Antisense Oligonucleotide Against Clusterin Regulates Human Hepatocellular Carcinoma Invasion Through Transcriptional Regulation of Matrix Metalloproteinase-2 and E-Cadherin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
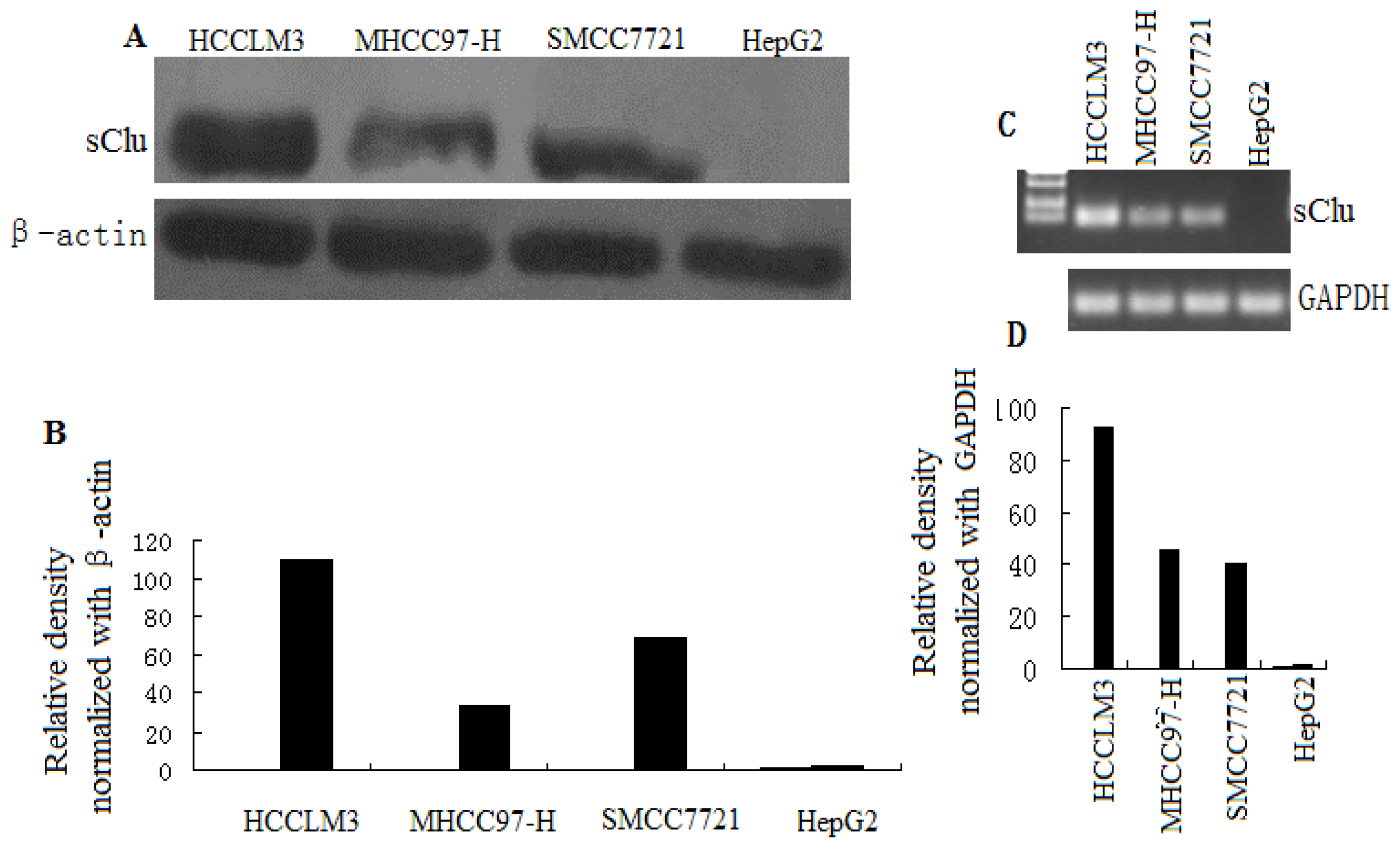
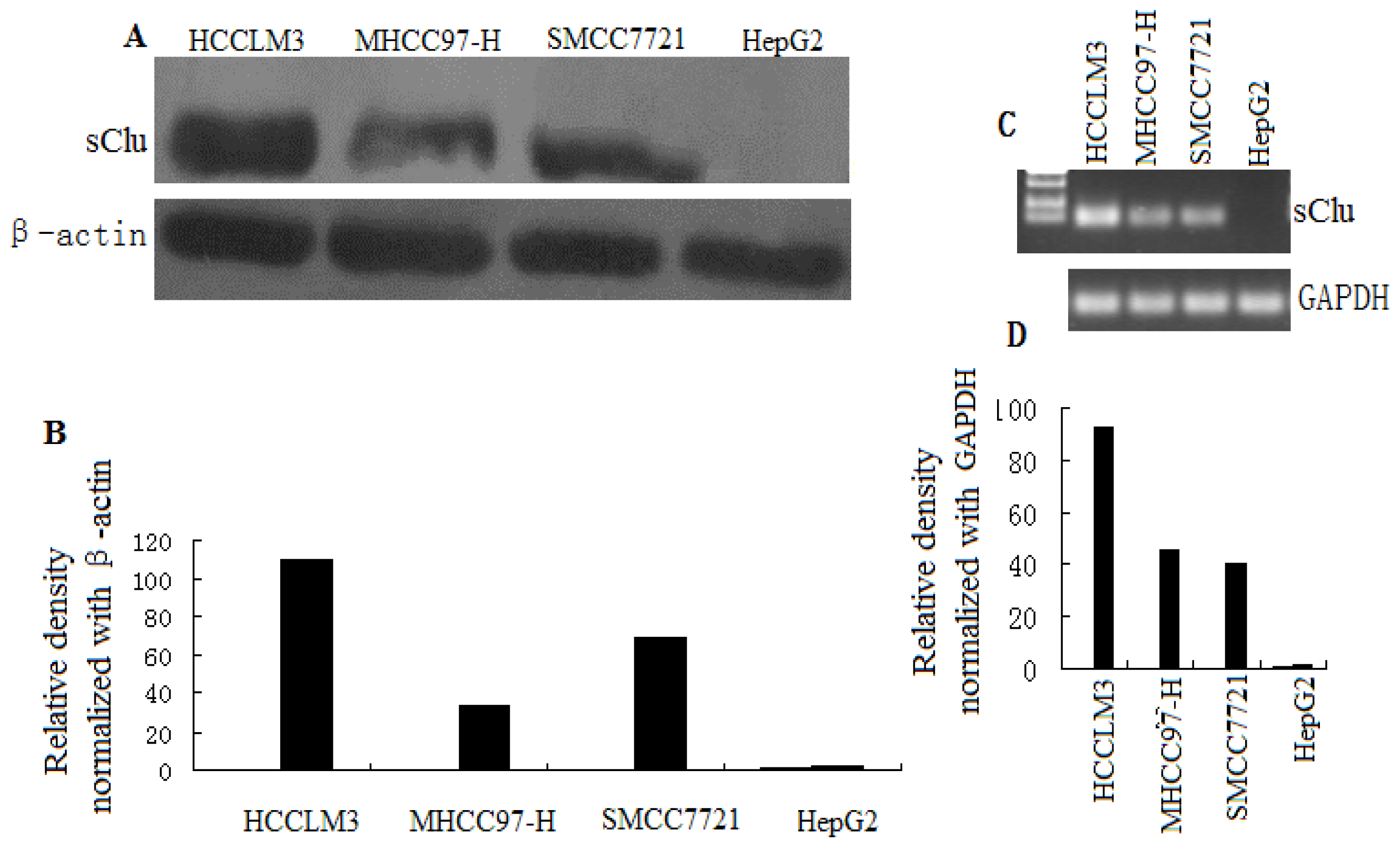
2.1. Clusterin Expression in Human HCC Cells
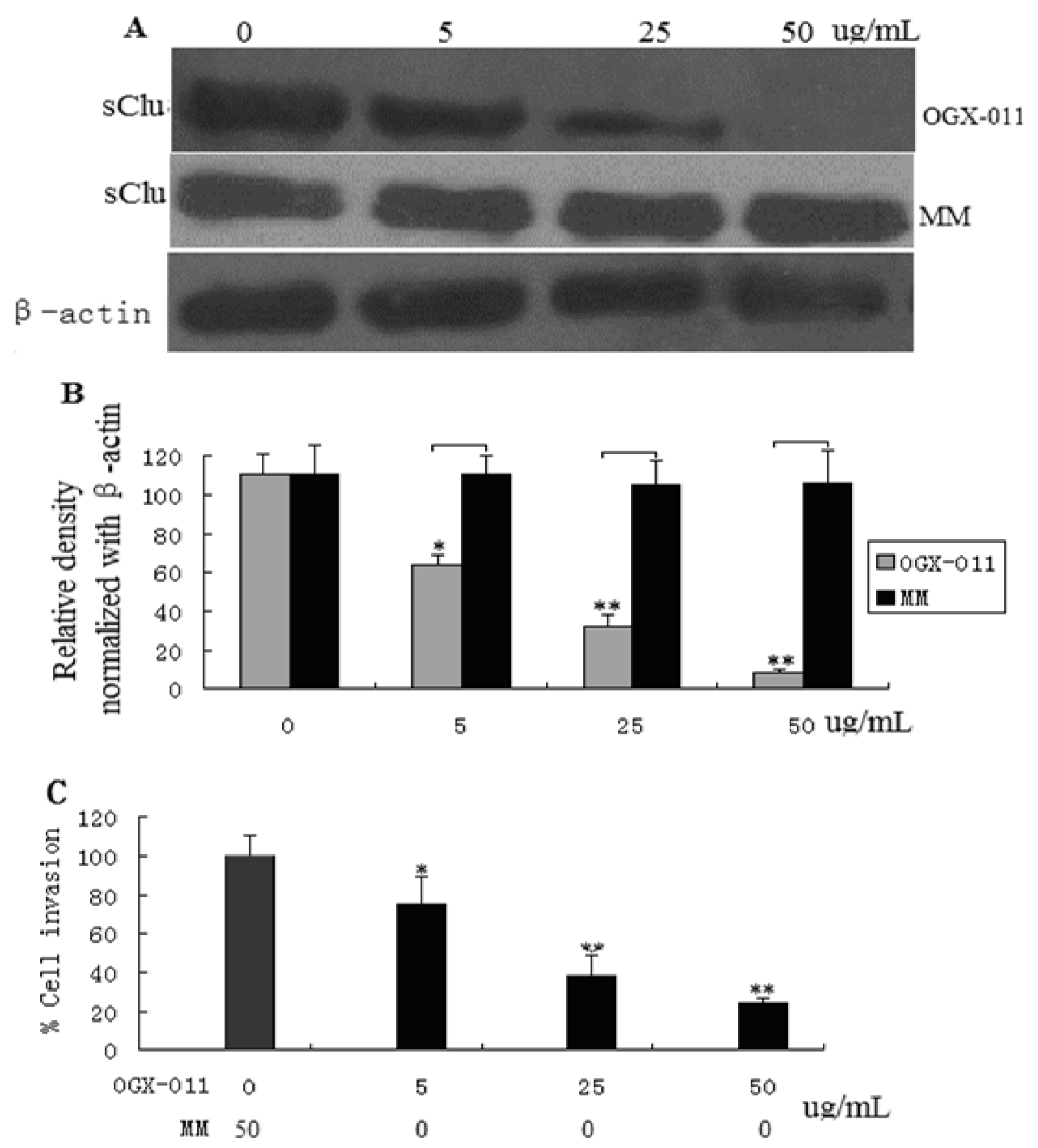
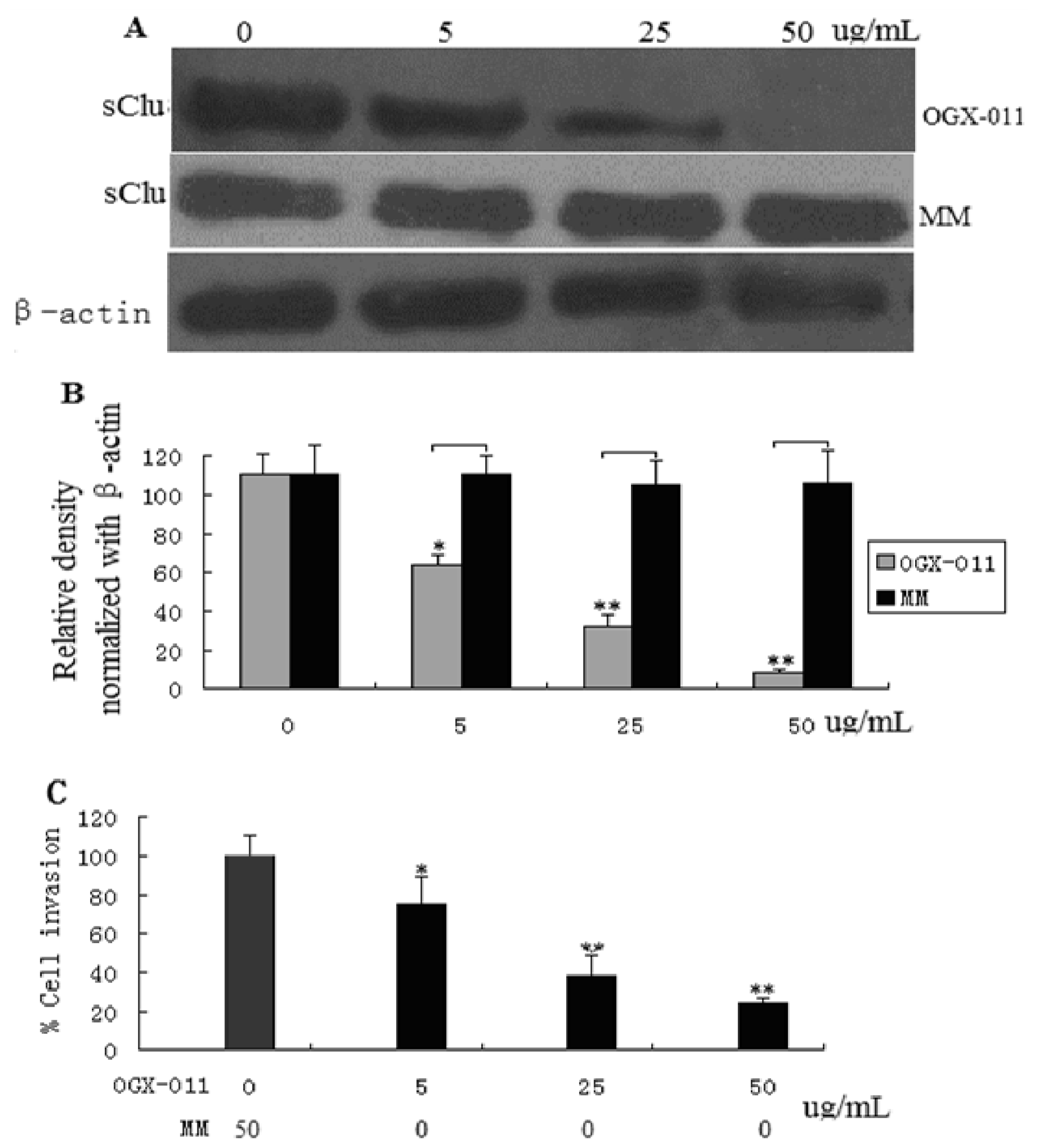
2.2. sCLU Knockdown Decreases Cell Invasion
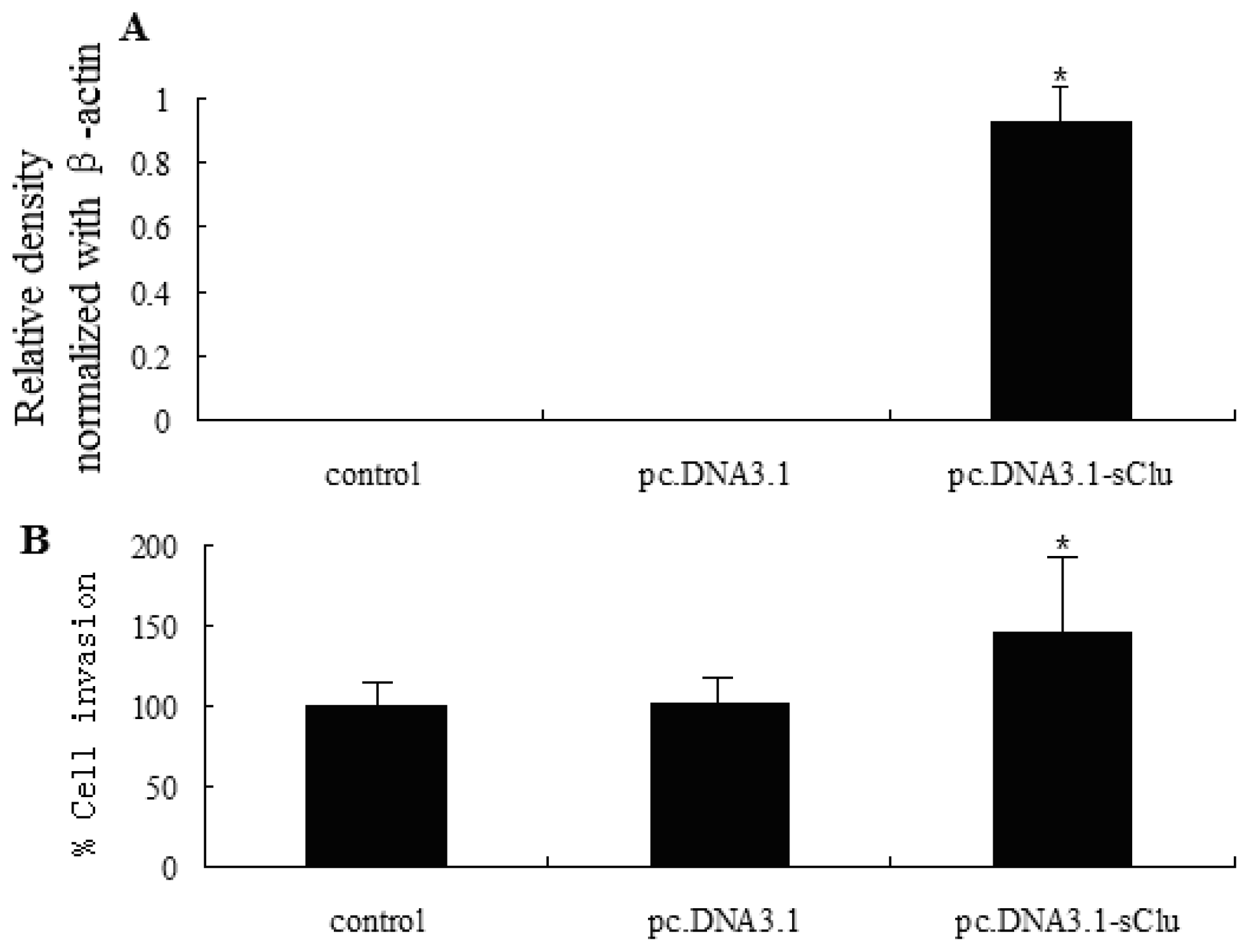
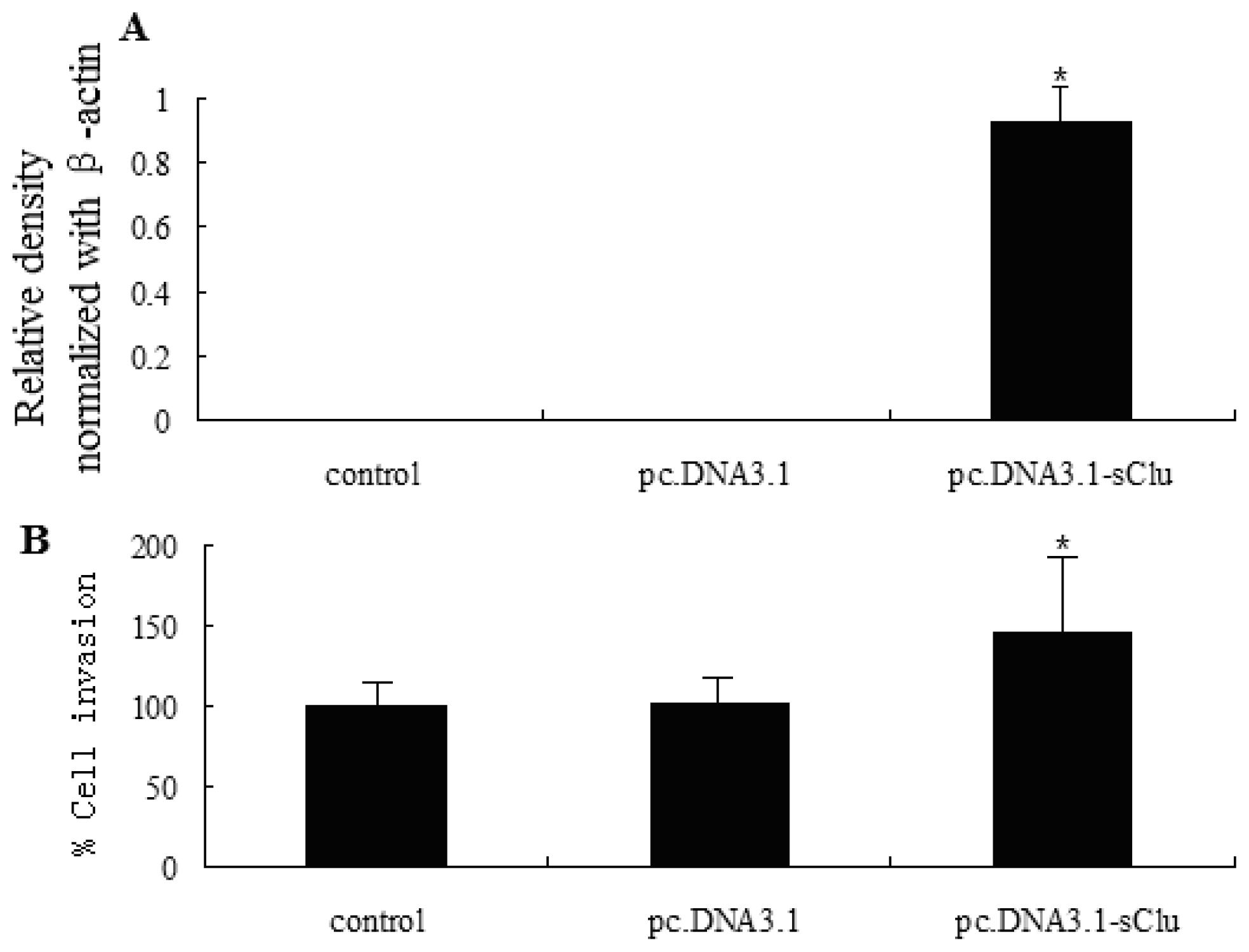
2.3. sCLU Overexpression Increases Cell Invasion
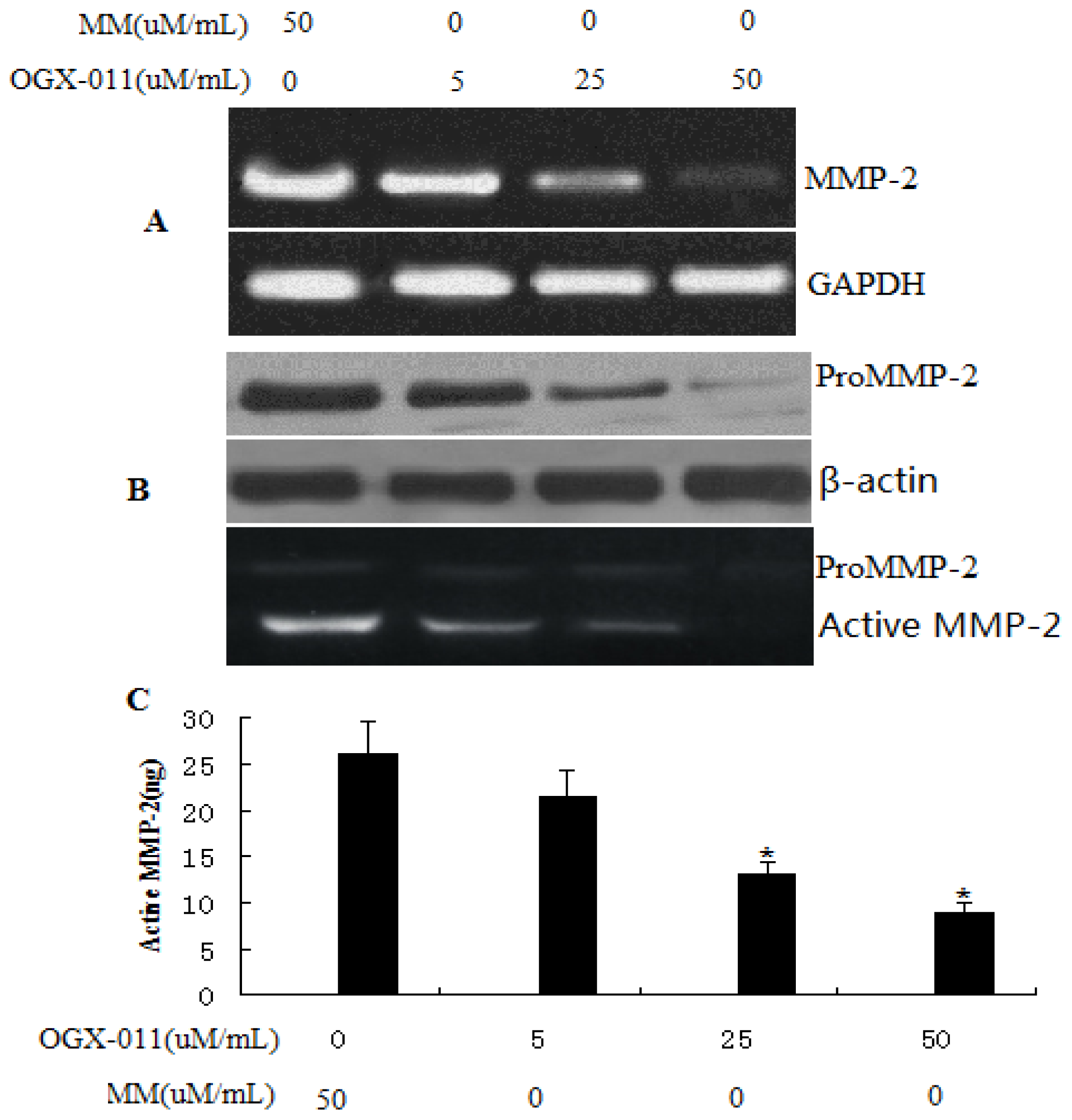
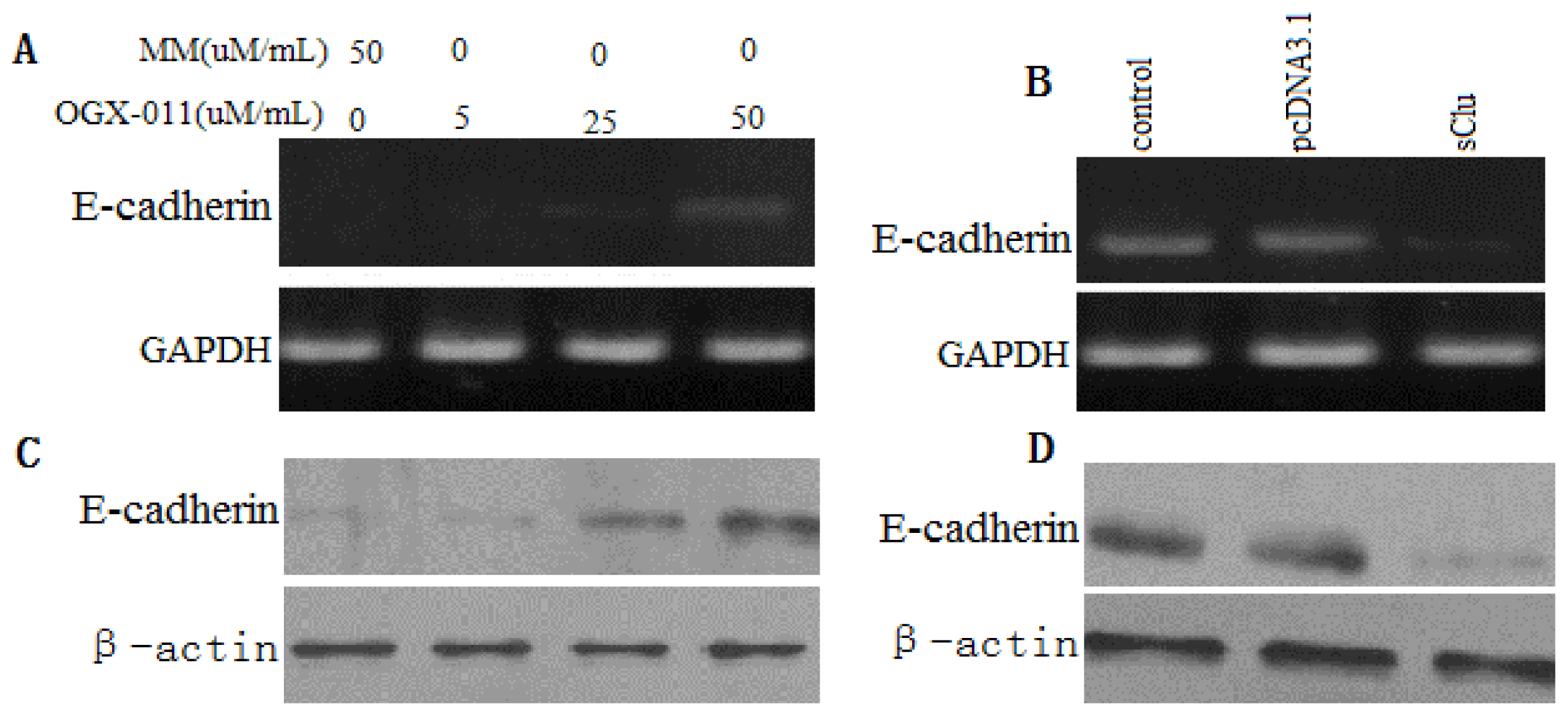
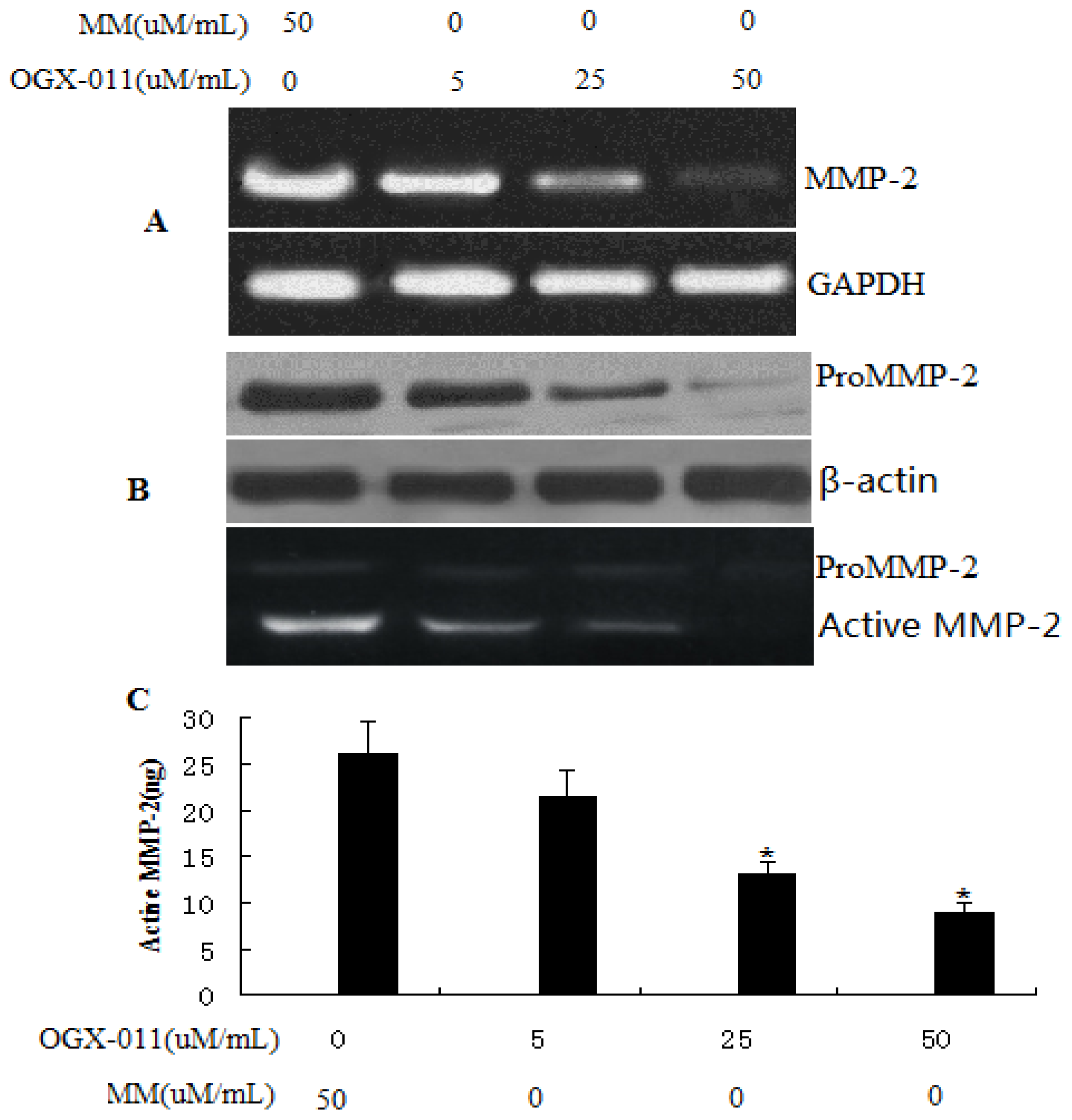
2.4. Effect of sCLU Gene Knockdown on MMP-2 Expression
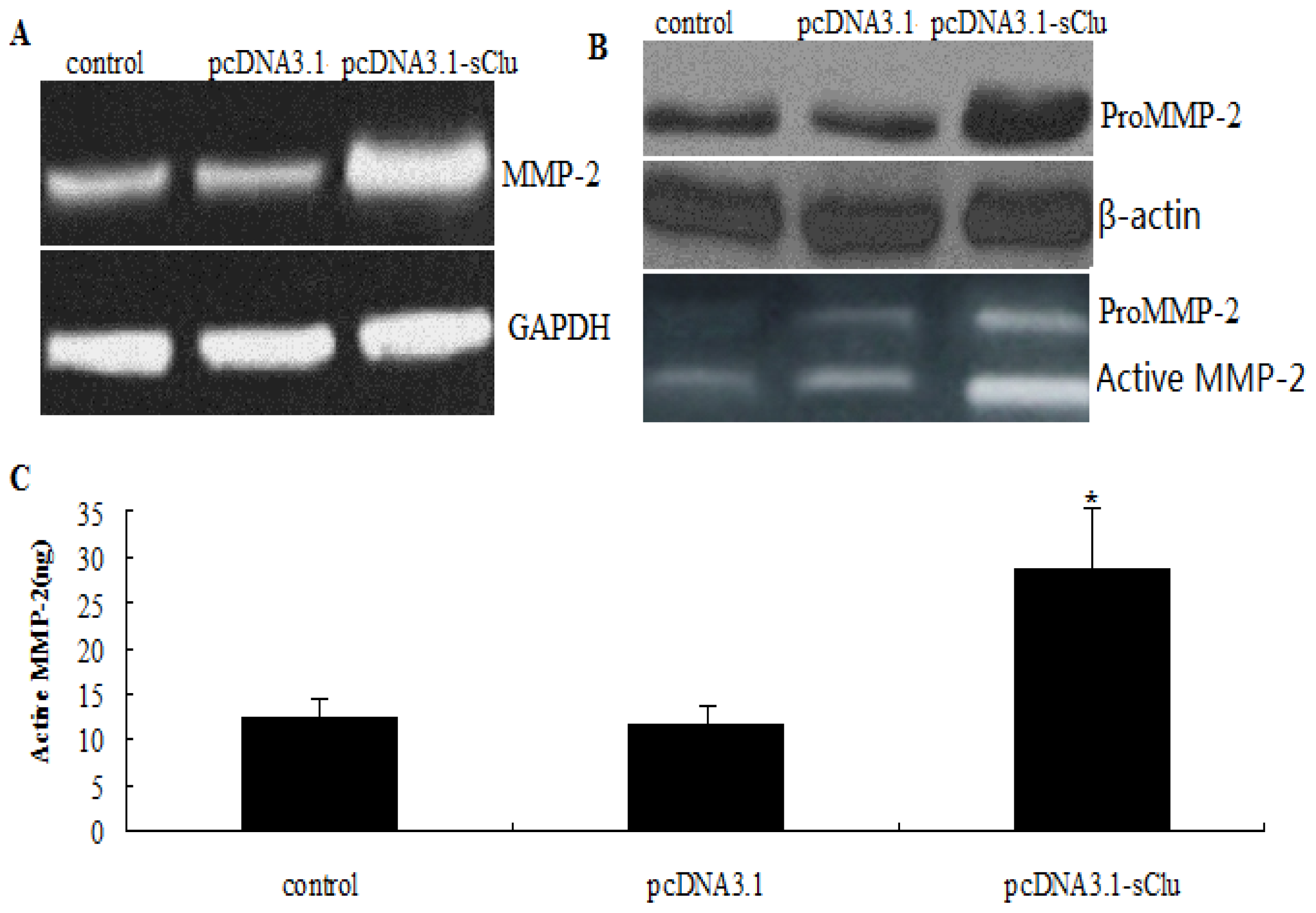
2.5. Effect of sCLU over Expression on MMP-2 Expression
2.6. Effect of sCLU Gene on E-Cadher in Expression
2.7. Discussion
3. Materials and Methods
3.1. Cell Culture
3.2. Antisense Oligonucleotide
3.3. pc.DNA3.1-sCLU Plasmid Construction and Transfection
3.4. Western Blot Analysis
3.5. Semi-Quantitative PCR
3.6. Gelatin Zymography
3.7. In Vitro Chemoinvasion Assay
3.8. Statistical Analysis
4. Conclusion
Acknowledgements
References
