Antinociceptive Action of Isolated Mitragynine from Mitragyna Speciosa through Activation of Opioid Receptor System

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
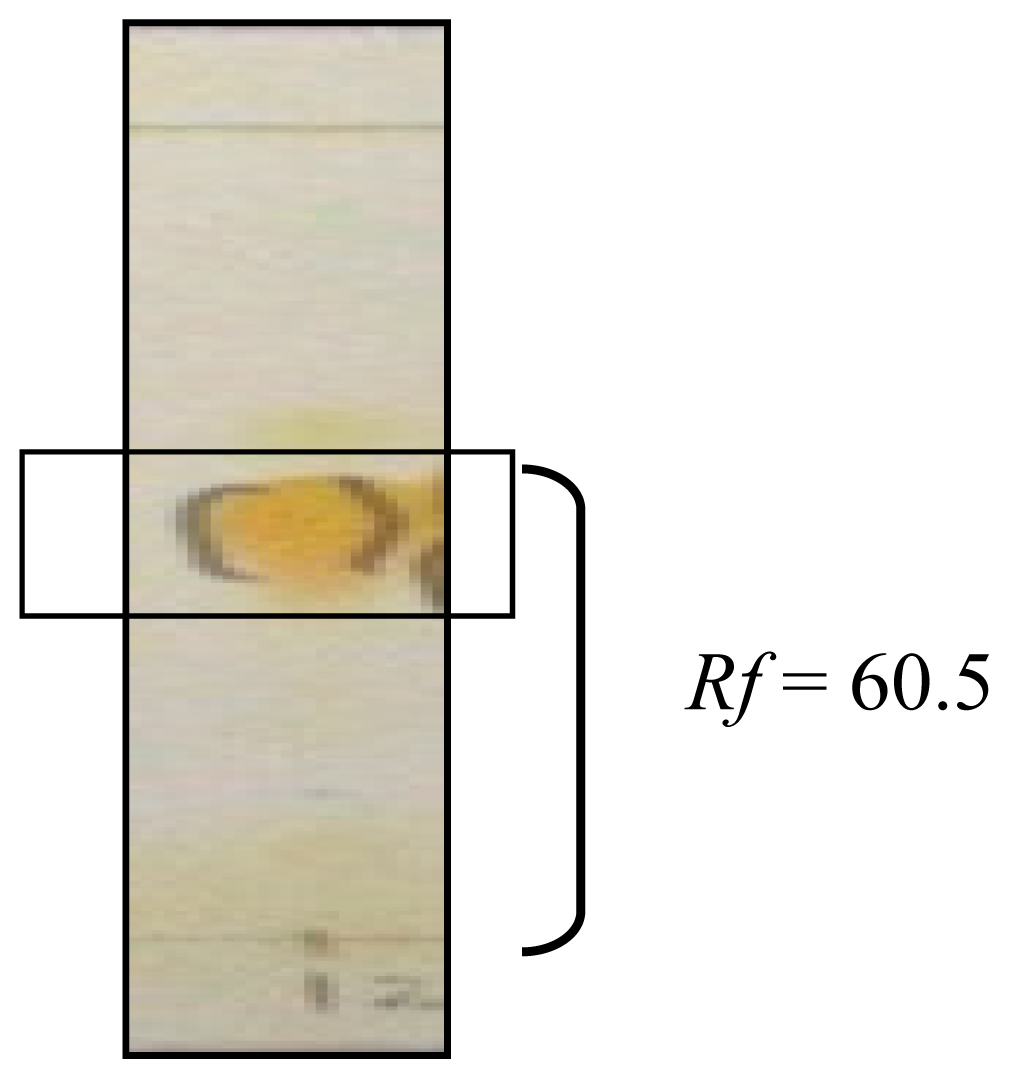
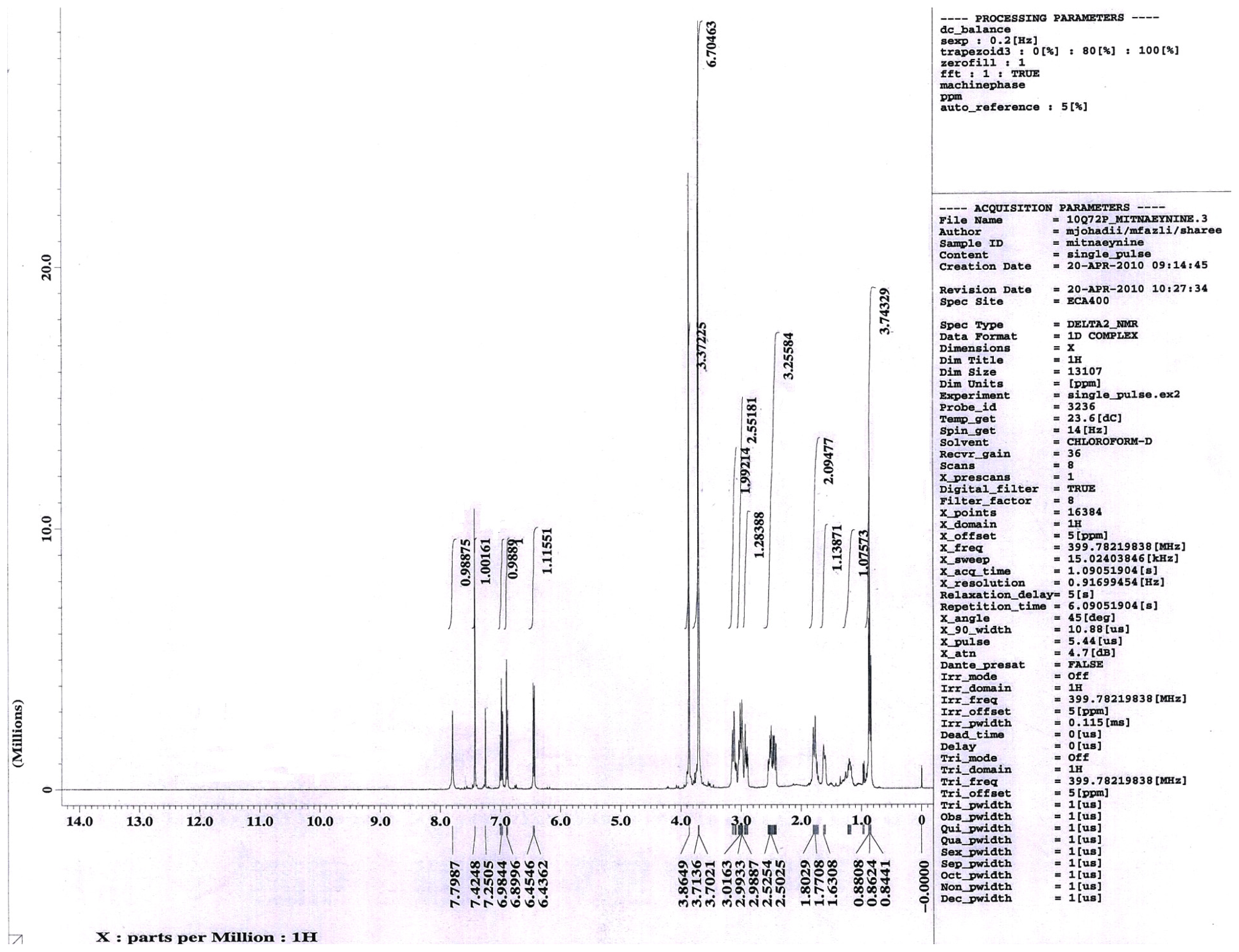
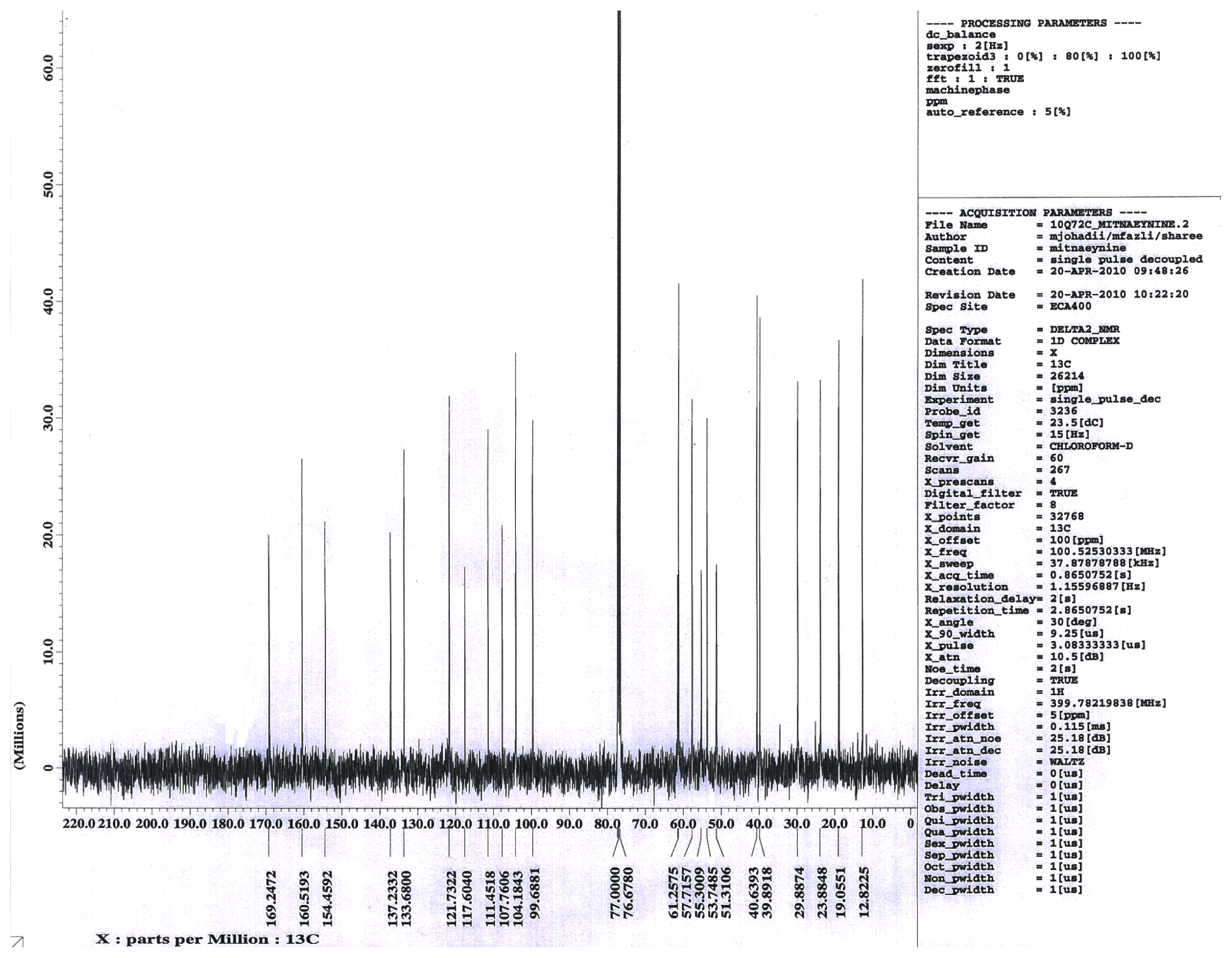
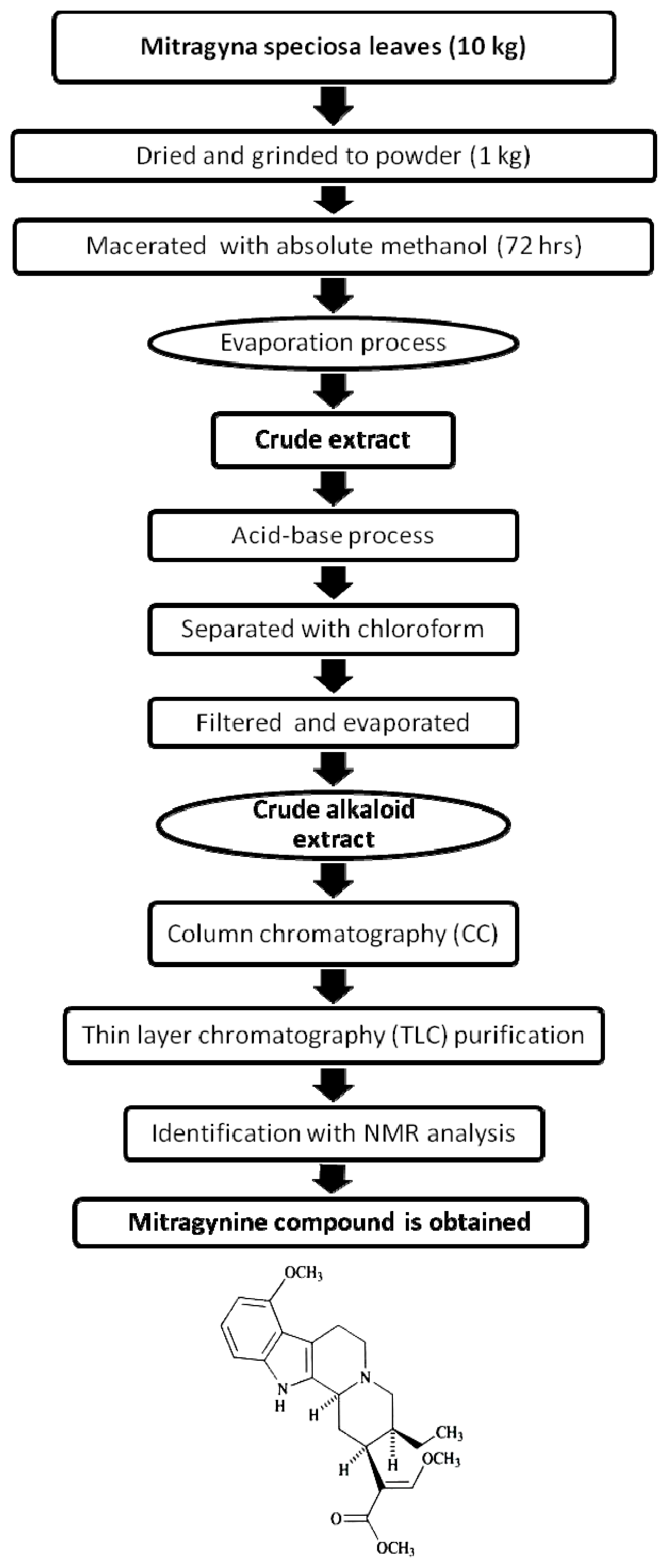
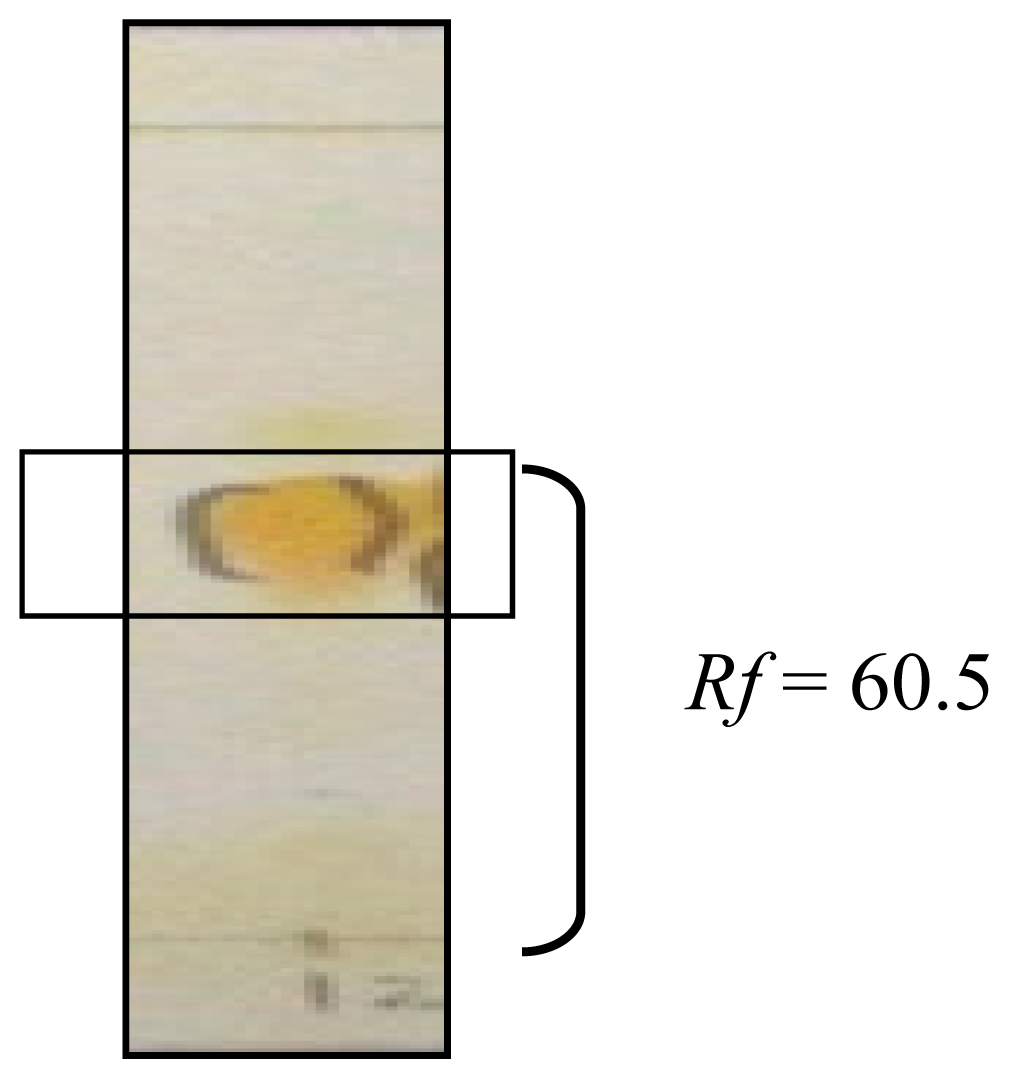
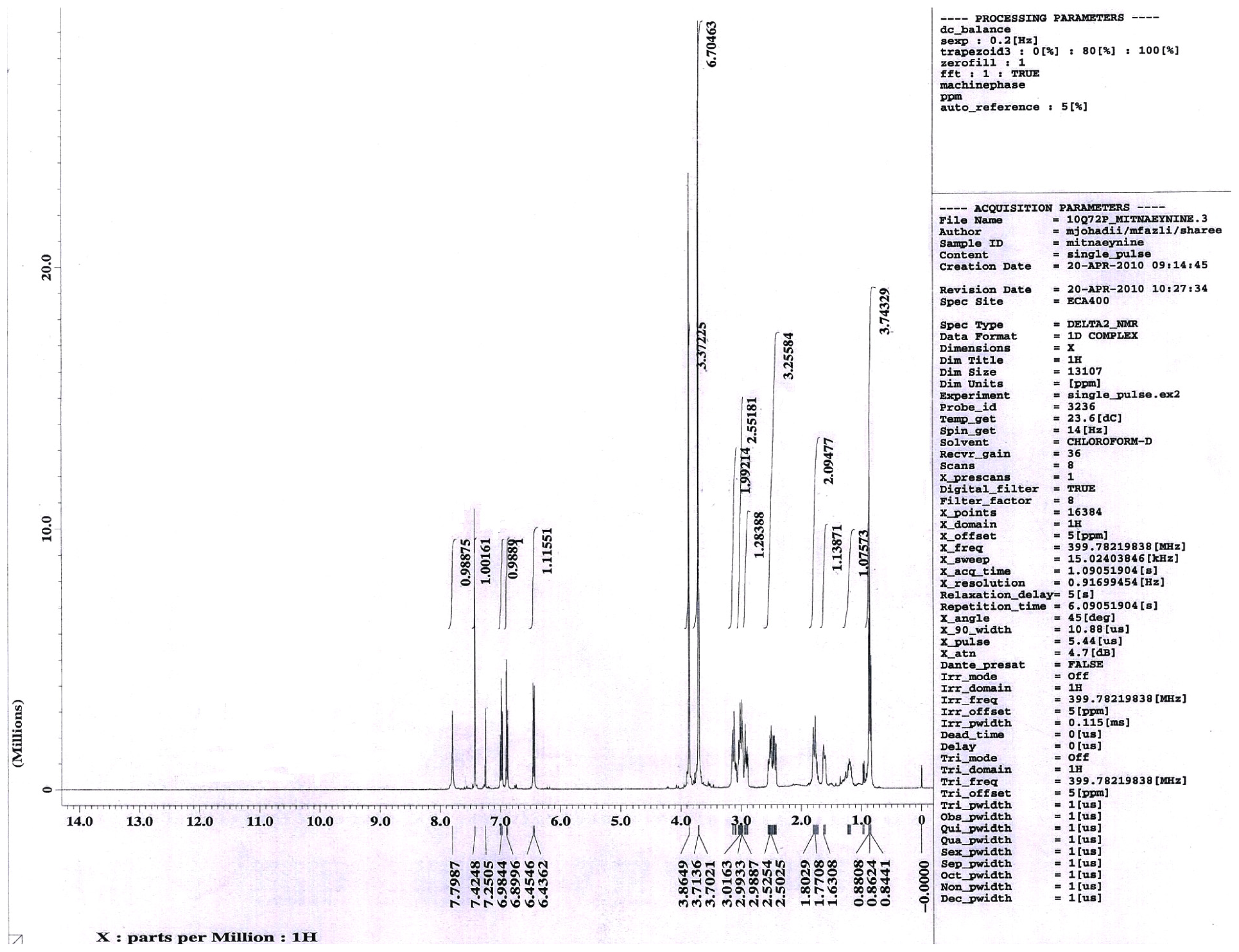
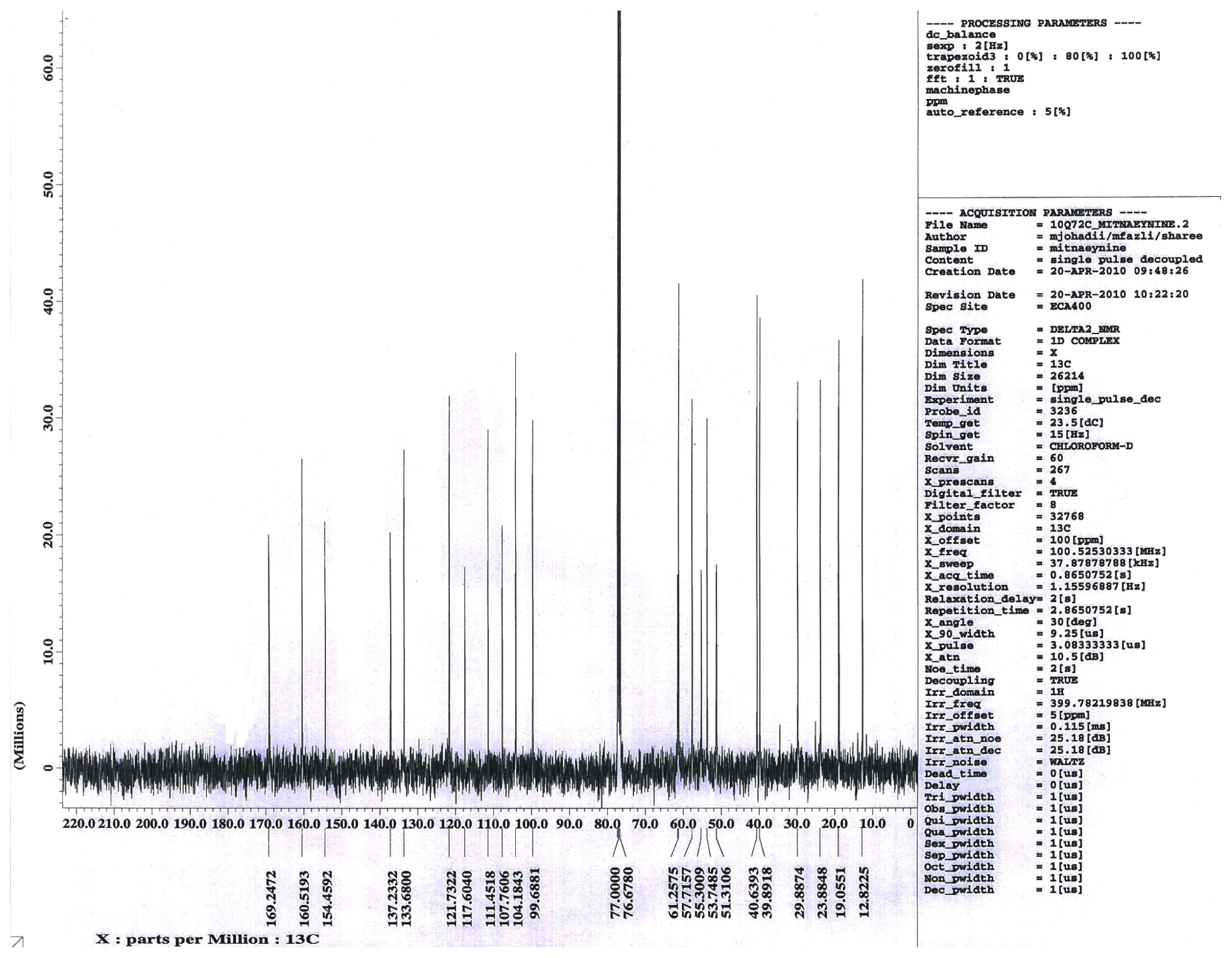
2.1.1. Isolation of Mitragynine from Mitragyna Speciosa Leaves
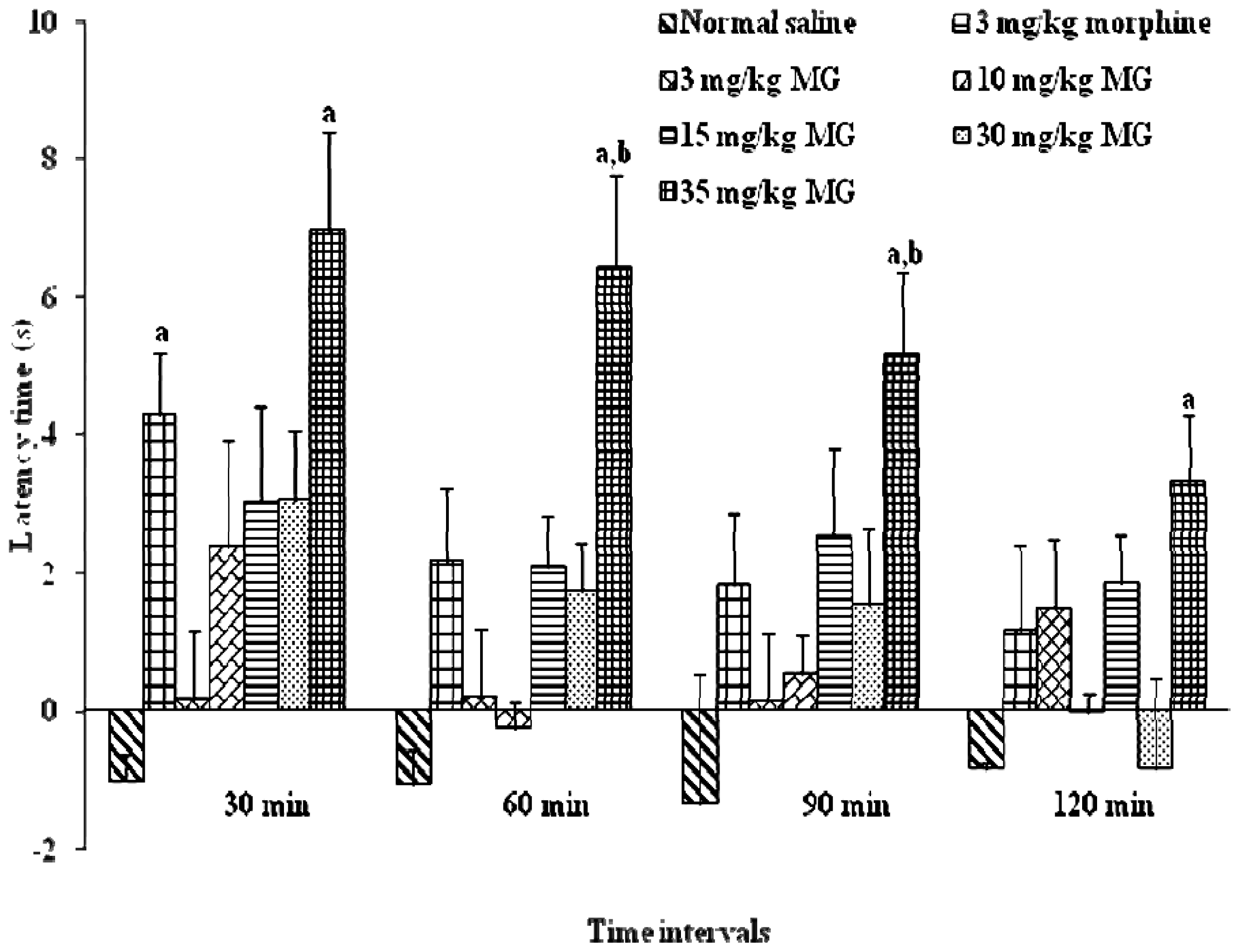
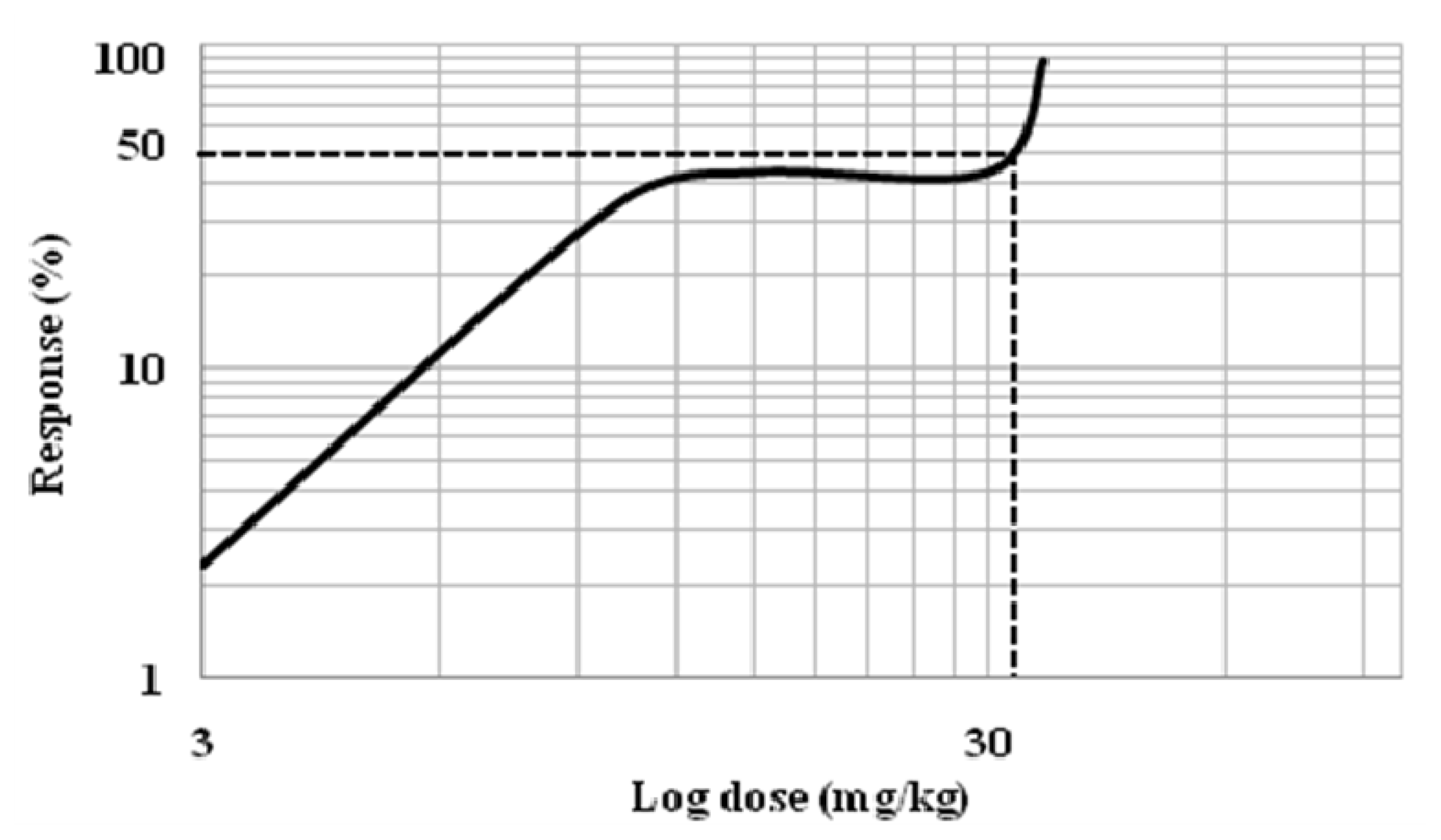
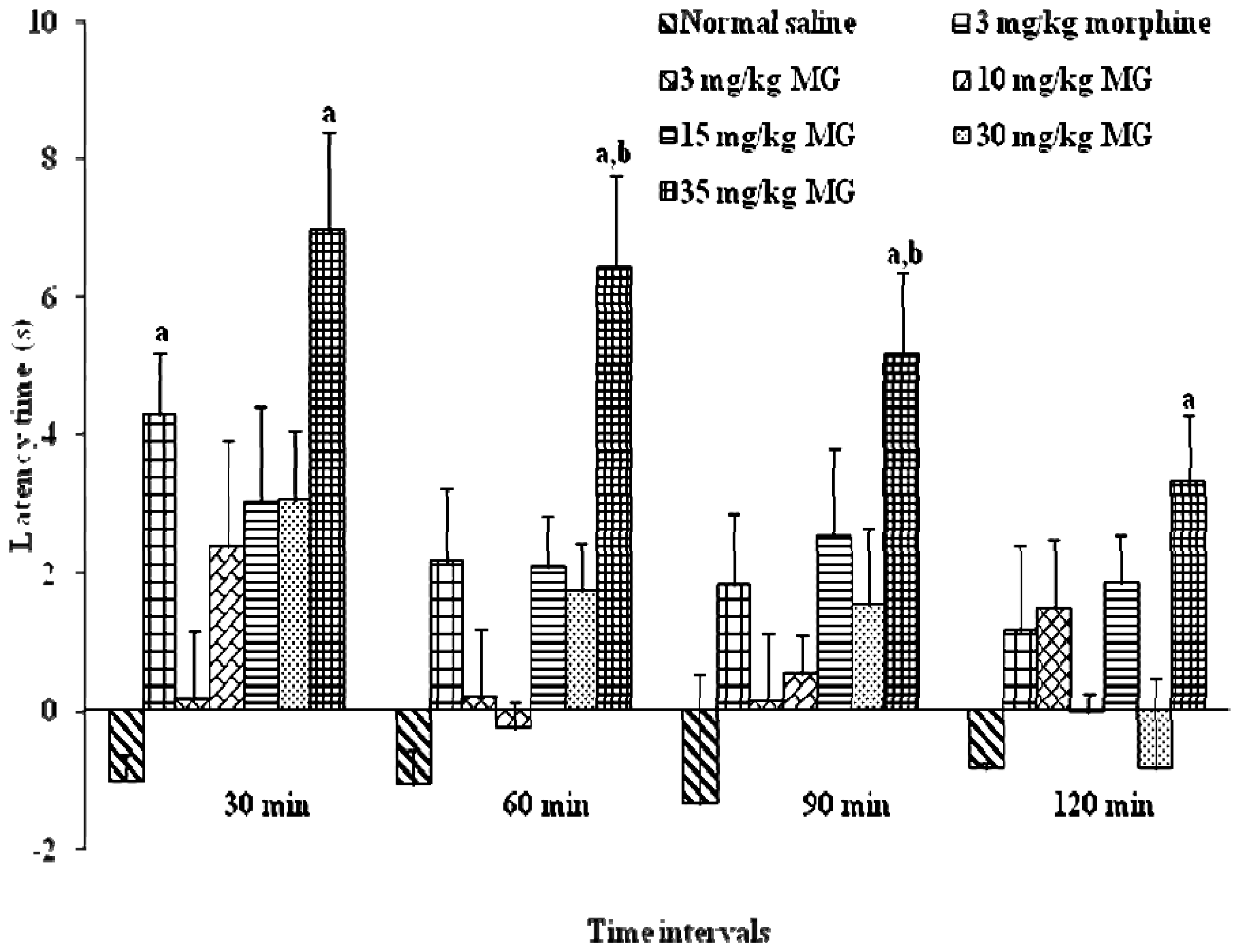
2.1.2. Effects of Mitragynine on Hot-Plate Test (HPT) and the Determination of Effective Dose 50 (ED50)
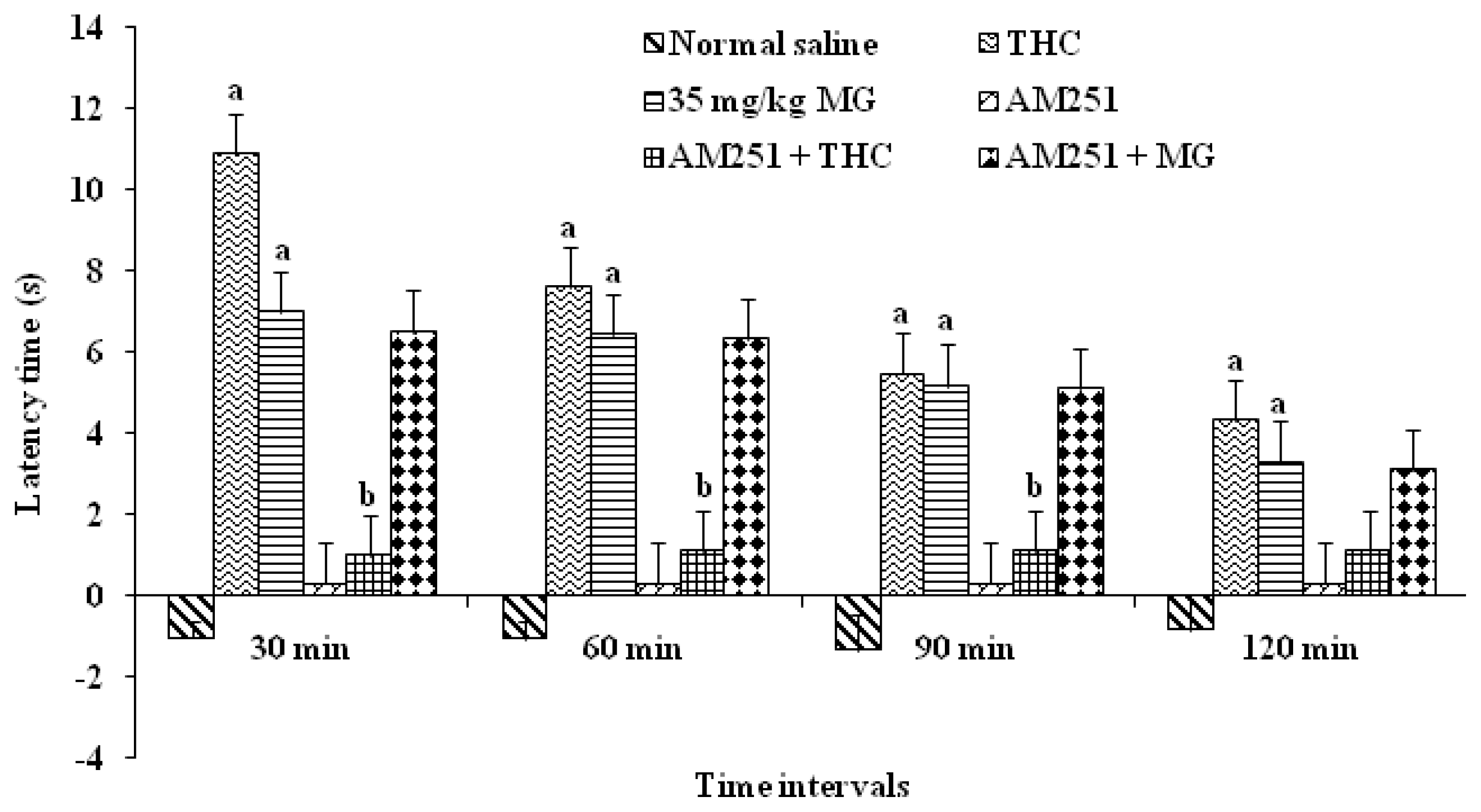
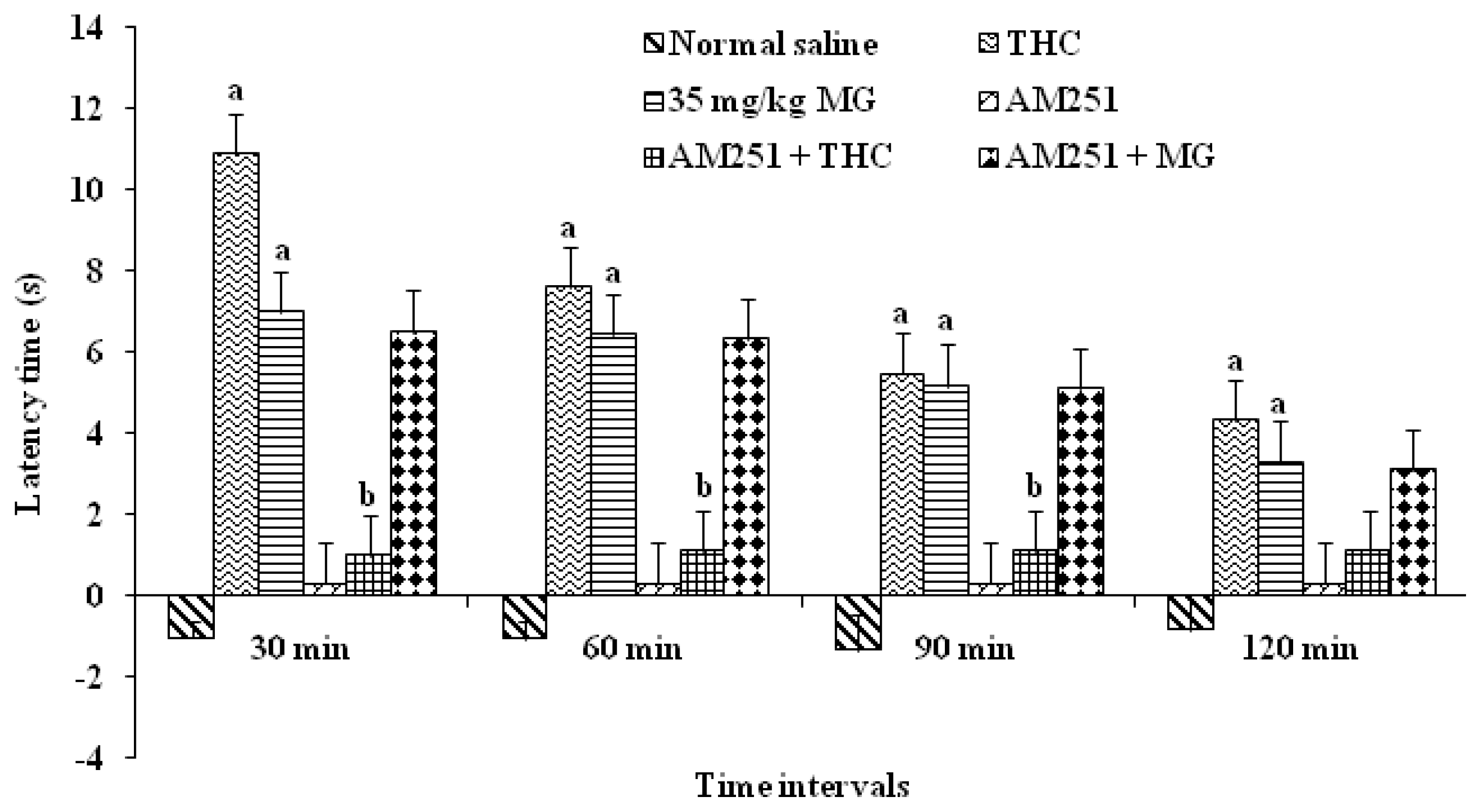
2.1.3. The determination of the Effect of Mitragynine Following the Administration of Antagonists
2.1.3.1. The Determination of the Effect of Mitragynine Following the Administration of CB1 Antagonists
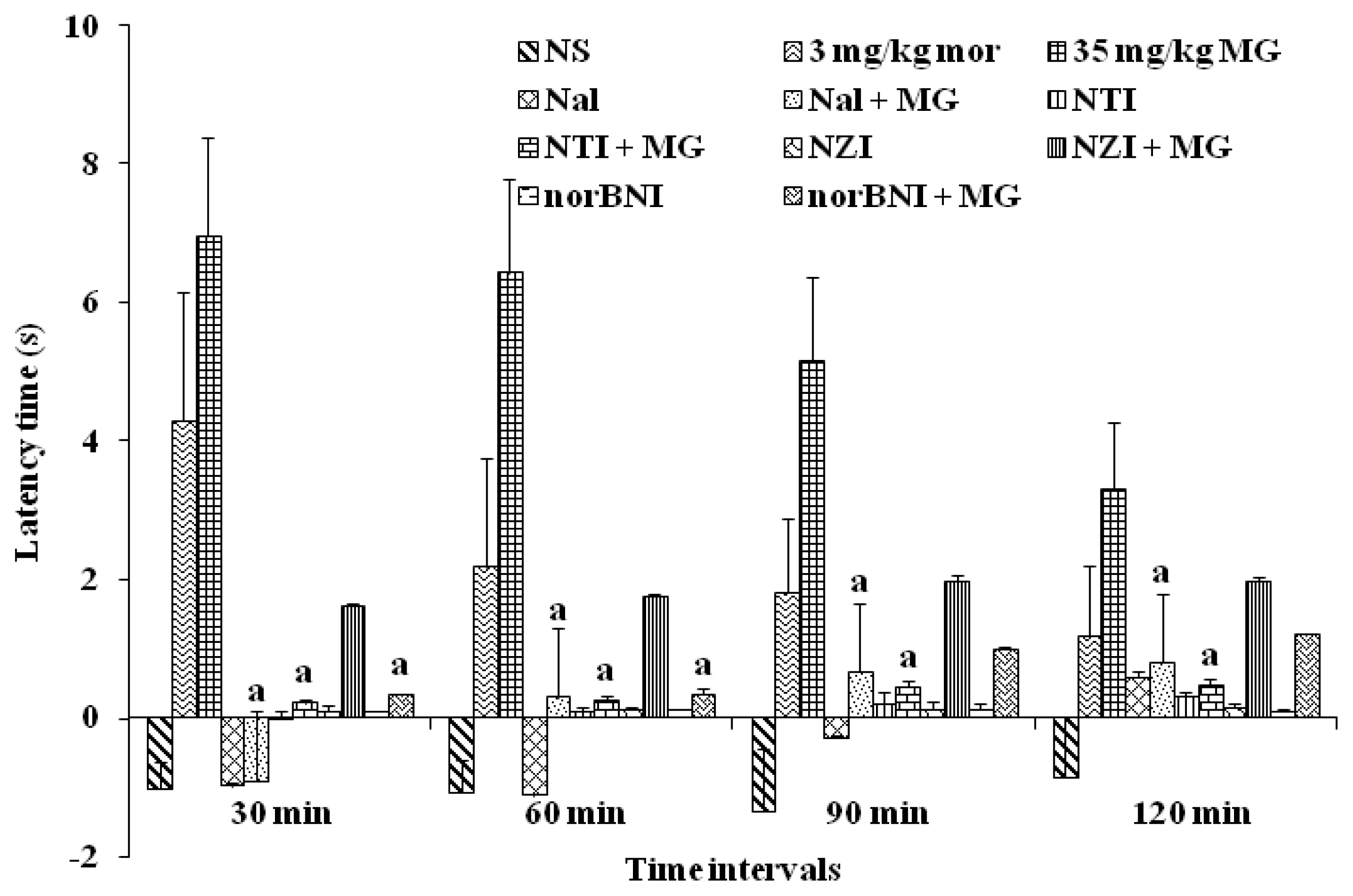
2.1.3.2. The Determination of the Effect of Mitragynine Following the Administration of Opioid Receptor Antagonists
2.2. Discussion
3. Experimental Section
3.1. Isolation of Mitragynine from Mitragyna Speciosa Leaves
