Effect of Dieckol, a Component of Ecklonia cava, on the Promotion of Hair Growth


Abstract
:1. Introduction
2. Results
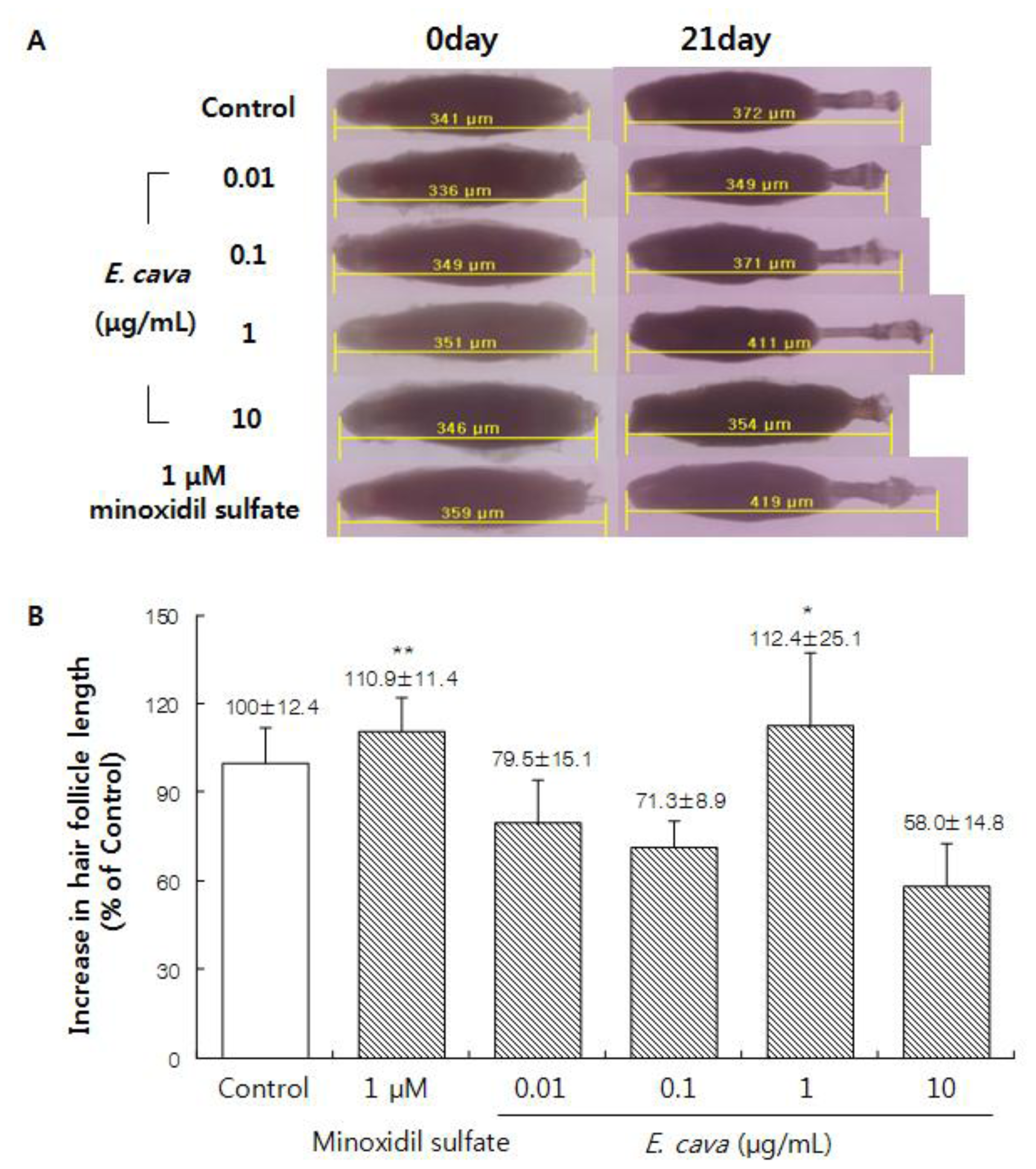
2.1. The Effect of Ecklonia cava Enzymatic Extract on the Hair-Fiber Elongation of Rat Vibrissa Follicle
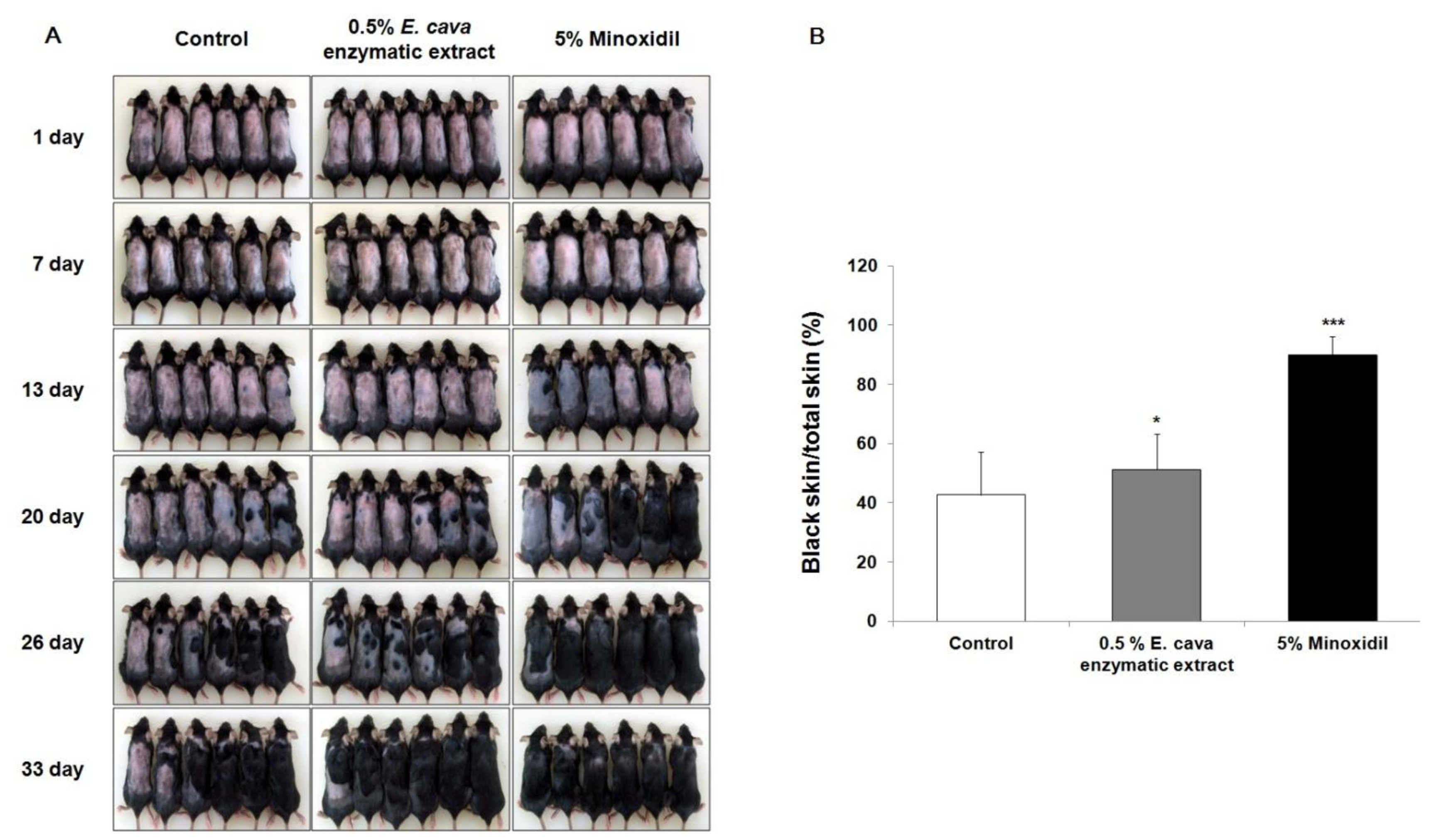
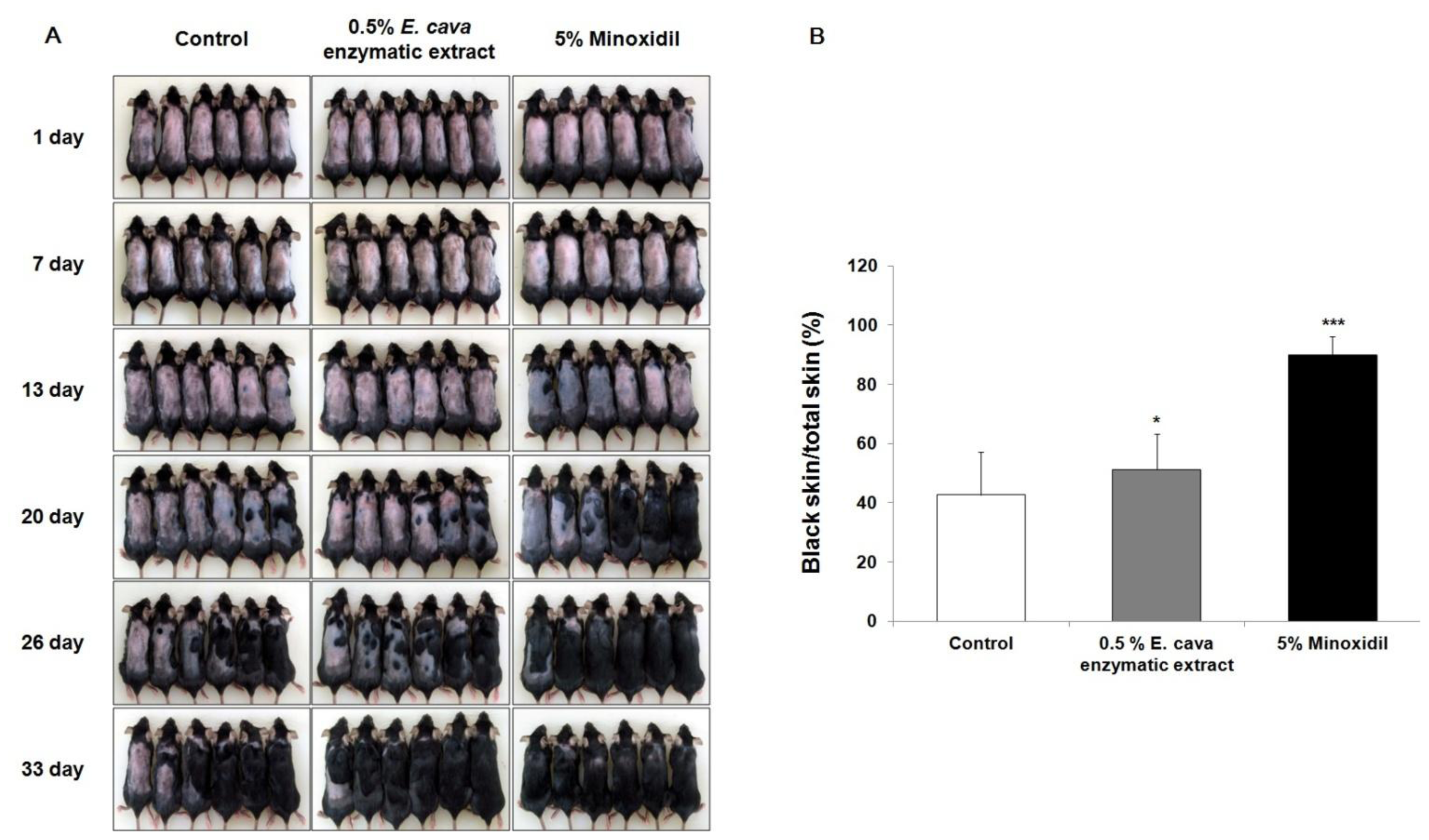
2.2. The Effect of Ecklonia cava Enzymatic Extract on the Anagen Induction in C57BL/6 Mice
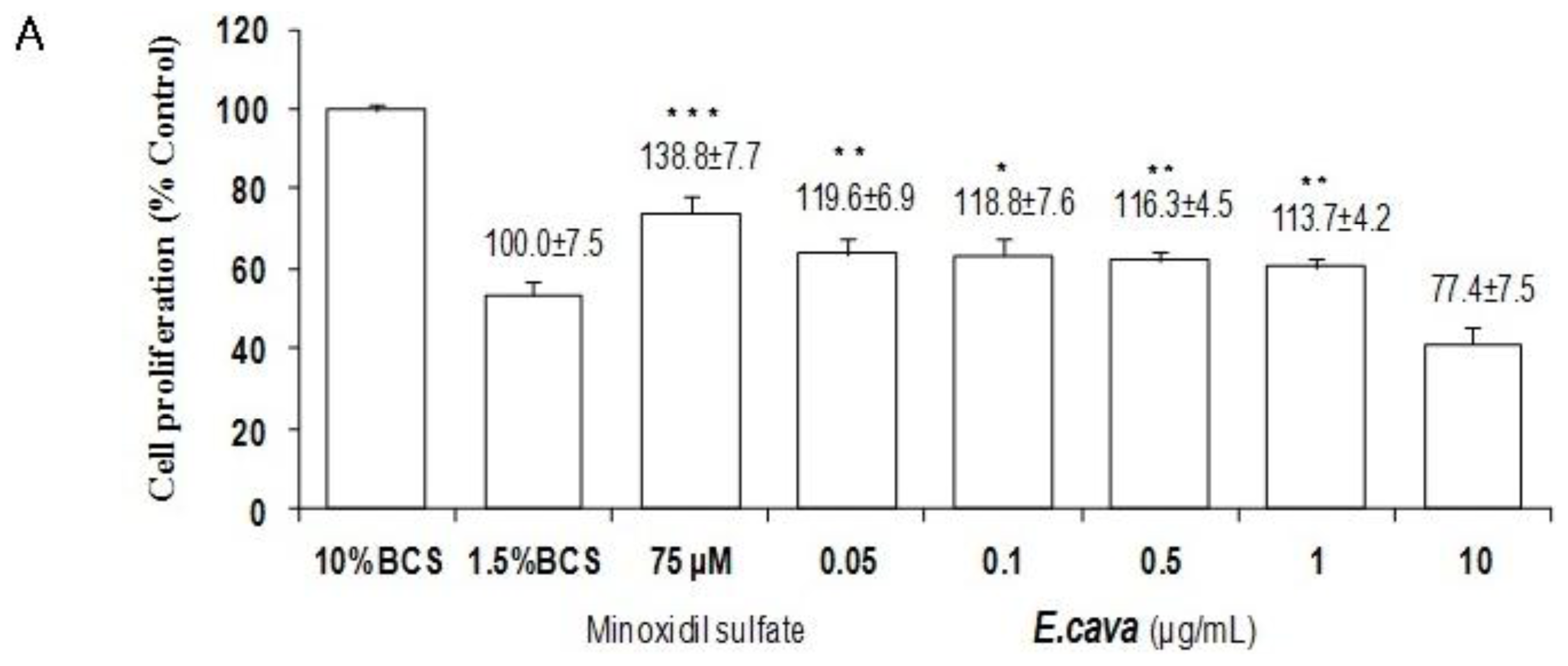
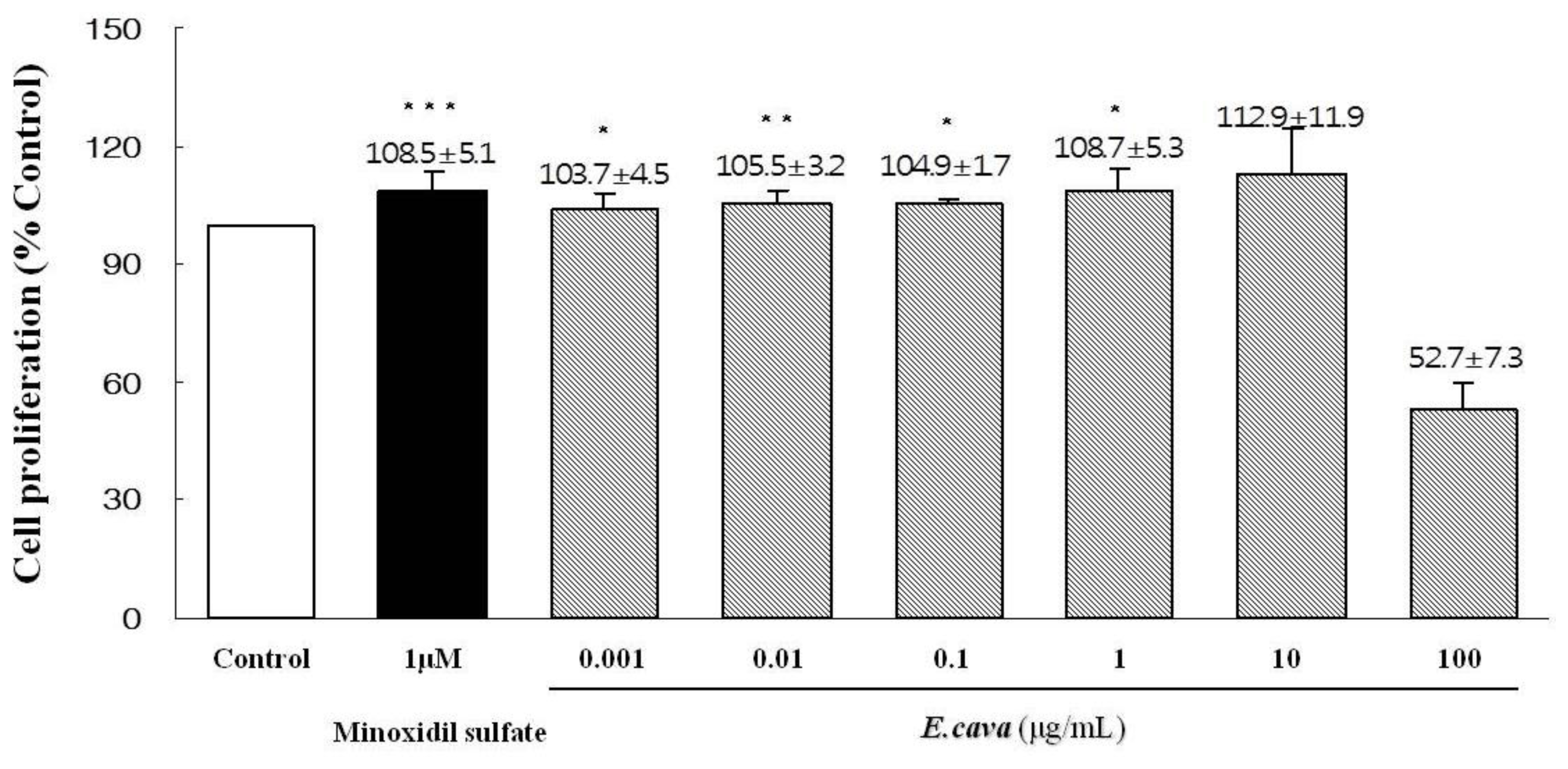
2.3. The Effects of Ecklonia cava Enzymatic Extract and Its Isolated Compounds on the Proliferation of Dermal Papilla Cells
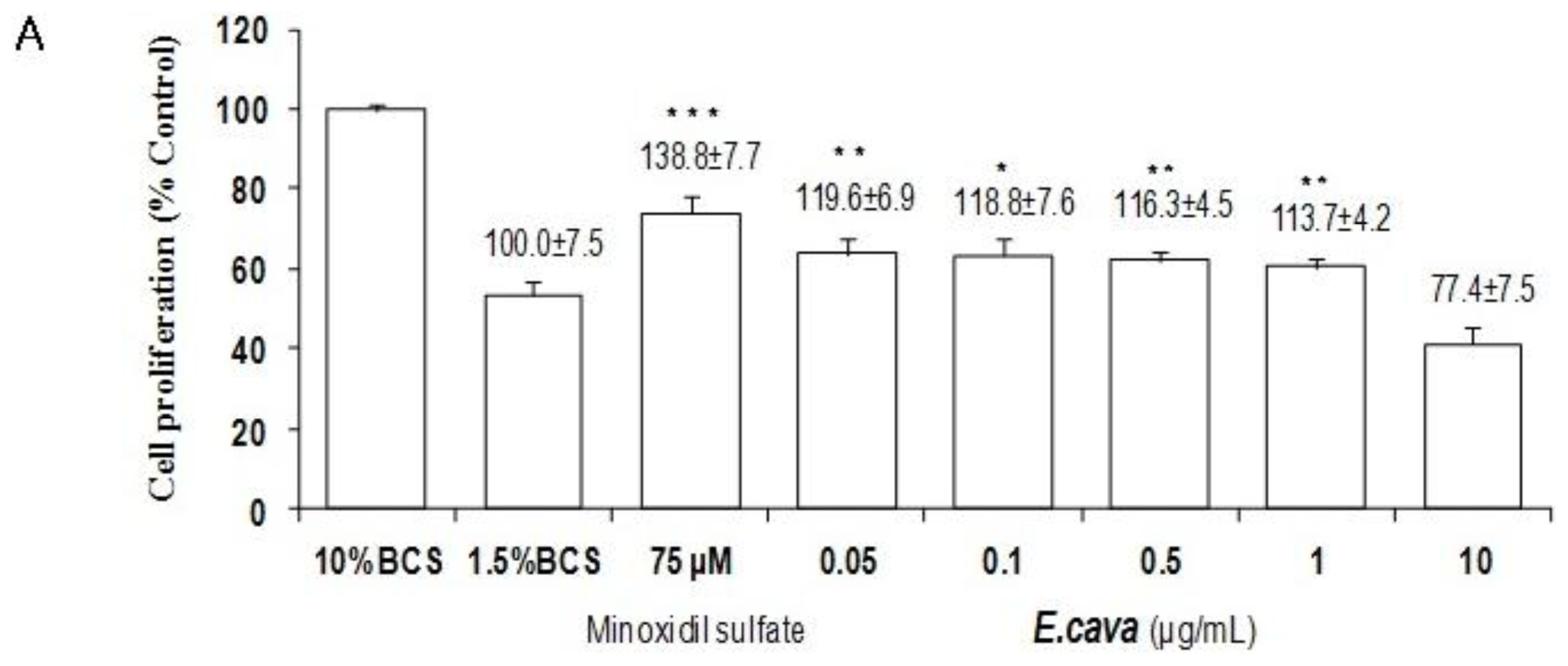
2.4. The Effects of Ecklonia cava Enzymatic Extract and Its Isolated Compounds on the Proliferation of NIH3T3 Fibroblasts
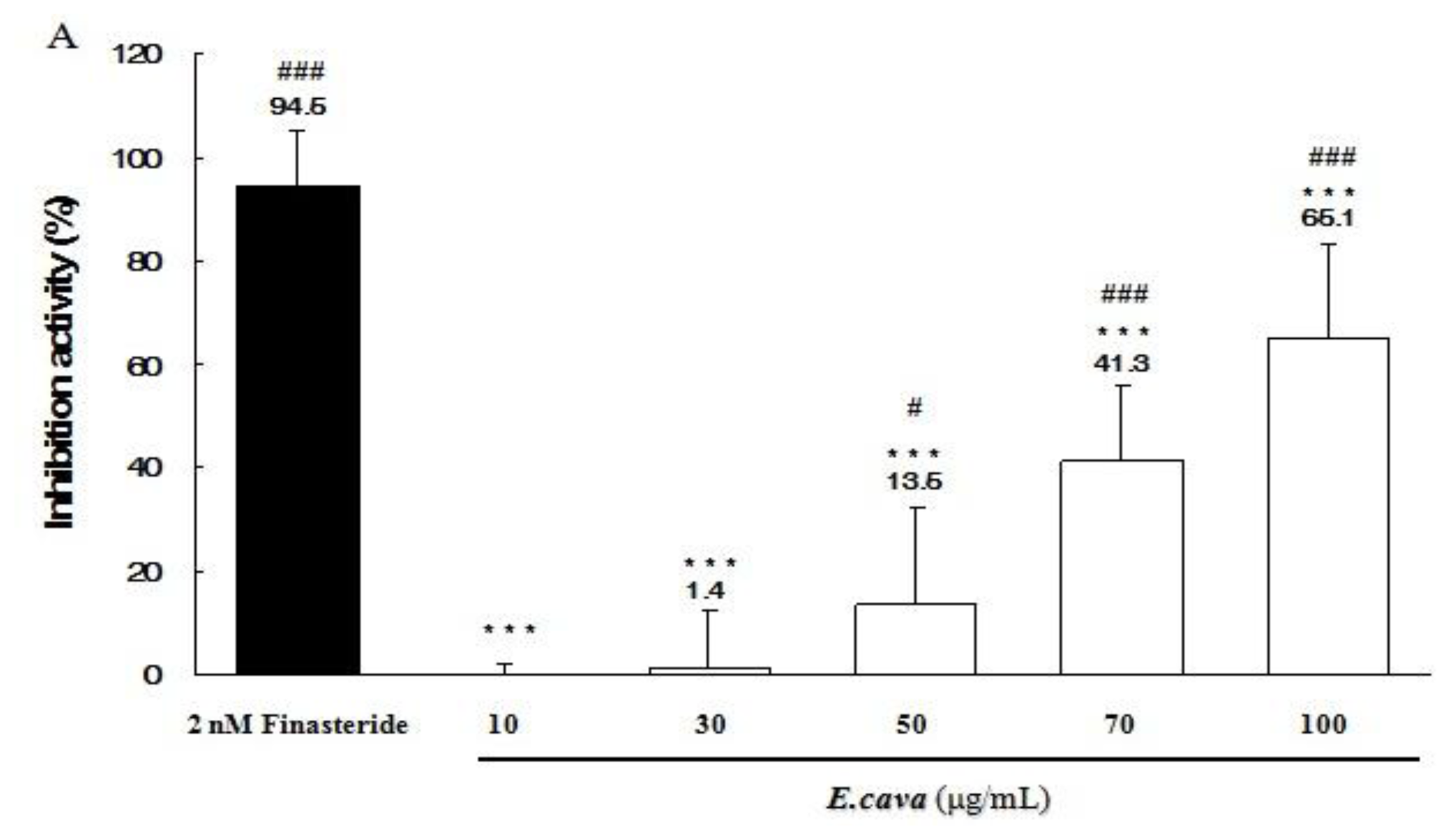
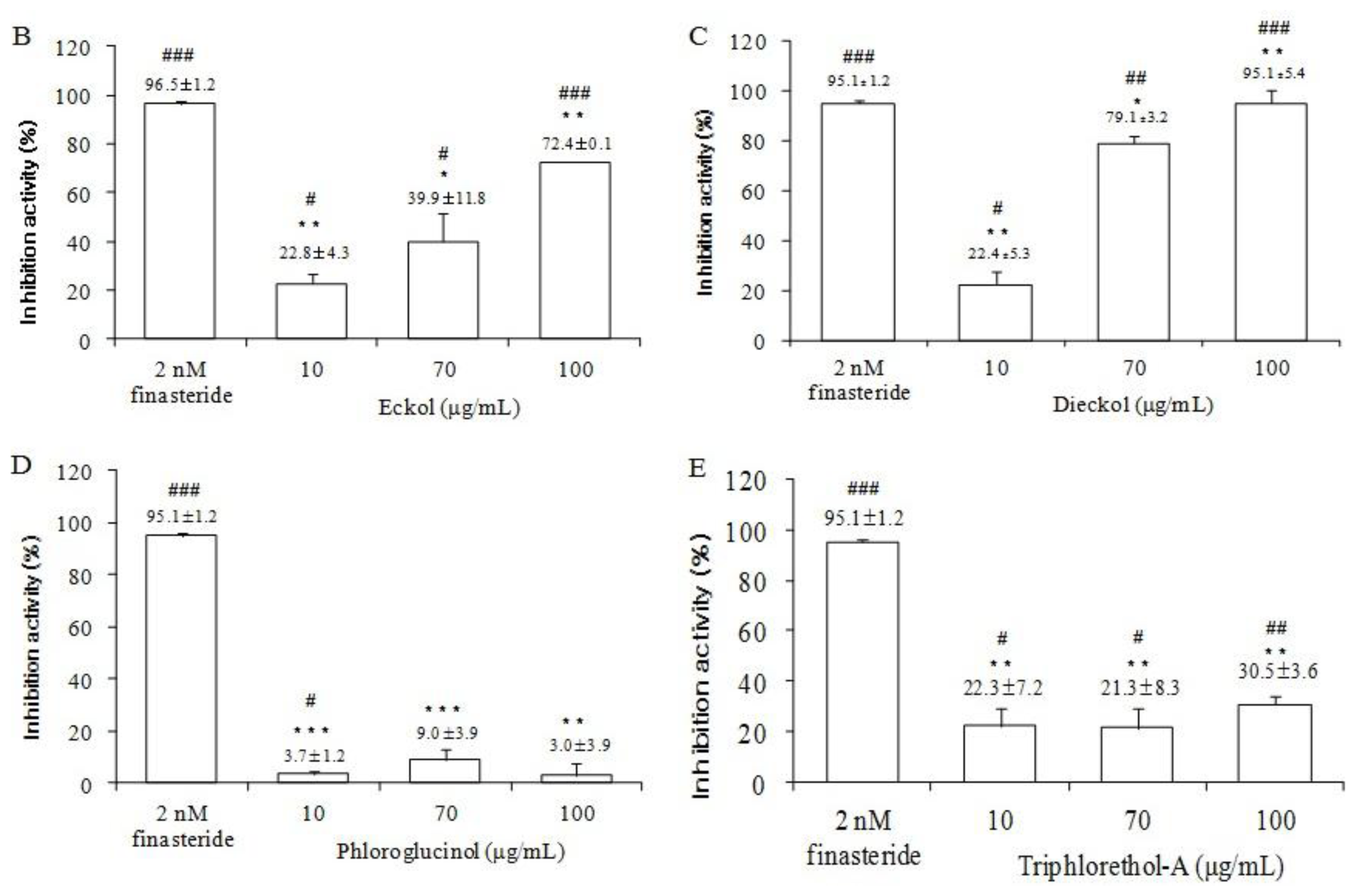
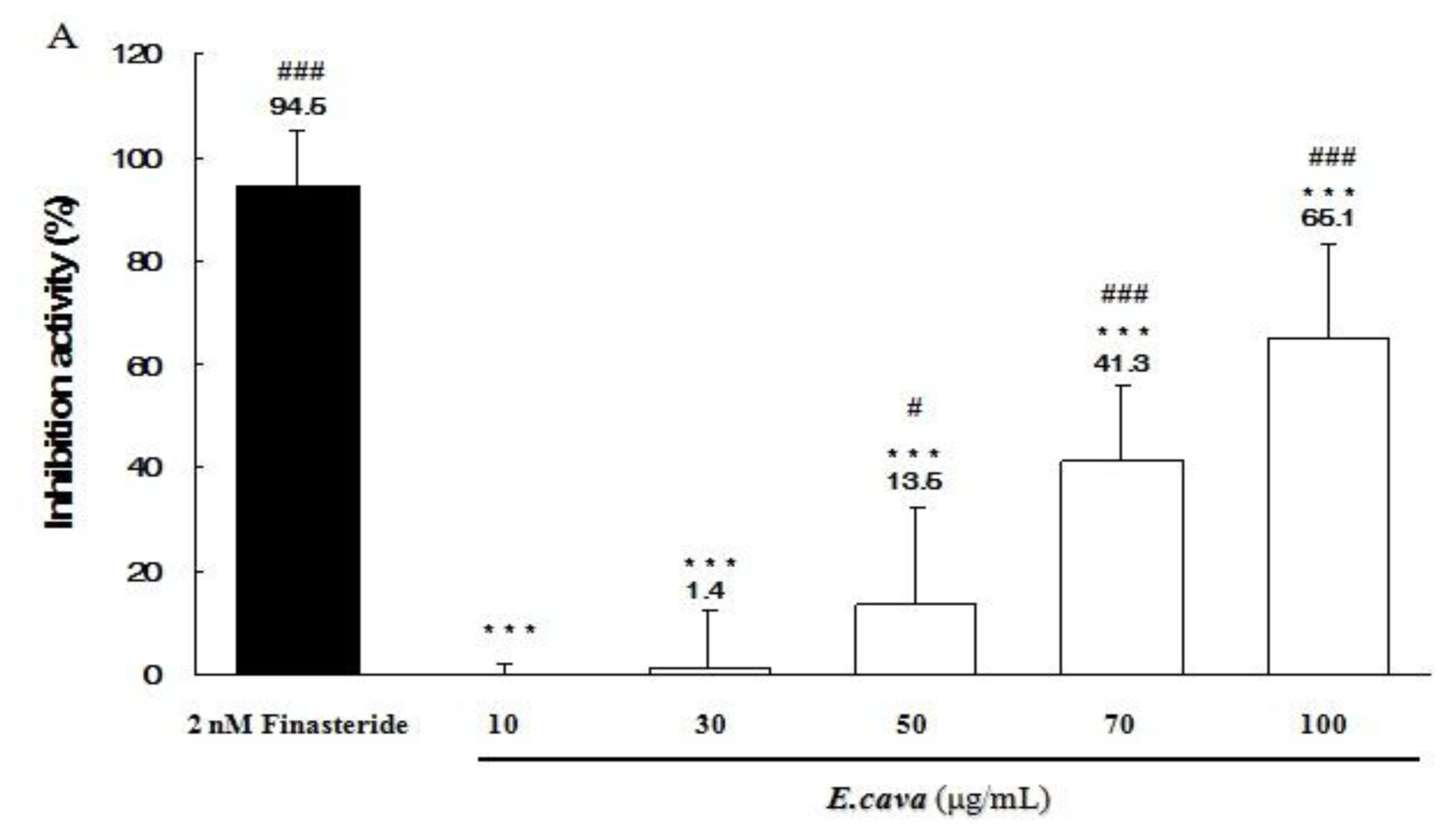
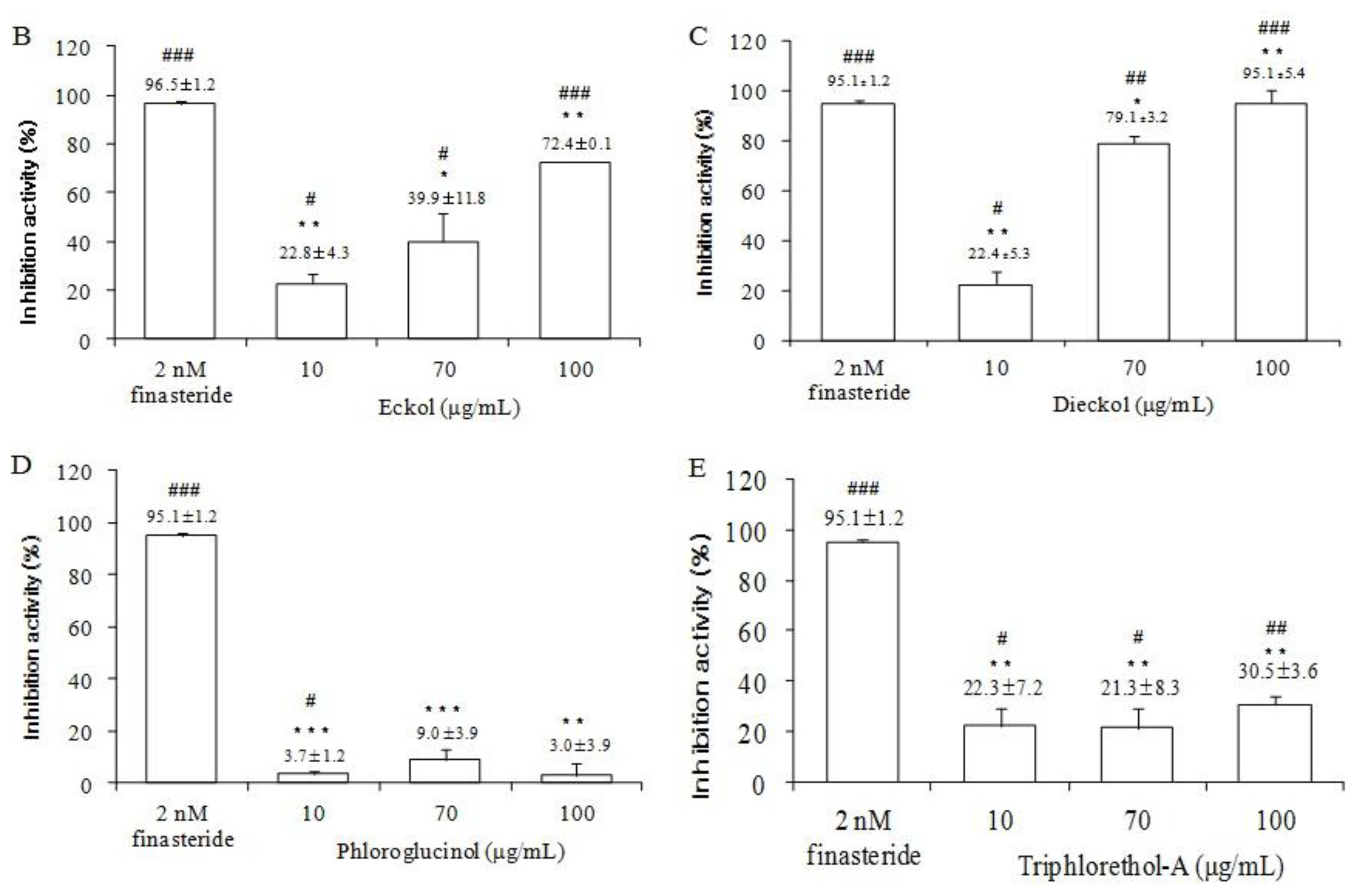
2.5. The Effects of Ecklonia cava Enzymatic Extract and Its Isolated Compounds on 5α-Reductase Activities
3. Discussion
4. Experimental Section
4.1. Alga Material
4.2. Preparation of E. cava Enzymatic Extract