- Block, T.M.; Mehta, A.S.; Fimmel, C.J.; Jordan, R. Molecular viral oncology of hepatocellular carcinoma. Oncogene 2003, 22, 5093–5097. [Google Scholar]
- Okuda, K. Hepatocellular carcinoma. J. Hepatol 2000, 32, 225–237. [Google Scholar]
- Parkin, D.M.; Pisani, P.; Ferlay, J. Global cancer statistics. CA A Cancer J. Clin 1999, 49, 33–64. [Google Scholar]
- Nagao, T.; Inoue, S.; Yoshimi, F.; Sodeyama, M.; Omori, Y.; Mizuta, T.; Kawano, N.; Morioka, Y. Postoperative recurrence of hepatocellular carcinoma. Ann. Surg. 1990, 211, 28–33. [Google Scholar]
- Yang, J.; Mani, S.A.; Donaher, J.L. Twist, a master regulator of morphogenesis, plays an essential role in tumor metastasis. Cell 2004, 117, 927–939. [Google Scholar]
- Hirohashi, S. Inactivation of the E-cadherin-mediated cell adhesion system in human cancers. Am. J. Pathol 1998, 153, 333–339. [Google Scholar]
- Sugimachi, K.; Taguchi, K.; Aishima, S. Altered expression of β-catenin without genetic mutation in intrahepatic cholangiocarcinoma. Mod. Pathol 2001, 14, 900–905. [Google Scholar]
- Liu, L.; Zhu, X.D.; Wang, W.Q.; Shen, Y.; Qin, Y.; Ren, Z.G.; Sun, H.C.; Tang, Z.Y. Activation of β-catenin by hypoxia in hepatocellular carcinoma contributes to enhanced metastatic potential and poor prognosis. Clin. Canc. Res 2010, 16, 2740–2750. [Google Scholar]