3.2. Animals
3.3. Drugs
3.4. Antinociceptive Study and the Determination of Effective Dose 50 (ED50)
3.5. The Determination of the Effect of Mitragynine Following the Administration of Cannabinoid and Opioid Antagonists
3.6. Hot-Plate test
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Howlett, A.C.; Barth, F.; Bonner, T.I.; Cabral, G.; Casellas, P.; Devane, W.A.; Felder, C.C.; Herkenham, M.; Mackie, K.; Martin, B.R.; et al. International Union of Pharmacology. XXVII. Classification of cannabinoid receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar]
- Martin, B.R.; Compton, D.R.; Thomas, B.F.; Prescott, W.R.; Little, P.J.; Razdan, R.K.; Johnson, M.R.; Melvin, L.S.; Mechoulam, R.; Ward, S.J. Behavioral, biochemical, and molecular modeling evaluations of cannabinoid analogs. Pharmacol. Biochem. Behav 1991, 40, 471–478. [Google Scholar]
- Herkenham, H. Localization of cannabinoid receptors in brain and periphery. In Cannabinoid receptors; Pertwee, R.G., Ed.; Academic Press: London, UK, 1995; pp. 145–166. [Google Scholar]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar]
- Krogsgaard-Larsen, P. Textbook of Drug Design and Discovery, 5th ed; Tayor and Francis; New York, NY, USA, 2010; pp. 215–240. [Google Scholar]