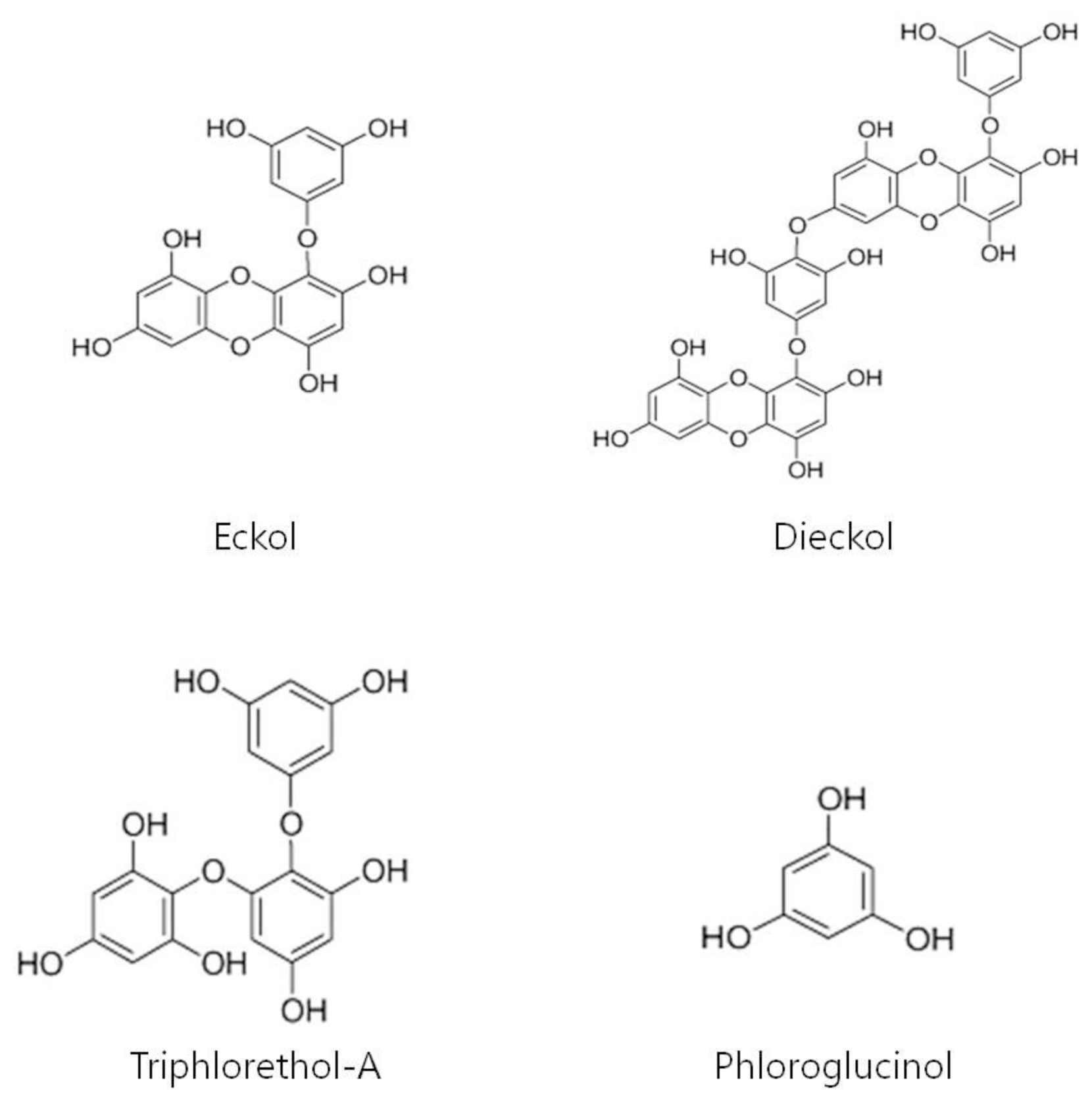
4.3. Extraction and Isolation of Phlorotannins from Ecklonia cava
4.4. Animals
4.5. Isolation and Culture of Rat Vibrissa Follicles
4.6. Hair Growth Activity in Vivo
4.7. Culture and Proliferation Assay of Dermal Papilla Cells
4.8. Culture and Proliferation Assay of NIH3T3 Fibroblasts
4.9. Assay of Rat Prostatic 5α-Reductase
4.10. Statistical Analyses
5. Conclusions
Acknowledgments
References
- Cotsarelis, G.; Millar, S.E. Towards a molecular understanding of hair loss and its treatment. Trends Mol. Med 2001, 7, 293–301. [Google Scholar]
- Price, V.H. Treatment of hair loss. N. Engl. J. Med 1999, 341, 964–973. [Google Scholar]
- Kaufman, K.D.; Olsen, E.A.; Whiting, D.; Savin, R.; DeVillez, R.; Bergfeld, W.; Price, V.H.; van Neste, D.; Roberts, J.L.; Hordinsky, M.; et al. Finasteride in the treatment of men with androgenetic alopecia. Finasteride Male Pattern Hair Loss Study Group. J. Am. Acad. Dermatol 1998, 39, 578–589. [Google Scholar]
- Burton, J.L.; Marshall, A. Hypertrichosis due to minoxidil. Br. J. Dermatol 1979, 101, 593–595. [Google Scholar]
- Gromley, G.J. Finasteride: A clinical review. Biomed. Pharmacother 1995, 49, 319–324. [Google Scholar]
