Res. Ther 2006. [Google Scholar] [CrossRef]
- Simonaro, C.M.; Sachot, S.; Ge, Y.; He, X.; Deangelis, V.A.; Eliyahu, E.; Leong, D.J.; Sun, H.B.; Mason, J.B.; Haskins, M.E.; et al. Acid ceramidase maintains the chondrogenic phenotype of expanded primary chondrocytes and improves the chondrogenic differentiation of bone marrow-derived mesenchymal stem cells. PLoS One 2013, 8, e62715. [Google Scholar]
- Laplante, M.; Sabatini, D.M. mTOR signaling at a glance. J. Cell Sci 2009, 122, 3589–3594. [Google Scholar]
- André, C.; Cota, D. Coupling nutrient sensing to metabolic homoeostasis: The role of the mammalian target of rapamycin complex 1 pathway. Proc. Nutr. Soc 2012, 71, 502–510. [Google Scholar]
- Mordier, S.; Iynedjian, P.B. Activation of mammalian target of rapamycin complex 1 and insulin resistance induced by palmitate in hepatocytes. Biochem. Biophys. Res. Commun 2007, 362, 206–211. [Google Scholar]
- Rivas, D.A.; Yaspelkis, B.B.; Hawley, J.A.; Lessard, S.J. Lipid-induced mTOR activation in rat skeletal muscle reversed by exercise and 5′-aminoimidazole-4-carboxamide-1-β-d-ribofuranoside. J. Endocrinol 2009, 202, 441–451. [Google Scholar]
- Wen, Z.-H.; Su, Y.-C.; Lai, P.-L.; Zhang, Y.; Xu, Y.-F.; Zhao, A.; Yao, G.-Y.; Jia, C.-H.; Lin, J.; Xu, S.; et al. Critical role of arachidonic acid-activated mTOR signaling in breast carcinogenesis and angiogenesis. Oncogene 2013, 32, 160–170. [Google Scholar]
- Pattingre, S.; Bauvy, C.; Levade, T.; Levine, B.; Codogno, P. Ceramide-induced autophagy: To junk or to protect cells? Autophagy 2009, 5, 558–560. [Google Scholar]
- Mathis, A.S.; Jin, S.; Friedman, G.S.; Peng, F.; Carl, S.M.; Knipp, G.T. The pharmacodynamic effects of sirolimus and sirolimus-calcineurin inhibitor combinations on macrophage scavenger and nuclear hormone receptors. J. Pharm. Sci 2007, 96, 209–222. [Google Scholar]
- Xu, J.; Dang, Y.; Ren, Y.R.; Liu, J.O. Cholesterol trafficking is required for mTOR activation in endothelial cells. Proc. Natl. Acad. Sci. USA 2010, 107, 4764–4769. [Google Scholar]
- Srinivas, V.; Bohensky, J.; Shapiro, I.M. Autophagy: A new phase in the maturation of growth plate chondrocytes is regulated by HIF, mTOR and AMP kinase. Cells Tissues Organs 2009, 189, 88–92. [Google Scholar]
- Phornphutkul, C.; Wu, K.-Y.; Auyeung, V.; Chen, Q.; Gruppuso, P.A. mTOR signaling contributes to chondrocyte differentiation. Dev. Dyn 2008, 237, 702–712. [Google Scholar]
- Caramés, B.; Taniguchi, N.; Otsuki, S.; Blanco, F.J.; Lotz, M. Autophagy is a protective mechanism in normal cartilage, and its aging-related loss is linked with cell death and osteoarthritis. Arthritis Rheum 2010, 62, 791–801. [Google Scholar]
- Caramés, B.; Hasegawa, A.; Taniguchi, N.; Miyaki, S.; Blanco, F.J.; Lotz, M. Autophagy activation by rapamycin reduces severity of experimental osteoarthritis. Ann. Rheum. Dis 2012, 71, 575–581. [Google Scholar]
- Sasaki, H.; Takayama, K.; Matsushita, T.; Ishida, K.; Kubo, S.; Matsumoto, T.; Fujita, N.; Oka, S.; Kurosaka, M.; Kuroda, R. Autophagy modulates osteoarthritis-related gene expression in human chondrocytes. Arthritis Rheum 2012, 64, 1920–1928. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Villalvilla, A.; Gómez, R.; Largo, R.; Herrero-Beaumont, G. Lipid Transport and Metabolism in Healthy and Osteoarthritic Cartilage. Int. J. Mol. Sci. 2013, 14, 20793-20808. https://doi.org/10.3390/ijms141020793
Villalvilla A, Gómez R, Largo R, Herrero-Beaumont G. Lipid Transport and Metabolism in Healthy and Osteoarthritic Cartilage. International Journal of Molecular Sciences. 2013; 14(10):20793-20808. https://doi.org/10.3390/ijms141020793
Chicago/Turabian StyleVillalvilla, Amanda, Rodolfo Gómez, Raquel Largo, and Gabriel Herrero-Beaumont. 2013. "Lipid Transport and Metabolism in Healthy and Osteoarthritic Cartilage" International Journal of Molecular Sciences 14, no. 10: 20793-20808. https://doi.org/10.3390/ijms141020793