- Li, J.; Daughters, R.S.; Bullis, C.; Bengiamin, R.; Stucky, M.W.; Brennan, J.; Simone, D.A. The cannabinoid receptor agonist WIN 55,212-2 mesylate blocks the development of hyperalgesia produced by capsaicin in rats. Pain 1999, 81, 25–33. [Google Scholar]
- Herzberg, U.; Eliav, E.; Bennett, G.J.; Kopin, I.J. The analgesic effects of R(+)-WIN 55,212-2 mesylate, a high affinity cannabinoid agonist, in a rat model of neuropathic pain. Neurosci. Lett 1997, 221, 157–160. [Google Scholar]
- Walker, J.M.; Hohmann, A.G.; Martin, W.J.; Strangman, N.M.; Huang, S.M.; Tsou, K. The neurobiology of cannabinoid analgesia. Life Sci 1999, 65, 665–673. [Google Scholar]
- Cichewicz, D.L. Synergistic interactions between cannabinoid and opioid analgesics. Life Sci 2004, 74, 1317–1324. [Google Scholar]
- Smith, P.B.; Welch, S.P.; Martin, B.R. Interactions between delta 9-tetrahydrocannabinol and kappa opioids in mice. J. Pharma. Exp. Ther 1994, 268, 1381–1387. [Google Scholar]
- Welch, S.P.; Eads, M. Synergistic interactions of endogenous opioids and cannabinoid systems. Brain Res 1999, 848, 183–190. [Google Scholar]
- DuPen, A.; Shen, D.; Ersek, M. Mechanisms of opioid-induced tolerance and hyperalgesia. Pain Manag. Nurs 2007, 8, 113–121. [Google Scholar]