- Na, D.C.; Lee, J.E.; Yoo, J.E.; Oh, B.K.; Choi, G.H.; Park, Y.N. Invasion and EMT-associated genes are up-regulated in B viral hepatocellular carcinoma with high expression of CD133-human and cell culture study. Exp. Mol. Pathol 2011, 90, 66–73. [Google Scholar]
- Chen, D.; Zheng, X.; Jiao, X.; Gao, Y.; Zhang, K.; Liang, J. Transcriptional repressor snail and metastasis in hepatocellular carcinoma. Hepatogastroenterology 2012, 59, 1359–1365. [Google Scholar]
- Jones, S.E.; Jomary, C. Clusterin. Int. J. Biochem. Cell. Biol 2002, 34, 427–431. [Google Scholar]
- Wilson, M.R.; Easterbrook-Smith, S.B. Clusterin is a secreted mammalian chaperone. Trends Biochem. Sci 2000, 25, 95–98. [Google Scholar]
- Blaschuk, O.; Burdzy, K.; Fritz, I.B. Purification and characterization of a cell-aggregating factor (clusterin), the major glycoprotein in ram rete testis fluid. J. Biol. Chem 1983, 258, 7714–7720. [Google Scholar]
- Redondo, M.; Villar, E.; Torres-Muñoz, J.; Tellez, T.; Morell, M.; Petito, C.K. Overexpression of clusterin in human breast carcinoma. Am. J. Pathol 2000, 157, 393–399. [Google Scholar]
