Gender Related Differences in Kidney Injury Induced by Mercury

Abstract
:1. Introduction
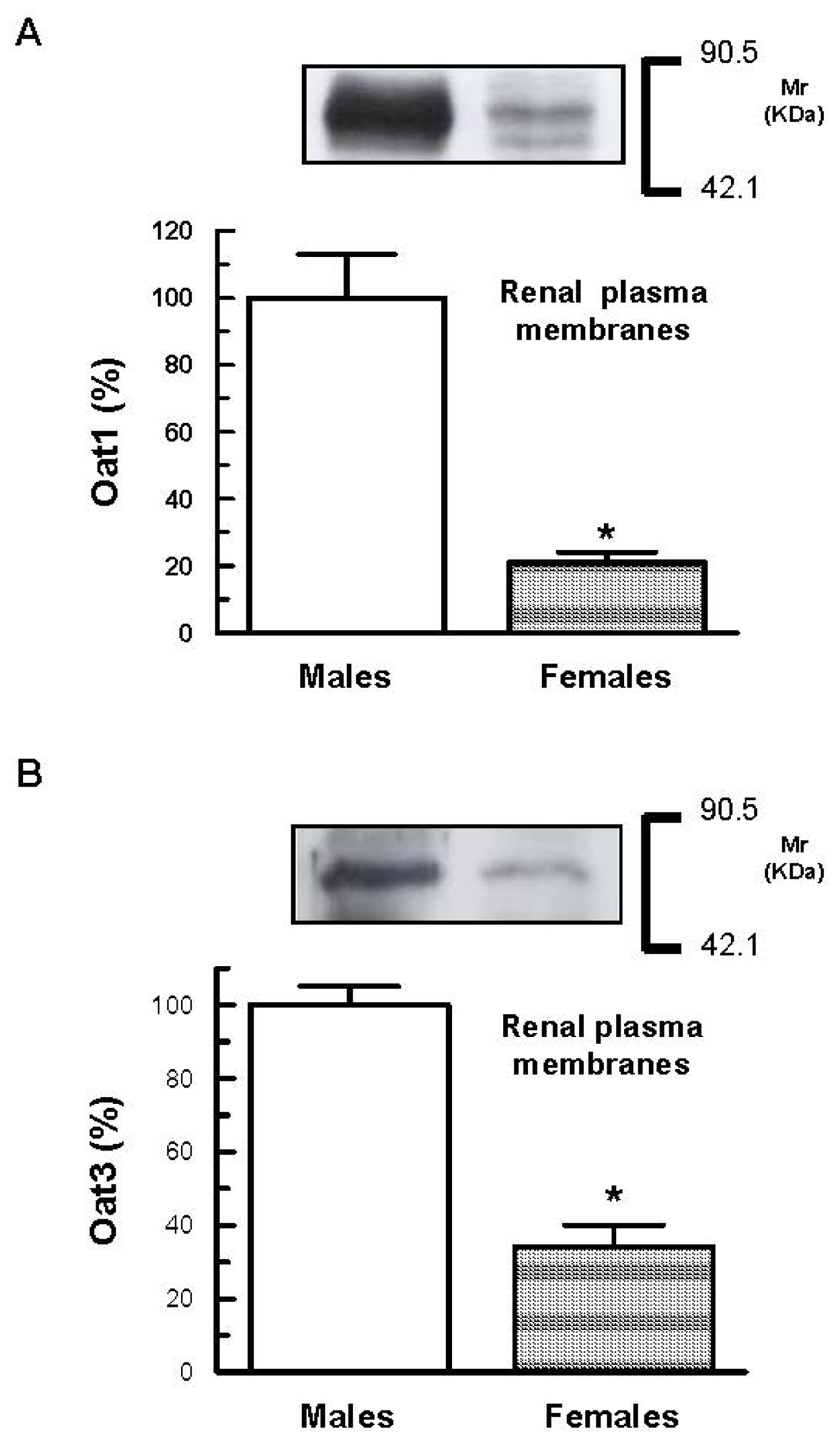
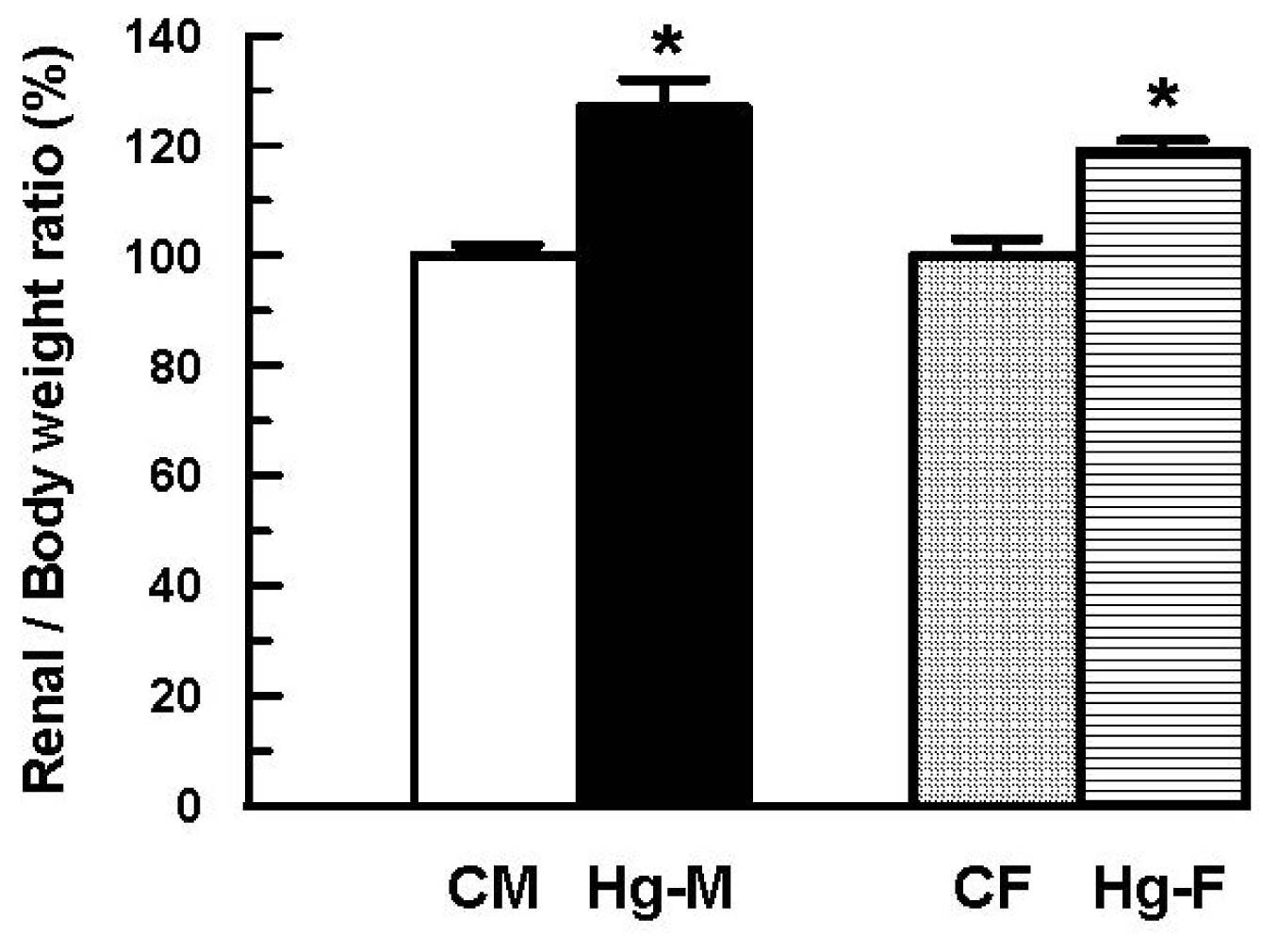