- McClellan, K.J.; Markham, A. Finasteride: A review of its use male pattern hair loss. Drugs 1999, 57, 111–126. [Google Scholar]
- Mehta, P.K.; Mamdani, B.; Shansky, R.M.; Mahurkar, S.D.; Dunea, G. Severe hypertension. Treatment with minoxidil. J. Am. Med. Assoc 1975, 233, 249–252. [Google Scholar]
- Tanigaki-Obana, N.; Ito, M. Effects of cepharanthine and minoxodil on proliferation, differentiation and keratinozation of cultured cells from the murine hair apparatus. Arch. Dermatol. Res 1992, 284, 290–296. [Google Scholar]
- Uno, H.; Cappas, A.; Schlagel, C. Cyclic dynamics of hair follicles and the effect of minoxidil on the bald scalps of stumptailed macaques. Am. J. Dermatopathol 1985, 7, 283–297. [Google Scholar]
- Han, J.H.; Kwon, O.S.; Chung, J.H.; Cho, K.H.; Eun, H.C.; Kim, K.H. Effect of minoxodil on proliferation and apoptosis in dermal papilla cells of human hair follicle. J. Dermatol. Sci 2004, 34, 91–98. [Google Scholar]
- Hamaoka, H.; Minakuchi, K.; Miyoshi, H.; Arase, S.; Chen, C.H.; Nakaya, Y. Effect of K+ channel openers on K+ channel in cultured human dermal papilla cells. J. Med. Invest 1997, 44, 73–77. [Google Scholar]