- Lynn, W.; Crespo, L.M.; George, D.; Carl, F.G.; Stephanie, W. Brody’s Human Pharmacology Molecular to Clinical; Mosby Elsevier: Pennsylvania, PA, USA, 2010; pp. 74–78. [Google Scholar]
- Jansen, K.L.; Prast, C.J. Ethnopharmacology of kratom and the Mitragyna alkaloids. J. Ethnopharmacol 1988, 23, 115–119. [Google Scholar]
- Shaik Mossadeq, W.M.; Sulaiman, M.R.; Tengku Mohamad, T.A.; Chiong, H.S.; Zakaria, Z.A.; Jabit, M.L.; Baharuldin, M.T.; Israf, D.A. Anti-inflammatory and antinociceptive effects of Mitragyna speciosa Korth methanolic extract. Med. Princ. Pract 2009, 18, 378–384. [Google Scholar]
- Takayama, H. Chemistry and pharmacology of analgesic indole alkaloids from the rubiaceous plant. Mitragyna speciosa. Chem. Pharmceut. Bull. (Tokyo) 2004, 52, 916–928. [Google Scholar]
- Takayama, H.; Ishikawa, H.; Kurihara, M.; Kitajima, M.; Aimi Ponglux, N.; Koyama, F.; Matsumoto, K.; Moriyama, T.; Yamamoto, L.T.; Watanabe, K.; et al. Studies on the synthesis and opioid agonistic activities of mitragynine-related indole alkaloids: Discovery of opioid agonists structurally different from other opioid ligands. J. Med. Chem. 2002, 45, 1949–1956. [Google Scholar]
- Matsumoto, K.; Takayama, H.; Ishikawa, H.; Aimi, N.; Ponglux, D.; Watanabe, K.; Horie, S. Partial agonistic effect of 9-hydroxycorynantheidine on mu-opioid receptor in the guinea-pig ileum. Life Sci 2006, 78, 2265–2271. [Google Scholar]