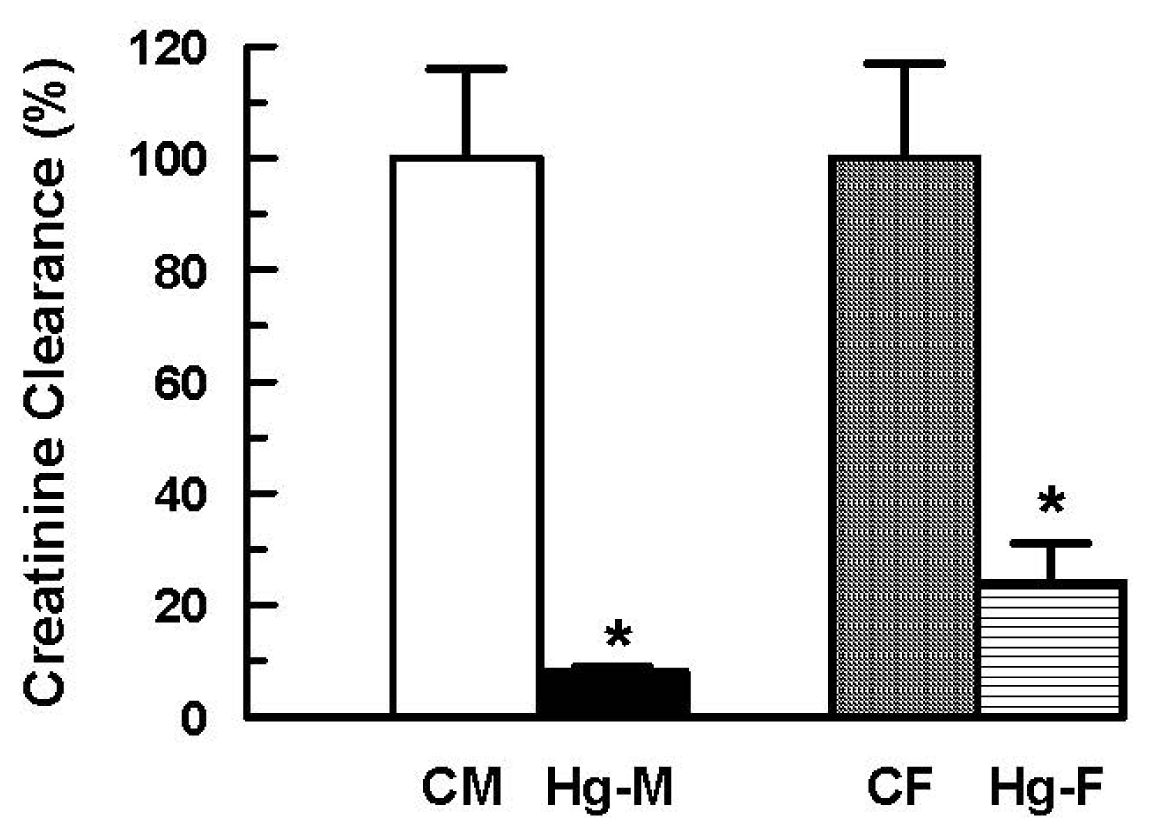
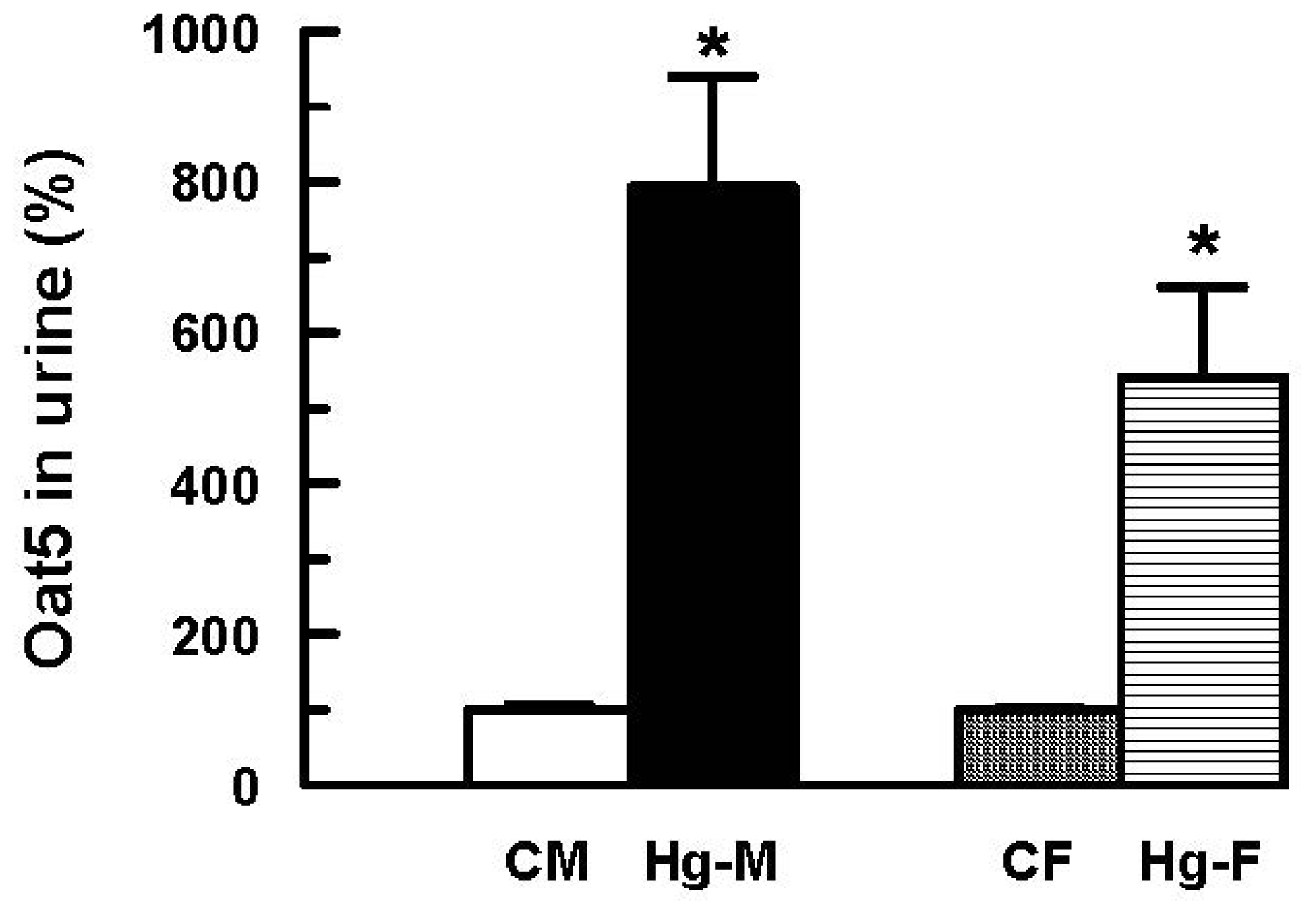
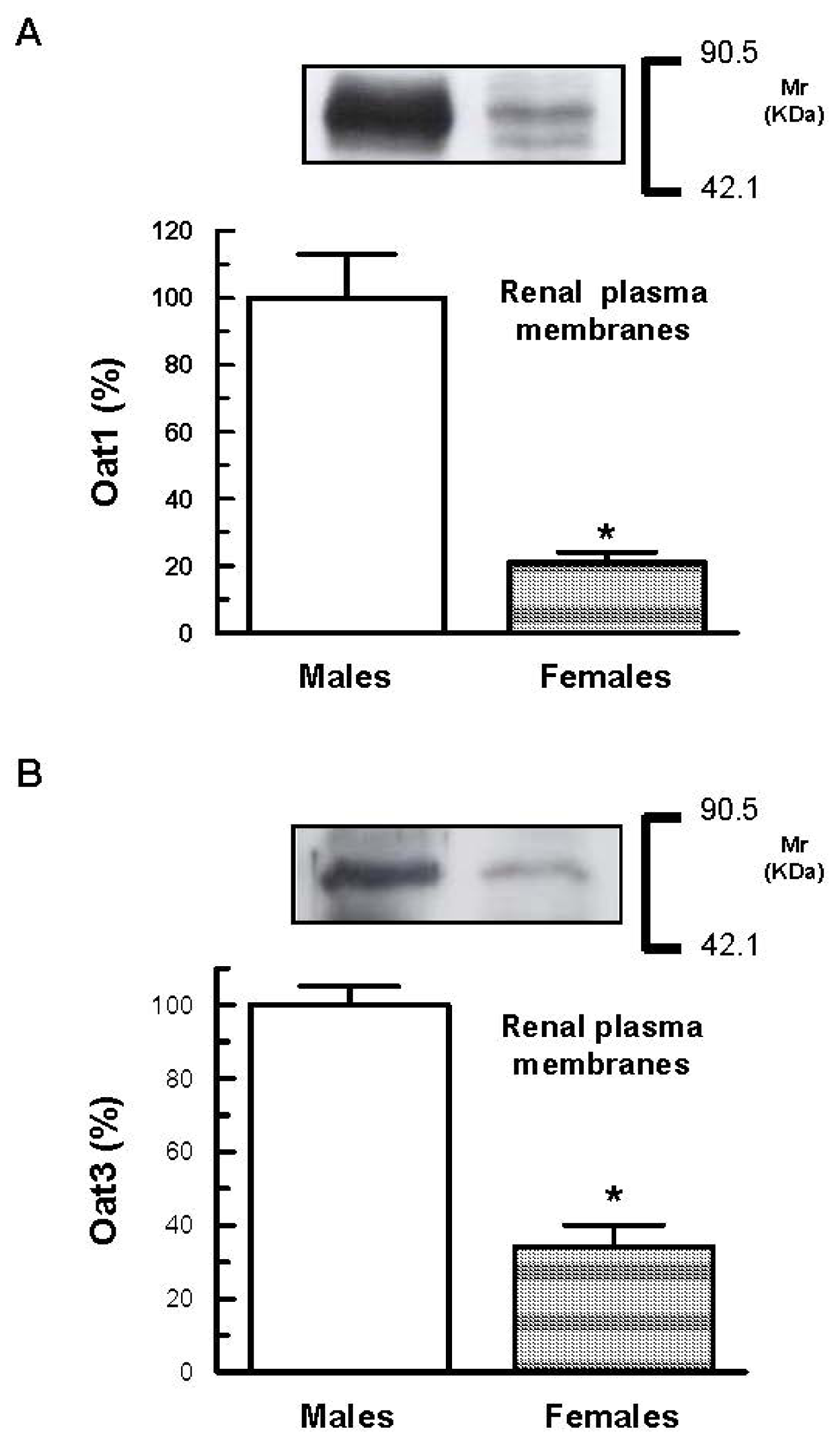
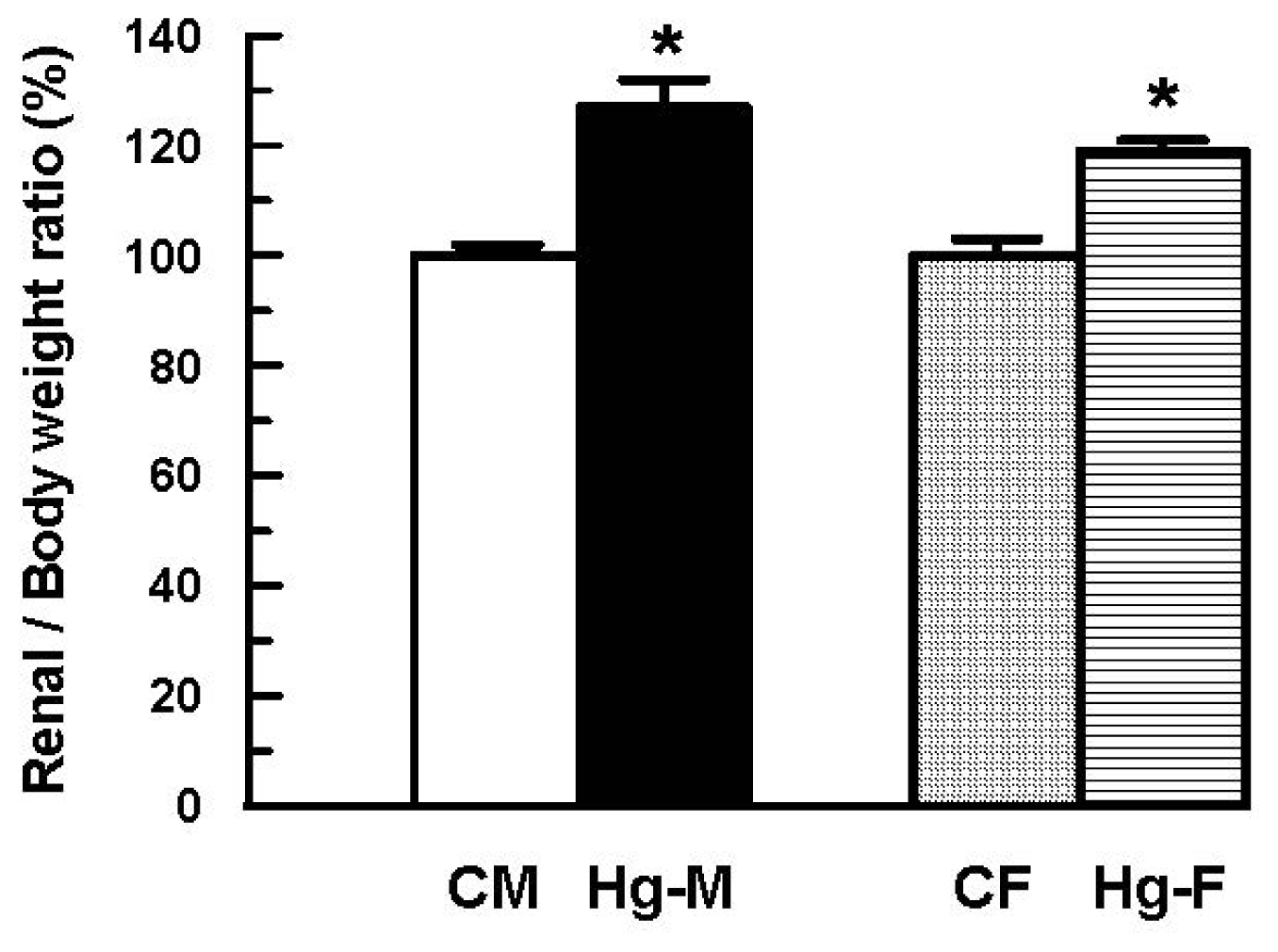