- Xie, D.; Lau, S.H.; Sham, J.S.; Wu, Q.L.; Fang, Y.; Liang, L.Z.; Che, L.H.; Zeng, Y.X.; Guan, X.Y. Up-regulated expression of cytoplasmic clusterin in human ovarian carcinoma. Cancer 2005, 103, 277–283. [Google Scholar]
- Chen, X.; Halberg, R.B.; Ehrhardt, W.M.; Torrealba, J.; Dove, W.F. Clusterin as a biomarker in murine and human intestinal neoplasia. Proc. Natl. Acad. Sci. USA 2003, 100, 530–9535. [Google Scholar]
- Rodríguez-Piñeiro, A.M.; García-Lorenzo, A.; Blanco-Prieto, S.; Alvarez-Chaver, P.; Rodríguez-Berrocal, F.J.; Cadena, M.P.; Martínez-Zorzano, V.S. Secreted clusterin in colon tumor cell models and its potential as diagnostic marker for colorectal cancer. Cancer Investig 2012, 30, 72–77. [Google Scholar]
- Steinberg, J.; Oyasu, R.; Lang, S.; Sintich, S.; Rademaker, A.; Lee, C.; Kozlowski, J.M.; Sensibar, J.A. Intracellular levels of SGP-2 (Clusterin) correlate with tumor grade in prostate cancer. Clin. Cancer Res 2010, 3, 1707–1711. [Google Scholar]
- Miyake, H.; Gleave, M.E.; Arakawa, S.; Kamidono, S.; Hara, I. Introducing the clusterin gene into human renal cell carcinoma cells enhances their metastatic potential. J. Urol 2002, 167, 2203–2208. [Google Scholar]
- Bi, J.; Guo, A.L.; Lai, Y.R.; Li, B.; Zhong, J.M.; Wu, H.Q.; Xie, Z.; He, Y.L.; Lv, Z.L.; Lau, S.H.; et al. Overexpression of clusterin correlates with tumor progression, metastasis in gastric cancer: a study on tissue microarrays. Neoplasma 2010, 57, 191–197. [Google Scholar]