- Boyer, E.W.; Babu, K.M.; Adkins, J.E.; McCurdy, C.R.; Halpern, J.H. Self-treatment of opioid withdrawal using kratom (Mitragynia speciosa Korth). Addiction 2008, 103, 1048–1050. [Google Scholar]
- Vicknasingam, B.; Narayanan, S.; Beng, G.T.; Mansor, S.M. The informal use of ketum (Mitragyna speciosa) for opoid withdrawal in the northern states of peninsular Malaysia and implications for drug substitution therapy. Int. J. Drug Pol 2010, 21, 283–288. [Google Scholar]
- Watanabe, K.; Yano, S.; Horie, S.; Yamamoto, L.T. Inhibitory effect of mitragynine, an alkaloid with analgesic effect from Thai medicinal plant Mitragyna speciosa, on electrically stimulated contraction of isolated guinea-pig ileum through the opioid receptor. Life Sci 1997, 60, 933–942. [Google Scholar]
- Gurdeep, R.; Chatwal, S.; Anand, K. Instrumental methods of chemical analysis; Himalaya Publishing House: Maharashtra, India, 1998; pp. 2.185–2.234. [Google Scholar]
- Chittrakarn, S.; Keawpradub, N.; Sawangjaroen, K.; Kansenalak, S.; Janchawee, B. The neuromuscular blockade produced by pure alkaloid, mitragynine and methanol extract of kratom leaves (Mitragyna speciosa Korth.). J. Ethnopharmacol 2010, 129, 344–349. [Google Scholar]