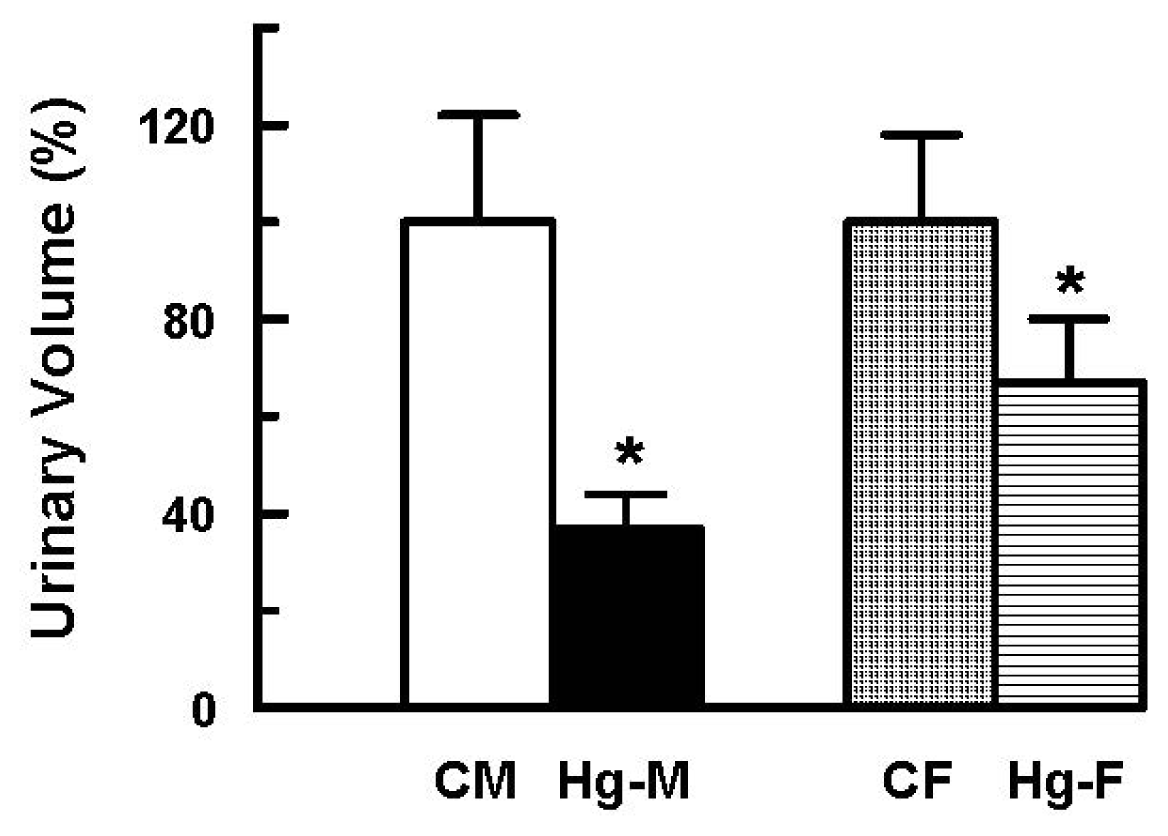
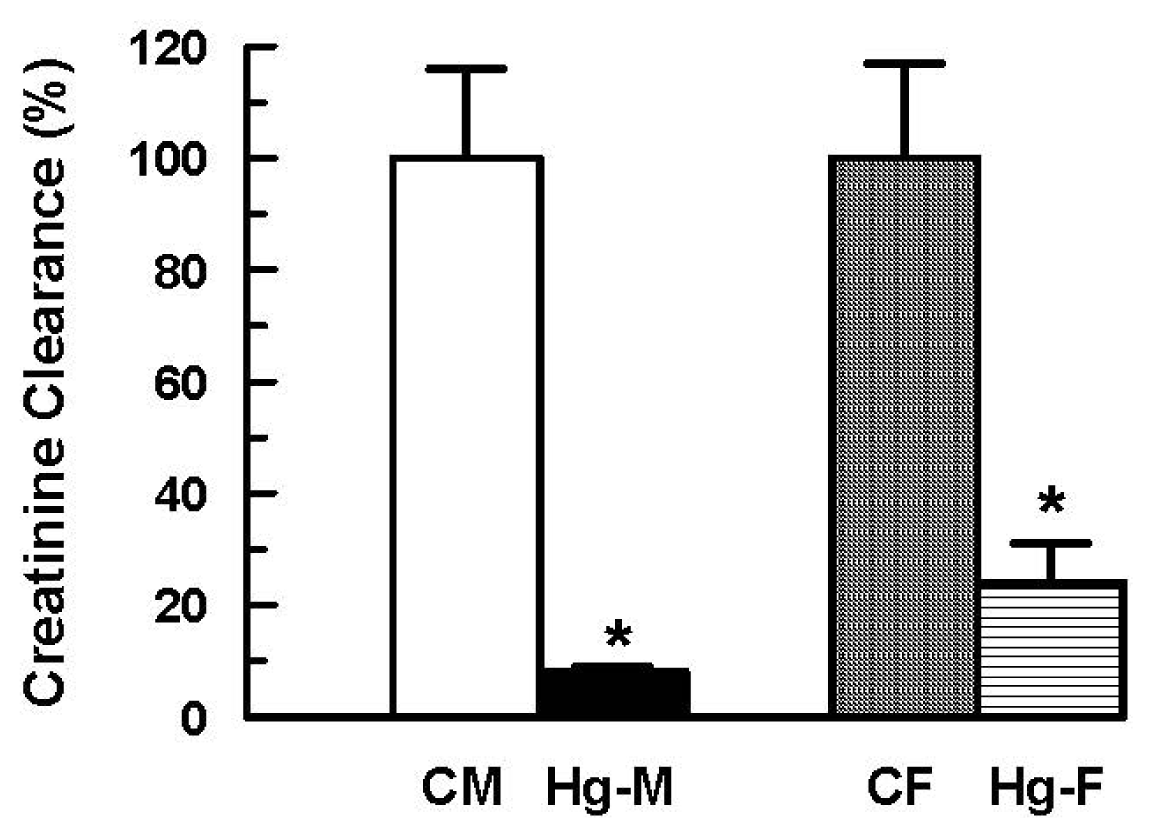
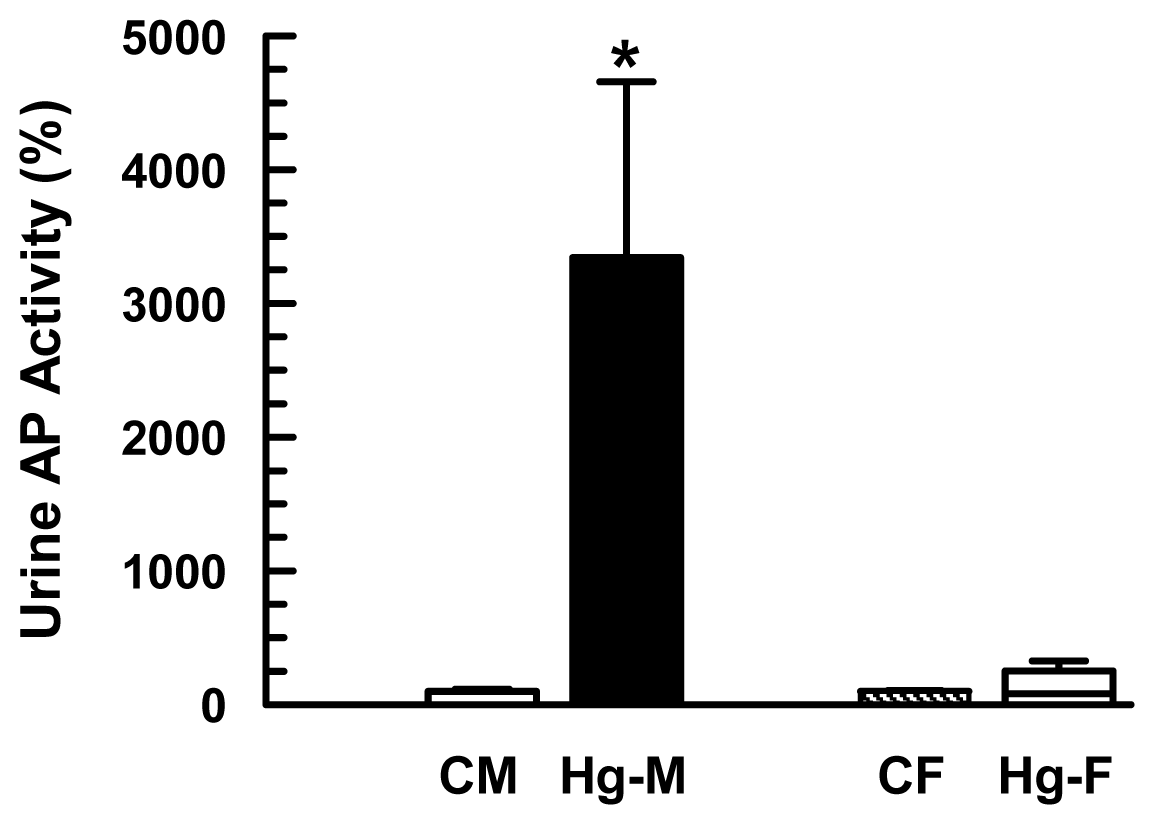
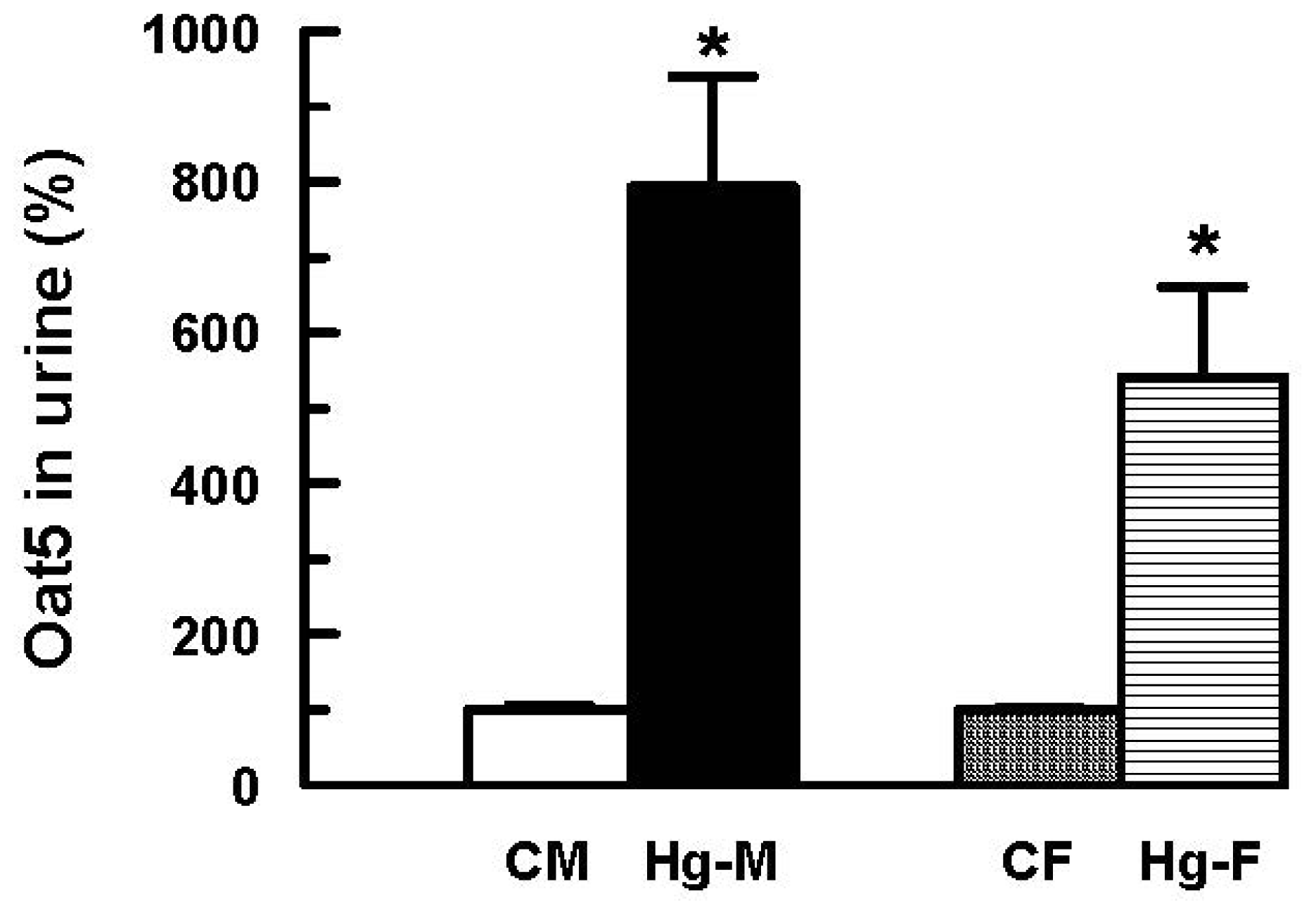