- Lau, S.H.; Sham, J.S.; Xie, D.; Tzang, C.H.; Tang, D.; Ma, N.; Hu, L.; Wang, Y.; Wen, J.M.; Xiao, G.; et al. Clusterin plays an important role in hepatocellular carcinoma metastasis. Oncogene 2006, 25, 1242–1250. [Google Scholar]
- Kang, Y.K.; Hong, S.W.; Lee, H.; Kim, W.H. Overexpression of clusterin in human hepatocellular carcinoma. Hum. Pathol 2004, 35, 1340–1346. [Google Scholar]
- Flanagan, L.; Whyte, L.; Chatterjee, N.; Tenniswood, M. Effects of clusterin over-expression on metastatic progression and therapy in breast cancer. BMC Cancer 2010, 22, 100–107. [Google Scholar]
- Panico, F.; Rizzi, F.; Fabbri, L.M.; Bettuzzi, S.; Luppi, F. Clusterin (CLU) and lung cancer. Adv. Cancer Res 2009, 105, 63–76. [Google Scholar]
- So, A.; Sinnemann, S.; Huntsman, D.; Fazli, L.; Gleave, M. Knockdown of the cytoprotective chaperone, clusterin, chemosensitizes human breast cancer cells both in vitro and in vivo. Mol. Cancer Ther 2005, 4, 1837–1849. [Google Scholar]
- Chou, T.Y.; Chen, W.C.; Lee, A.C.; Hung, S.M.; Shih, N.Y.; Chen, M.Y. Clusterin silencing in human lung adenocarcinoma cells induces a mesenchymal-to-epithelial transition through modulating the ERK/Slug pathway. Cell Signal 2009, 21, 704–711. [Google Scholar]
