BTG1 Expression Correlates with the Pathogenesis and Progression of Ovarian Carcinomas

Abstract
:1. Introduction
2. Results
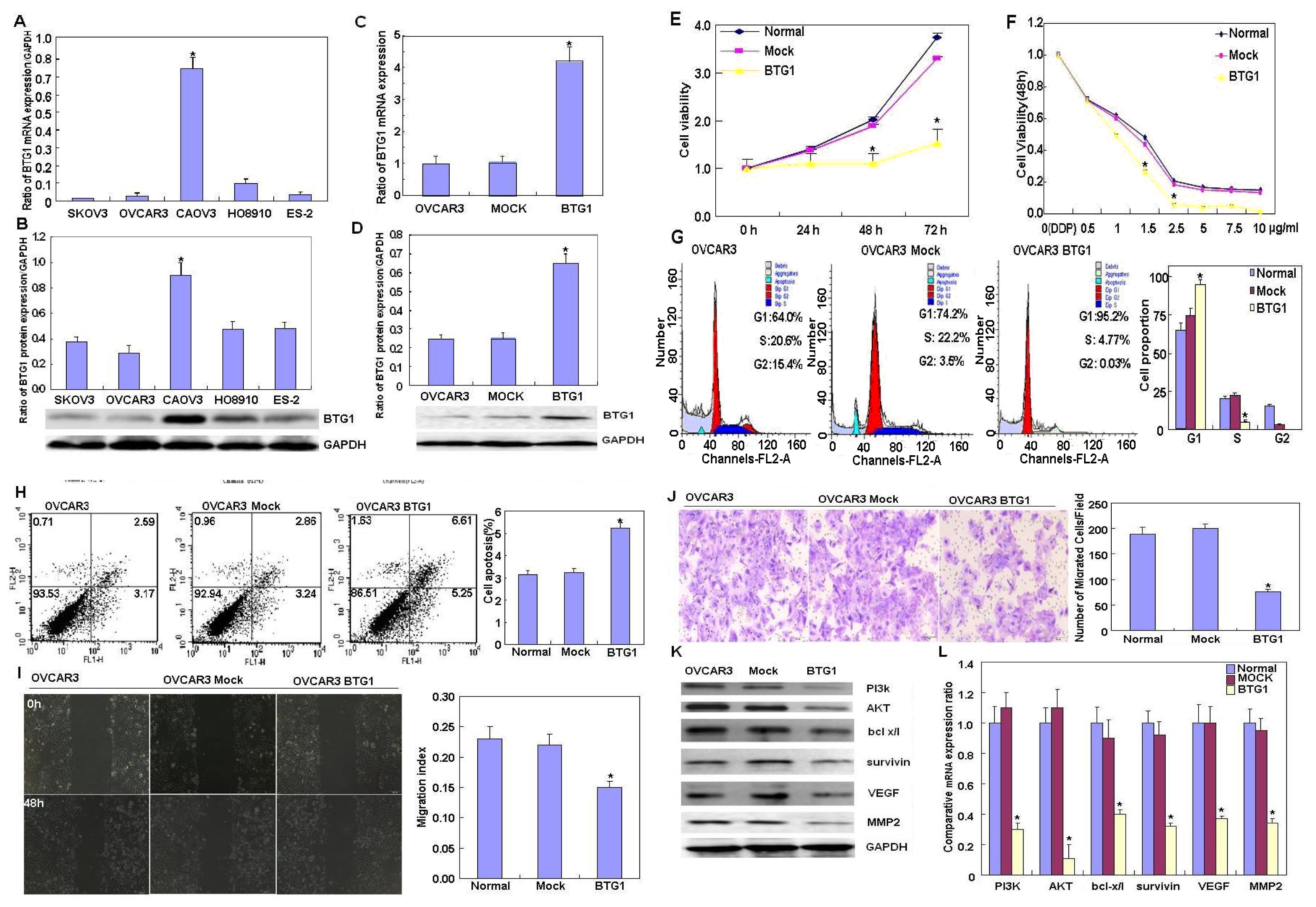
2.1. Effects of BTG1 Overexpression on the Phenotypes and Related Proteins of Ovarian Carcinoma
2.2. Correlation of BTG1 mRNA Expression with Tumorigenesis and Clinicopathological Features of Ovarian Carcinoma
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Plasmid Construction and Transfection
4.3. Cell Cycle Analysis
4.4. Apoptosis Assay
4.5. Wound Healing Assay
4.6. Cell Invasion Assays
4.7. Subjects
4.8. Pathology
4.9. Real-Time RT-PCR
4.10. Western Blotting
4.11. Statistical Analysis
5. Conclusions
Supplementary Information
ijms-14-19670-s001.pdf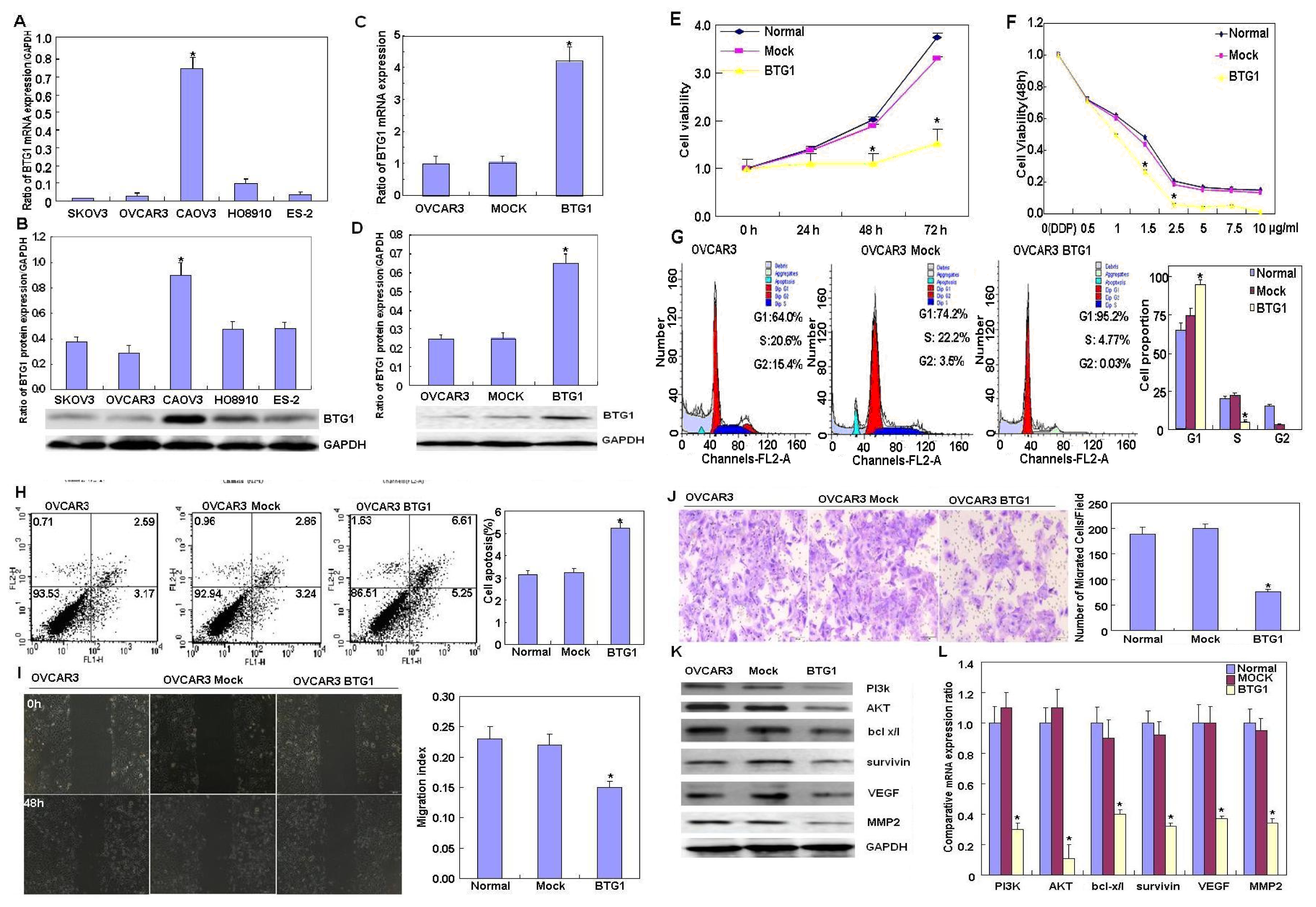

{kind=link}
| Group | n | BTG1 mRNA expression |
|---|---|---|
| Normal | 17 | 0.197 ± 0.080 ** |
| Benign tumors | 12 | 0.127 ± 0.077 * |
| Carcinoma | 64 | 0.077 ± 0.059 |
| Clinicopathological features | n | BTG1 mRNA expression |
|---|---|---|
| Pathological classification | ||
| Serous adenocarcinoma | 50 | 0.076 ± 0.062 |
| Mucinous adenocarcinoma | 7 | 0.069 ± 0.038 |
| Miscellaneous subtypes | 7 | 0.089 ± 0.059 |
| FIGO staging: | ||
| I–II | 30 | 0.093 ± 0.069 * |
| III–IV | 34 | 0.063 ± 0.045 |
| Differentiation | ||
| Well-differentiated | 21 | 0.084 ± 0.046 |
| Moderately differentiated | 22 | 0.076 ± 0.053 |
| Poorly differentiated | 21 | 0.071 ± 0.076 |
Acknowledgements
Conflicts of Interest
References
- Piek, J.M.; van Diest, P.J.; Verheijen, R.H. Ovarian carcinogenesis: An alternative hypothesis. Adv. Exp. Med. Biol 2008, 622, 79–87. [Google Scholar]
- Bandera, C.A. Advances in the understanding of risk factors for ovarian cancer. J. Reprod. Med 2005, 50, 399–406. [Google Scholar]
- Kawamura-Tsuzuku, J.; Suzuki, T.; Yoshida, Y.; Yamamoto, T. Nuclear localization of Tob is important for regulation of its antiproliferative activity. Oncogene 2004, 23, 6630–6638. [Google Scholar]