- Kumarnsit, E.; Keawpradub, N.; Nuankaew, W. Effect of Mitragyna speciosa aqueous extract on ethanol withdrawal symptoms in mice. Fitoterapia 2007, 78, 182–185. [Google Scholar]
- Kumarnsit, E.; Vongvatcharanon, U.; Keawpradub, N.; Intasaro, P. Fos-like immunoreactivity in rat dorsal raphe nuclei induced by alkaloid extract of Mitragyna speciosa. Neurosci. Lett 2007, 416, 128–132. [Google Scholar]
- Houghton, P.J.; Ikram, M.S. 3-Dehydromitragynine: an alkaloid from Mitragyna speciosa. Phytochemistry 1986, 25, 2910–2912. [Google Scholar]
- Yaksh, T.L.; Malmberg, A.B. Spinal actions of NSAIDS in blocking spinally mediated hyperalgesia: The role of cyclooxygenase products. Agents Actions Suppl 1993, 41, 89–100. [Google Scholar]
- Matsumoto, K.; Mizowaki, M.; Suchitra, T.; Murakami, Y.; Takayama, H.; Sakai, S.; Aimi, N.; Watanabe, H. Central antinociceptive effects of mitragynine in mice: contribution of descending noradrenergic and serotonergic systems. Eur. J. Pharmacol 1996, 317, 75–81. [Google Scholar]
- Matsumoto, K.; Mizowaki, M.; Suchitra, T.; Takayama, H.; Sakai, S.; Aimi, N.; Watanabe, H. Antinociceptive action of mitragynine in mice: Evidence for the involvement of supraspinal opioid receptors. Life Sci 1996, 59, 1149–1155. [Google Scholar]