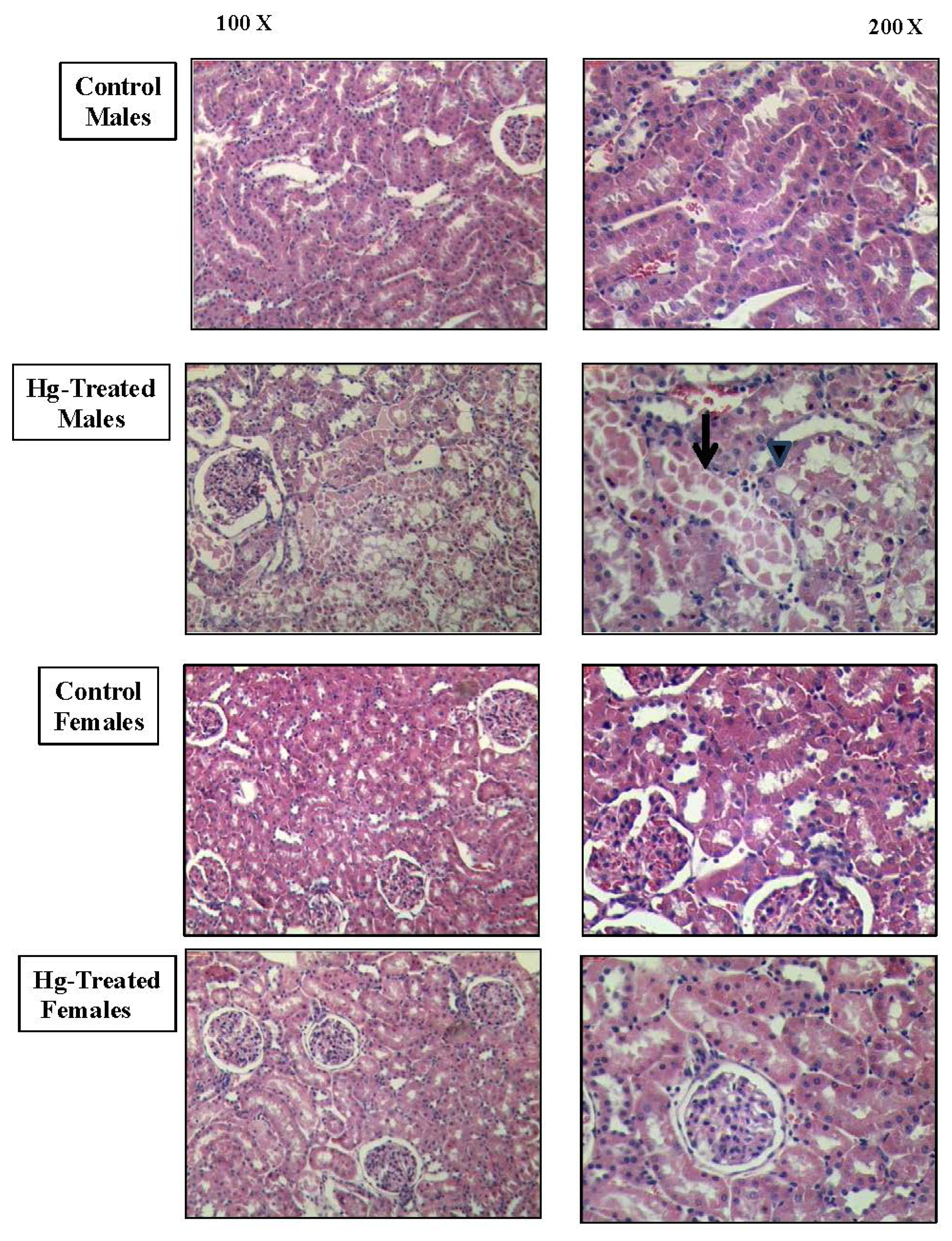
2. Results and Discussion
3. Experimental Section
3.1. Animals and Experimental Protocols
3.2. Biochemical Determinations
3.3. Preparation of Total Plasma Membranes from Kidneys
3.4. Electrophoresis and Immunoblotting
3.5. Histopathological Studies
3.6. Materials
3.7. Statistical Analysis
4. Conclusion
Acknowledgements
References
- WHO, Inorganic Mercury. In Environmental Health Criteria; Volume 118, World Health Organization: Geneva, Switzerland; p. 1991.