- Shorter, K.; Farjo, N.P.; Picksley, S.M.; Randall, V.A. Human hair follicles contain two forms of ATP-sensitive potassium channels, only one of which is sensitive to minoxidil. FASEB J 2008, 22, 1725–1736. [Google Scholar]
- Lachgar, S.; Moukadiri, H.; Jonca, F.; Charveron, M.; Bouhaddioui, N.; Gall, Y.; Bonafe, J.L.; Plouet, J. Vascular endothelial growth factor is an autocrine growth factor for hair dermal papilla cells. J. Invest. Dermatol 1996, 106, 17–23. [Google Scholar]
- Kwack, M.H.; Kang, B.M.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Minoxidil activates beta-catenin pathway in human dermal papilla cells: A possible explanation for its anagen prolongation effect. J. Dermatol. Sci 2011, 62, 154–159. [Google Scholar]
- Itami, S.; Kurata, S.; Takayasu, S. Androgen induction of follicular epithelial cell growth is mediated via insulin-like growth factor-I from dermal papilla cells. Biochem. Biophys. Res. Commun 1995, 212, 988–994. [Google Scholar]
- Shimaoka, S.; Imai, R.; Ogawa, H. Dermal papilla cells express hepatocyte growth factor. J. Dermatol. Sci 1994, 7, S79–S83. [Google Scholar]
- Soma, T.; Dohrmann, C.E.; Hibino, T.; Raftery, L.A. Profile of transforming growth factor-beta responses during the murine hair cycle. J. Invest. Dermatol 2003, 121, 65–68. [Google Scholar]