- Pan, Q.; Bao, L.W.; Merajver, S.D. Tetrathiomolybdate inhibits angiogenesis and metastasis through suppression of the NF-κB signaling cascade. Mol. Cancer Res 2003, 1, 701–706. [Google Scholar]
- Aalinkeel, R.; Nair, M.P.; Sufrin, G.; Mahajan, S.D.; Chadha, K.C.; Chawda, R.P.; Schwartz, S.A. Gene expression of angiogenic factors correlates with metastatic potential of prostate cancer cells. Cancer Res 2004, 64, 5311–5321. [Google Scholar]
- Poon, R.T.; Fan, S.T.; Wong, J. Risk factors, prevention and management of postoperative recurrence after resection of hepatocellular carcinoma. Ann. Surg 2000, 232, 10–24. [Google Scholar]
- Soo, K.; O’Rourke, M.P.; Khoo, P.L. Twist function is required for the morphogenesis of the cephalic neural tube and the differentiation of the cranial neural crest cells in the mouse embryo. Dev. Biol. 2002, 247, 251–270. [Google Scholar]
- Zhao, X.L.; Sun, T.; Che, N.; Sun, D.; Zhao, N.; Dong, X.Y.; Gu, Q.; Yao, Z.; Sun, B.C. Promotion of hepatocellular carcinoma metastasis through matrix metalloproteinase activation by epithelial-mesenchymal transition regulator Twist1. Cell Mol. Med 2011, 15, 691–700. [Google Scholar]