- Ikram, M. Studies on the components of fresh leaves of Mitragyna speciosa. Proceedings of 2nd Meeting of the Natural Products Research Group, (NPRG’86), Chemistry Department, Universiti Kebangsaan Malaysia, Selangor, Malaysia; 1986; pp. 123–127. [Google Scholar]
- Gaoni, Y.; Mechoulam, R. Isolation, structure and partial synthesis of an active constituent of hashish. J. Am. Chem. Soc 1964, 86, 1646–1647. [Google Scholar]
- Rios, C.; Gomes, I.; Devi, L.A. Mu opioid and CB1 cannabinoid receptor interactions: reciprocal inhibition of receptor signaling and neuritogenesis. Br. J. Pharmacol 2006, 148, 387–395. [Google Scholar]
- Macko, E.; Weisbach, J.A.; Douglas, B. Some observations on the pharmacology of mitragynine. Arch. Int. Pharmacodyn. Ther 1972, 198, 145–161. [Google Scholar]
- Thongpradichote, S.; Matsumoto, K.; Tohda, M.; Takayama, H.; Aimi, N.; Sakai, S.; Watanabe, H. Identification of opioid receptor subtypes in antinociceptive actions of supraspinally-administered mitragynine in mice. Life Sci 1998, 62, 1371–1378. [Google Scholar]
- Ponglux, D.; Wongseripipatana, S.; Takayama, H.; Kikuchi, M.; Kurihara, M.; Kitajima, M.; Aimi, N.; Sakai, S. A New Indole Alkaloid, 7 alpha-Hydroxy-7H-mitragynine, from Mitragyna speciosa in Thailand. Planta Med 1994, 60, 580–581. [Google Scholar]