- Soma, T.; Tsuji, Y.; Hibino, T. Involvement of transforming growth factor-beta2 in catagen induction during the human hair cycle. J. Invest. Dermatol 2002, 118, 993–997. [Google Scholar]
- Choi, J.G.; Kang, O.H.; Brice, O.O.; Lee, Y.S.; Chae, H.S.; Oh, Y.C.; Sohn, D.H.; Park, H.; Choi, H.G.; Kim, S.G.; et al. Antibacterial activity of Ecklonia cava against methicillin-resistant Staphylococcus aureus and Salmonella spp. Foodborne. Pathog. Dis 2010, 7, 435–441. [Google Scholar]
- Kim, T.H.; Bae, J.S. Ecklonia cava extracts inhibit lipopolysaccharide induced inflammatory responses in human endothelial cell. Food Chem. Toxicol 2010, 48, 1682–1687. [Google Scholar]
- Ahn, M.J.; Yoon, K.D.; Min, S.Y.; Lee, J.S.; Kim, J.H.; Kim, T.G.; Kim, S.H.; Kim, N.G.; Huh, H.; Kim, J. Inhibition of HIV-1 reverse transcriptase and protease by phlorotannins from the brown alga Ecklonia cava. Biol. Pharm. Bull 2004, 27, 544–547. [Google Scholar]
- Kang, K.A.; Lee, K.H.; Chae, S.; Koh, Y.S.; Yoo, B.S.; Kim, J.H.; Ham, Y.M.; Baik, J.S.; Lee, N.H.; Hyun, J.W. Triphlorethol-A from Ecklonia cava protects V79-4 lung fibroblast against hydrogen peroxide induced cell damage. Free Radic. Res 2005, 39, 883–892. [Google Scholar]