- Winkler, G.S. The mammalian anti-proliferative BTG/Tob protein family. J. Cell. Physiol 2010, 222, 66–72. [Google Scholar]
- Rodier, A.; Rochard, P.; Berthet, C.; Rouault, J.P.; Casas, F.; Daury, L.; Busson, M.; Magaud, J.P.; Wrutniak-Cabello, C.; Cabello, G. Identification of functional domains involved in BTG1 cell localization. Oncogene 2001, 20, 2691–2703. [Google Scholar]
- Lin, W.J.; Gary, J.D.; Yang, M.C.; Clarke, S.; Herschman, H.R. The mammalian immediate-early TIS21 protein and the leukemia-associated BTG1 protein interact with a protein-arginine N-methyltransferase. J. Biol. Chem 1996, 271, 15034–15044. [Google Scholar]
- Rimokh, R.; Rouault, J.P.; Wahbi, K.; Gadoux, M.; Lafage, M.; Archimbaud, E.; Charrin, C.; Gentilhomme, O.; Germain, D.; Samarut, J. A chromosome 12 coding region is juxtaposed to the MYC protooncogene locus in a t(8;12)(q24;q22) translocation in a case of B-cell chronic lymphocytic leukemia. Genes Chromosomes Cancer 1991, 3, 24–36. [Google Scholar]
- Cho, J.W.; Kim, J.J.; Park, S.G.; Lee, D.H.; Lee, S.C.; Kim, H.J.; Park, B.C.; Cho, S. Identification of B-cell translocation gene 1 as a biomarker for monitoring the remission of acute myeloid leukemia. Proteomics 2004, 4, 3456–3463. [Google Scholar]
- Rouault, J.P.; Rimokh, R.; Tessa, C.; Paranhos, G.; Ffrench, M.; Duret, L.; Garoccio, M.; Germain, D.; Samarut, J.; Magaud, J.P. BTG1, a member of a new family of antiproliferative genes. EMBO J 1992, 11, 1663–1670. [Google Scholar]
- Busson, M.; Carazo, A.; Seyer, P.; Grandemange, S.; Casas, F.; Pessemesse, L.; Rouault, J.P.; Wrutniak-Cabello, C.; Cabello, G. Coactivation of nuclear receptors and myogenic factors induces the major BTG1 influence on muscle differentiation. Oncogene 2005, 24, 1698–1710. [Google Scholar]
- Rouault, J.P.; Prevot, D.; Berthet, C.; Birot, A.M.; Billaud, M.; Magaud, J.P.; Corbo, L. Interaction of BTG1 and p53-regulated BTG2 gene products with mCaf1, the murine homolog of a component of the yeast CCR4 transcriptional regulatory complex. J. Biol. Chem 1998, 273, 22563–22569. [Google Scholar]