- Sawada, S.; Murakami, K.; Murata, J.; Tsukada, K.; Saiki, I. Accumulation of extracellular matrix in the liver induces high metastatic potential of hepatocellular carcinoma to the lung. Int. J. Oncol 2001, 1965–1970. [Google Scholar]
- Thiery, J.P. Epithelial-mesenchymal transitions in tumor progression. Nat. Rev. Cancer 2002, 2, 2442–2454. [Google Scholar]
- Huber, G.F.; Züllig, L.; Soltermann, A.; Roessle, M.; Graf, N.; Haerle, S.K.; Studer, G.; Jochum, W.; Moch, H.; Stoeckli, S.J. Down regulation of E-Cadherin (ECAD)—A predictor for occult metastatic disease in sentinel node biopsy of early squamous cell carcinomas of the oral cavity and oropharynx. BMC Cancer 2011, 11, 217:1–217:8. [Google Scholar]
- Chen, X.; Wang, Y.; Xia, H.; Wang, Q.; Jiang, X.; Lin, Z.; Ma, Y.; Yang, Y.; Hu, M. Loss of E-cadherin promotes the growth, invasion and drug resistance of colorectal cancer cells and is associated with liver metastasis. Mol. Biol. Rep 2012, 39, 6707–6714. [Google Scholar]
- Wells, A.; Yates, C.; Shepard, C.R. E-cadherin as an indicator of mesenchymal to epithelial reverting transitions during the metastatic seeding of disseminated carcinomas. Clin. Exp. Metastasis 2008, 25, 621–628. [Google Scholar]