- Kang, K.A.; Lee, K.H.; Park, J.W.; Lee, N.H.; Na, H.K.; Surh, Y.J.; You, H.J.; Chung, M.H.; Hyun, J.W. Triphlorethol-A induces heme oxygenase-1 via activation of ERK and NF-E2 related factor 2 transcription factor. FEBS Lett 2007, 581, 2000–2008. [Google Scholar]
- Kong, C.S.; Kim, J.A.; Yoon, N.Y.; Kim, S.K. Induction of apoptosis by phloroglucinol derivative from Ecklonia cava in MCF-7 human breast cancer cells. Food Chem. Toxicol 2009, 47, 1653–1658. [Google Scholar]
- Park, E.; Ahn, G.N.; Lee, N.H.; Kim, J.M.; Yun, J.S.; Hyun, J.W.; Jeon, Y.J.; Wie, M.B.; Lee, Y.J.; Park, J.W.; Jee, Y. Radioprotective properties of eckol against ionizing radiation in mice. FEBS Lett 2008, 582, 925–930. [Google Scholar]
- Jung, W.K.; Heo, S.J.; Jeon, Y.J.; Lee, C.M.; Park, Y.M.; Byun, H.G.; Choi, Y.H.; Park, S.G.; Choi, I.W. Inhibitory effects and molecular mechanism of dieckol isolated from marine brown alga on COX-2 and iNOS in microglial cells. Agric. Food Chem 2009, 57, 4439–4446. [Google Scholar]