- Hata, K.; Nishijima, K.; Mizuguchi, J. Role for Btg1 and Btg2 in growth arrest of WEHI-231 cells through arginine methylation following membrane immunoglobulin engagement. Exp. Cell Res 2007, 313, 2356–2366. [Google Scholar]
- Bakker, W.J.; Blazquez-Domingo, M.; Kolbus, A.; Besooyen, J.; Steinlein, P.; Beug, H.; Coffer, P.J.; Löwenberg, B.; von Lindern, M.; van Dijk, T.B. FoxO3a regulates erythroid differentiation and induces BTG1, an activator of protein arginine methyl transferase 1. J. Cell. Biol 2004, 164, 175–184. [Google Scholar]
- Bogdan, J.A.; Adams-Burton, C.; Pedicord, D.L.; Sukovich, D.A.; Benfield, P.A.; Corjay, M.H.; Stoltenborg, J.K.; Dicker, I.B. Human carbon catabolite repressor protein (CCR4)-associative factor 1: Cloning, expression and characterization of its interaction with the B-cell translocation protein BTG1. Biochem. J 1998, 336, 471–481. [Google Scholar]
- Ikematsu, N.; Yoshida, Y.; Kawamura-Tsuzuku, J.; Ohsugi, M.; Onda, M.; Hirai, M.; Fujimoto, J.; Yamamoto, T. Tob2, a novel anti-proliferative Tob/BTG1 family member, associates with a component of the CCR4 transcriptional regulatory complex capable of binding cyclin-dependent kinases. Oncogene 1999, 18, 7432–7441. [Google Scholar]
- Nahta, R.; Yuan, L.X.; Fiterman, D.J.; Zhang, L.; Symmans, W.F.; Ueno, N.T.; Esteva, F.J. B cell translocation gene 1 contributes to antisense Bcl-2-mediated apoptosis in breast cancer cells. Mol. Cancer Ther 2006, 5, 1593–1601. [Google Scholar]
- Prevot, D.; Voeltzel, T.; Birot, A.M.; Morel, A.P.; Rostan, M.C.; Magaud, J.P.; Corbo, L. The leukemia-associated protein Btg1 and the p53-regulated protein Btg2 interact with the homeoprotein Hoxb9 and enhance its transcriptional activation. J. Biol. Chem 2000, 275, 147–153. [Google Scholar]
- Corjay, M.H.; Kearney, M.A.; Munzer, D.A.; Diamond, S.M.; Stoltenborg, J.K. Antiproliferative gene BTG1 is highly expressed in apoptotic cells in macrophage-rich areas of advanced lesions in Watanabe heritable hyperlipidemic rabbit and human. Lab. Invest 1998, 78, 847–858. [Google Scholar]