- Vahter, M.; Akesson, A.; Lind, B.; Björs, U.; Schütz, A.; Berglund, M. Longitudinal study of methylmercury and inorganic mercury in blood and urine of pregnant and lactating women, as well as umbilical cord blood. Environ. Res 2000, 84, 186–194. [Google Scholar]
- McRill, C.; Boyer, L.V.; Flood, T.J.; Ortega, L. Mercury toxicity due to the use of a cosmetic cream. J. Occup. Environ. Med 2000, 42, 4–7. [Google Scholar]
- Li, A.M.; Chan, M.H.M.; Leung, T.F.; Cheung, R.C.K.; Lam, C.W.K.; Fok, T.F. Mercury intoxication presenting with tics. Arch. Dis. Child 2000, 83, 174–175. [Google Scholar]
- Hultman, P.; Nielsen, J.B. The effect of dose, gender, and non-H-2 genes in murine mercury-induced autoimmunity. J. Autoimm 2001, 17, 27–37. [Google Scholar]
- Akesson, I.; Schutz, A.; Attewell, R.; Skerfving, S.; Glantz, P.O. Status of mercury and selenium in dental personnel: Impact of amalgam work and own fillings. Arch. Environ. Health 1991, 46, 102–109. [Google Scholar]
- Thomas, D.J.; Fisher, H.L.; Sumler, M.R.; Mushak, P.; Hall, I.I. Sexual differences in the excretion of organic and inorganic mercury by methyl mercury-treated rats. Environ. Res 1987, 43, 203–227. [Google Scholar]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F. Cognitive performance of children preferentially exposed to “safe” levels of methylmercury. Environ. Res 1998, 77, 165–172. [Google Scholar]
- McKeown-Eyssen, G.E.; Ruedy, J.; Neims, A. Methyl mercury exposure in northern Quebec. II. Neurologic findings in children. Am. J. Epidemiol 1983, 118, 470–479. [Google Scholar]
- Gimenez-Llort, L.; Ahlbom, D.; Daré, E.; Vahter, M.; Ogren, S.; Ceccatelli, S. Prenatal exposure to methylmercury changes dopamine-modulated motor activity during early ontogeny: Age and gender-dependent effects. Environ. Toxicol. Pharmacol 2001, 9, 61–70. [Google Scholar]
- Rossi, A.D.; Ahlbom, E.; Ogren, S.O.; Nicotera, P.; Ceccatelli, S. Prenatal exposure to methylmercury alters locomotor activity of male but not female rats. Exp. Brain. Res 1997, 117, 428–436. [Google Scholar]
- Vahter, M.; Akesson, A.; Lidén, C.; Seccatelli, S.; Berglund, M. Gender differences in the disposition and toxicity of metals. Environ. Res 2007, 104, 85–95. [Google Scholar]
- Emanuelli, T.; Rocha, J.B.T.; Pereira, M.E.; Porciúncula, L.O.; Morsch, V.M.; Martins, A.F.; Souza, D.O. Effect of mercuric chloride intoxication and dimercaprol treatment on σ-aminolevulinate dehydratase from brain, liver and kidney of adult mice. Pharmacol. Toxicol 1996, 79, 136–143. [Google Scholar]
- Tanaka-Kagawa, T.; Suzuki, M.; Naganuma, A.; Yamanaka, N.; Imura, N. Strain difference in sensitivity of mice to renal toxicity of inorganic mercury. J. Pharmacol. Exp. Ther 1998, 285, 335–341. [Google Scholar]
- Zalups, R.K. Molecular interactions with mercury in the kidney. Pharmacol. Rev 2000, 52, 113–143. [Google Scholar]
- Bridges, C.C.; Zalups, R.K. Transport of inorganic mercury and methylmercury in target tissues and organs. J. Toxicol. Environ. Health Crit. Rev 2010, 13, 385–410. [Google Scholar]
- Zalups, R.K. Basolateral uptake of mercuric conjugates of N-acetylcysteine and cysteine in the kidney involves the organic anion transport system. J. Toxicol. Environ. Health 1998, 53, 615–636. [Google Scholar]
- Bridges, C.C.; Zalups, R.K. Molecular and ionic mimicry and the transport of toxic metals. Toxicol. Appl. Pharmacol 2005, 204, 274–308. [Google Scholar]
- Bridges, C.C.; Joshee, L.; Zalups, R.K. Multidrug resistance proteins and the renal elimination of inorganic mercury mediated by 2,3-dimercaptopropane-1-sulfonic acid and meso-2,3-dimercaptosuccinic acid. J. Pharmacol. Exp. Ther 2008, 324, 383–390. [Google Scholar]
- Torres, A.M.; Dnyanmote, A.V.; Bush, K.T.; Wu, W.; Nigam, S.K. Deletion of multispecific organic anion transporter (Oat1/Slc22a6) protects from mercury-induced kidney injury. J. Biol. Chem 2011, 286, 26391–26395. [Google Scholar]
- Buist, S.C.; Klaassen, C.D. Rat and mouse differences in gender-predominant expression of organic anion transporter (Oat1-3; Slc22a6-8) mRNA levels. Drug Metab. Dispos 2004, 32, 620–625. [Google Scholar]
- Cerrutti, J.A.; Brandoni, A.; Quaglia, N.B.; Torres, A.M. Sex differences is p-aminohippuric acid transport in rat kidney: Role of membrane fluidity and expression of OAT1. Mol. Cell. Biochem 2002, 233, 175–179. [Google Scholar]
- Ljubojevic, M.; Herak-Kramberger, C.M.; Hagos, Y.; Dahn, A.; Endou, H.; Burckhardt, G.; Sabolic, I. Rat renal cortical Oat1 and Oat3 exhibit gender differences determined by both androgen stimulation and estrogen inhibition. Am. J. Physiol 2004, 287, F124–F138. [Google Scholar]
- Cerrutti, J.A.; Quaglia, N.B.; Torres, A.M. Characterization of the mechanisms involved in the gender differences in p-aminohippurate renal elimination in rats. Can. J. Physiol. Pharmacol 2001, 79, 805–813. [Google Scholar]
- Munger, K.; Baylis, C. Sex differences in renal hemodynamics in rats. Am. J. Physiol 1988, 254, F223–F231. [Google Scholar]
- Remuzzi, A.; Puntorieri, S.; Mazzoleni, A.; Remuzzi, G. Sex related differences in glomerular ultrafiltration and proteinuria in Munich-Wistar rats. Kidney Int 1988, 34, 481–486. [Google Scholar]
- Heiene, R.; Moe, L.; Moelmen, G. Calculation of urinary enzyme excretion, with renal structure and function in dogs with pyometra. Res. Vet. Sci 2001, 70, 129–137. [Google Scholar]
- Price, R.G. Urinary enzymes, nephrotoxicity and renal disease. Toxicology 1982, 23, 99–134. [Google Scholar]