- Müller-Röver, S.; Handjiski, B.; van der Veen, C.; Maurer, M.; Eichmüller, S.; Ling, G.; Hofmann, U.; Foitzik, K.; Mecklenburg, L.; Handjiski, B. A comprehensive guide for the accurate classification of murine hair follicles in distinct hair cycle stages. J. Invest. Dermatol 2001, 117, 3–15. [Google Scholar]
- Ibrahim, L.; Wright, E.A. The growth of rats and mice vibrissae under normal and some abnormal conditions. J. Embryol. Exp. Morphol 1975, 33, 831–844. [Google Scholar]
- Philpott, M.P.; Kealey, T. Cyclical changes in rat vibrissa follicles maintained in vitro. J. Invest. Dermatol 2000, 115, 1152–1155. [Google Scholar]
- Philpott, M.P.; Green, M.R.; Kealey, T. Human hair growth in vitro. J. Cell Sci 1990, 97, 463–471. [Google Scholar]
- Botchkarev, V.A.; Kishimoto, J. Molecular control of epithelial-mesenchymal interactions during hair follicle cycling. J. Investig. Dermatol. Symp. Proc 2003, 8, 46–55. [Google Scholar]
- Paus, R.; Cotsarelis, G. The biology of hair follicles. N. Engl. J. Med 1999, 341, 491–497. [Google Scholar]
- Dubois, J.M.; Rouzaire-Dubois, B. Role of potassium channels in mitogenesis. Prog. Biophys. Mol. Biol 1993, 59, 1–21. [Google Scholar]
- Harmon, C.S.; Lutz, D.; Ducote, J. Potassium channel openers stimulate DNA synthesis in mouse epidermal keratinocyte and whole hair follicle cultures. Skin Pharmacol 1993, 6, 170–178. [Google Scholar]