- Rodier, A.; Marchal-Victorion, S.; Rochard, P.; Casas, F.; Cassar-Malek, I.; Rouault, J.P.; Magaud, J.P.; Mason, D.Y.; Wrutniak, C.; Cabello, G. BTG1: A triiodothyronine target involved in the myogenic influence of the hormone. Exp. Cell Res 1999, 249, 337–348. [Google Scholar]
- Cho, I.J.; Lee, A.K.; Lee, S.J.; Lee, M.G.; Kim, S.G. Repression by oxidative stress of iNOS and cytokine gene induction in macrophages results from AP-1 and NF-kappaB inhibition mediated by B cell translocation gene-1 activation. Free Radic. Biol. Med 2005, 39, 1523–1536. [Google Scholar]
- Li, F.; Liu, J.; Park, E.S.; Jo, M.; Curry, T.E., Jr. The B cell translocation gene (BTG) family in the rat ovary: Hormonal induction, regulation, and impact on cell cycle kinetics. Endocrinology 2009, 150, 3894–3902. [Google Scholar]
- Urzua, U.; Roby, K.F.; Gangi, L.M.; Cherry, J.M.; Powell, J.I.; Munroe, D.J. Transcriptomic analysis of an in vitro murine model of ovarian carcinoma: Functional similarity to the human disease and identification of prospective tumoral markers and targets. J. Cell. Physiol 2006, 206, 594–602. [Google Scholar]
- Waanders, E.; Scheijen, B.; van der Meer, L.T.; van Reijmersdal, S.V.; van Emst, L.; Kroeze, Y.; Sonneveld, E.; Hoogerbrugge, P.M.; van Kessel, A.G.; van Leeuwen, F.N.; et al. The origin and nature of tightly clustered BTG1 deletions in precursor B-cell acute lymphoblastic leukemia support a model of multiclonal evolution. PLoS Genet 2012, 8, e1002533. [Google Scholar]
- Deng, B.; Zhao, Y.; Gou, W.; Chen, S.; Mao, X.; Takano, Y.; Zheng, H. Decreased expression of BTG3 was linked to carcinogenesis, aggressiveness, and prognosis of ovarian carcinoma. Tumour Biol 2013. [Google Scholar] [CrossRef]