- Reanmongkol, W.; Keawpradub, N.; Sawangjaroen, K. Effects of the extracts from Mitragyna speciosa Korth leaves on analgesic and behavioral activities in experimental animals. Songklanakarin J. Sci. Technol 2007, 29, 39–48. [Google Scholar]


© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shamima, A.R.; Fakurazi, S.; Hidayat, M.T.; Hairuszah, I.; Moklas, M.A.M.; Arulselvan, P. Antinociceptive Action of Isolated Mitragynine from Mitragyna Speciosa through Activation of Opioid Receptor System. Int. J. Mol. Sci. 2012, 13, 11427-11442. https://doi.org/10.3390/ijms130911427
Shamima AR, Fakurazi S, Hidayat MT, Hairuszah I, Moklas MAM, Arulselvan P. Antinociceptive Action of Isolated Mitragynine from Mitragyna Speciosa through Activation of Opioid Receptor System. International Journal of Molecular Sciences. 2012; 13(9):11427-11442. https://doi.org/10.3390/ijms130911427
Chicago/Turabian StyleShamima, Abdul Rahman, Sharida Fakurazi, Mohamad Taufik Hidayat, Ithnin Hairuszah, Mohamad Aris Mohd Moklas, and Palanisamy Arulselvan. 2012. "Antinociceptive Action of Isolated Mitragynine from Mitragyna Speciosa through Activation of Opioid Receptor System" International Journal of Molecular Sciences 13, no. 9: 11427-11442. https://doi.org/10.3390/ijms130911427