- Di Giusto, G.; Anzai, N.; Endou, H.; Torres, A.M. Oat5 and NaDC1 protein abundance in kidney and urine after renal ischemic reperfusion injury. J. Histochem. Cytochem 2009, 57, 17–27. [Google Scholar]
- Di Giusto, G.; Torres, A.M. Organic anion transporter 5 renal expression and urinary excretion in rats exposed to mercuric chloride: A potential biomarker of mercury-induced nephropathy. Arch. Toxicol 2010, 84, 741–749. [Google Scholar]
- Stacchiotti, A.; Ricci, F.; Rezzani, R.; Volti, G.L.; Borsani, E.; Lavazza, A.; Bianchi, R.; Rodella, L.F. Tubular stress proteins and nitric oxide synthase expression in rat kidney exposed to mercuric chloride and melatonin. J. Histochem. Cytochem 2006, 54, 1149–1157. [Google Scholar]
- Nava, M.; Romero, F.; Quiroz, Y.; Parra, G.; Bonet, L.; Rodriguez-Iturbe, B. Melatonin attenuates acute renal failure and oxidative stress induced by mercuric chloride in rats. Am. J. Physiol 2000, 279, 910–918. [Google Scholar]
- Di Giusto, G.; Anzai, N.; Ruiz, M.L.; Endou, H.; Torres, A.M. Expression and function of Oat1 and Oat3 in rat kidney exposed to mercuric chloride. Arch. Toxicol 2009, 83, 887–897. [Google Scholar]
- Wright, S.H.; Dantzler, W.H. Molecular and cellular physiology of renal organic cation and anion transport. Physiol. Rev 2004, 84, 987–1049. [Google Scholar]
- Anzai, N.; Kanai, Y.; Endou, H. Organic anion transporter family: Current knowledge. J. Pharmacol. Sci 2006, 100, 411–426. [Google Scholar]
- Rizwan, A.N.; Burckhardt, G. Organic anion transporters of the SLC22 family: Biopharmaceutical, physiological, and pathological roles. Pharm. Res 2007, 24, 450–470. [Google Scholar]
- Kojima, R.; Sekine, T.; Kawachi, M.; Cha, S.H.; Suzuki, Y.; Endou, H. Immunolocalization of multispecific organic anion transporters, OAT1, OAT2, and OAT3, in rat kidney. J. Am. Soc. Nephrol 2002, 13, 848–857. [Google Scholar]
- Villar, S.R.; Brandoni, A.; Torres, A.M. Time course of organic anion excretion in rats with bilateral ureteral obstruction: Role of organic anion transporters (Oat1 and Oat3). Nephron. Physiol 2008, 110, 45–56. [Google Scholar]
- Di Giusto, G.; Anzai, N.; Endou, H.; Torres, A.M. Elimination of organic anions in response to an early stage of renal ischemia-reperfusion in rat: Role of basolateral plasma membrane transporter and cortical renal blood flow. Pharmacology 2008, 81, 127–136. [Google Scholar]
- Wen, J.G.; Frokiaer, J.; Jorgensen, T.M. Obstructive nephropathy. An update of the experimental research. Urol. Res 1999, 27, 29–39. [Google Scholar]
- Green, J.; Abassi, Z.; Winaver, J.; Skorecki, K.L. Acute Renal Failure: Clinical and Pathophysiologic Aspects. In The Kidney: Physiology and Pathophysiology, 3rd ed; Seldin, D.W., Giebisch, G., Eds.; Williams & Wilkins: Lippincott, PA, USA, 2000; pp. 2329–2373. [Google Scholar]
- Brady, H.R.; Brenner, B.M.; Lieberthal, W. Acute Renal Failure. In Brenner & Rector’s the Kidney, 7th ed; Brenner, B.M., Rector, F.C., Eds.; W.B. Saunders Company: PA, USA, 2004; pp. 1215–1292. [Google Scholar]
- Institute of Laboratory Animal Resources, Guide for the Care and Use of Laboratory Animals, 7th ed; National Academy Press: Washington, DC, USA; p. 1996.
- Jensen, R.E.; Berndt, W.O. Epinephrine and norepinephrine enhance p-aminohippurate transport into basolateral membrane vesicles. J. Pharmacol. Exp. Ther 1988, 244, 543–549. [Google Scholar]
- Villar, S.R.; Brandoni, A.; Anzai, N.; Endou, H.; Torres, A.M. Altered expression of rat renal cortical OAT1 and OAT3 in response to bilateral ureteral obstruction. Kidney Int 2005, 68, 2704–2713. [Google Scholar]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive and versatile assay for protein using Coomassie Brillant Blue G250. Anal. Biochem 1977, 79, 544–552. [Google Scholar]
- Anzai, N.; Jutabha, P.; Enomoto, A.; Yokoyama, H.; Nonoguchi, H.; Hirata, T.; Shiraya, K.; He, X.; Cha, S.H.; Takeda, M.; et al. Functional characterization of rat organic anion transporter 5 (Slc22a19) at the apical membrane of renal proximal tubules. J. Pharmacol. Exp. Ther 2005, 315, 534–544. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CM | Hg-M | CF | Hg-F | |
|---|---|---|---|---|
| Renal/Body weigth ratio × 10−3 | 6.49 ± 0.11 (100 ± 2) | 8.27 ± 0.31 * (127 ± 5 *) | 6.81 ± 0.22 (100 ± 3) | 8.12 ± 0.14 *(119 ± 2 *) |
| Urinary Volume × 10−3 (mL/min/100 g b.w.) | 2.38 ± 0.52 (100 ± 22) | 0.88 ± 0.18 * (37 ± 7 *) | 3.31 ± 0.60 (100 ± 18) | 2.22 ± 0.44 * (67 ± 13 *) |
| Creatinine clearance × 10−3 (mL/min/100 g b.w.) | 421 ± 80(100 ± 19) | 35 ± 3 * (8 ± 1 *) | 349 ± 60 (100 ± 17) | 85 ± 24 * (24 ± 7 *) |
| Urinary AP Activity (mUI/mg creatinine) | 165 ± 26(100 ± 16) | 5511 ± 2167 * (3340 ± 1313 *) | 230 ± 24 (100 ± 10) | 578 ± 169 (251 ± 73) |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hazelhoff, M.H.; Bulacio, R.P.; Torres, A.M. Gender Related Differences in Kidney Injury Induced by Mercury. Int. J. Mol. Sci. 2012, 13, 10523-10536. https://doi.org/10.3390/ijms130810523
Hazelhoff MH, Bulacio RP, Torres AM. Gender Related Differences in Kidney Injury Induced by Mercury. International Journal of Molecular Sciences. 2012; 13(8):10523-10536. https://doi.org/10.3390/ijms130810523
Chicago/Turabian StyleHazelhoff, María H., Romina P. Bulacio, and Adriana M. Torres. 2012. "Gender Related Differences in Kidney Injury Induced by Mercury" International Journal of Molecular Sciences 13, no. 8: 10523-10536. https://doi.org/10.3390/ijms130810523