- Zhai, B.; Yan, H.X.; Liu, S.Q.; Chen, L.; Wu, M.C.; Wang, H.Y. Reduced expression of E-cadherin/catenin complex in hepatocellular carcinomas. World J. Gastroenterol 2008, 14, 5665–5673. [Google Scholar]


© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chen, D.; Wang, Y.; Zhang, K.; Jiao, X.; Yan, B.; Liang, J. Antisense Oligonucleotide Against Clusterin Regulates Human Hepatocellular Carcinoma Invasion Through Transcriptional Regulation of Matrix Metalloproteinase-2 and E-Cadherin. Int. J. Mol. Sci. 2012, 13, 10594-10607. https://doi.org/10.3390/ijms130810594
Chen D, Wang Y, Zhang K, Jiao X, Yan B, Liang J. Antisense Oligonucleotide Against Clusterin Regulates Human Hepatocellular Carcinoma Invasion Through Transcriptional Regulation of Matrix Metalloproteinase-2 and E-Cadherin. International Journal of Molecular Sciences. 2012; 13(8):10594-10607. https://doi.org/10.3390/ijms130810594
Chicago/Turabian StyleChen, Dong, Yan Wang, Kejun Zhang, Xuelong Jiao, Bomin Yan, and Jun Liang. 2012. "Antisense Oligonucleotide Against Clusterin Regulates Human Hepatocellular Carcinoma Invasion Through Transcriptional Regulation of Matrix Metalloproteinase-2 and E-Cadherin" International Journal of Molecular Sciences 13, no. 8: 10594-10607. https://doi.org/10.3390/ijms130810594