- Chen, Q.; Watson, J.T.; Marengo, S.R.; Decker, K.S.; Coleman, I.; Nelson, P.S.; Sikes, R.A. Gene expression in the LNCaP human prostate cancer progression model: Progression associated expression in vitro corresponds to expression changes associated with prostate cancer progression in vivo. Cancer Lett. 2006, 244, 274–288. [Google Scholar]
- Wang, Y.; Shao, C.; Shi, C.H.; Zhang, L.; Yue, H.H.; Wang, P.F.; Yang, B.; Zhang, Y.T.; Liu, F.; Qin, W.J.; et al. Change of the cell cycle after flutamide treatment in prostate cancer cells and its molecular mechanism. Asian J. Androl 2005, 7, 375–380. [Google Scholar]
- Zheng, H.; Takahashi, H.; Murai, Y.; Cui, Z.; Nomoto, K.; Niwa, H.; Tsuneyama, K.; Takano, Y. Expressions of MMP-2, MMP-9 and VEGF are closely linked to growth, invasion, metastasis and angiogenesis of gastric carcinoma. Anticancer Res 2006, 26, 3579–3583. [Google Scholar]
- Ghayad, S.E.; Cohen, P.A. Inhibitors of the PI3K/Akt/mTOR pathway: New hope for breast cancer patients. Recent Pat. Anticancer Drug Discov 2010, 5, 29–57. [Google Scholar]
- Kunze, D.; Kraemer, K.; Erdmann, K.; Froehner, M.; Wirth, M.P.; Fuessel, S. Simultaneous siRNA- mediated knockdown of antiapoptotic BCL2, Bcl-xL, XIAP and survivin in bladder cancer cells. Int. J. Oncol. 2012, 41, 1271–1277. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, Y.; Gou, W.-F.; Chen, S.; Takano, Y.; Xiu, Y.-L.; Zheng, H.-C. BTG1 Expression Correlates with the Pathogenesis and Progression of Ovarian Carcinomas. Int. J. Mol. Sci. 2013, 14, 19670-19680. https://doi.org/10.3390/ijms141019670
Zhao Y, Gou W-F, Chen S, Takano Y, Xiu Y-L, Zheng H-C. BTG1 Expression Correlates with the Pathogenesis and Progression of Ovarian Carcinomas. International Journal of Molecular Sciences. 2013; 14(10):19670-19680. https://doi.org/10.3390/ijms141019670
Chicago/Turabian StyleZhao, Yang, Wen-Feng Gou, Shuo Chen, Yasuo Takano, Yin-Ling Xiu, and Hua-Chuan Zheng. 2013. "BTG1 Expression Correlates with the Pathogenesis and Progression of Ovarian Carcinomas" International Journal of Molecular Sciences 14, no. 10: 19670-19680. https://doi.org/10.3390/ijms141019670