- Sanders, D.A.; Fiddes, I.; Thompson, D.M.; Philpott, M.P.; Westgate, G.E.; Kealey, T. In the absence of streptomycin, minoxidil potentiates the mitogenic effects of fetal calf serum, insulin-like growth factor 1, and platelet-derived growth factor on NIH 3T3 fibroblasts in a K+ channel-dependent fashion. J. Invest. Dermatol 1996, 107, 229–234. [Google Scholar]
- Whiting, D.A. Male pattern hair loss: Current understanding. Int. J. Dermatol 1998, 37, 561–566. [Google Scholar]
- Trüeb, R.M. Molecular mechanisms of androgenetic alopecia. Exp. Gerontol 2002, 37, 981–990. [Google Scholar]
- Sinclair, R. Male pattern androgenetic alopecia. BMJ 1998, 317, 865–869. [Google Scholar]
- Hibino, T.; Nishiyama, T. Role of TGF-beta2 in the human hair cycle. J. Dermatol. Sci 2004, 35, 9–18. [Google Scholar]
- Inui, S.; Fukuzato, Y.; Nakajima, T.; Yoshikawa, K.; Itami, S. Androgen-inducible TGF-beta1 from balding dermal papilla cells inhibits epithelial cell growth: A clue to understand paradoxical effects of androgen on human hair growth. FASEB J 2002, 16, 1967–1969. [Google Scholar]
- Kwack, M.H.; Sung, Y.K.; Chung, E.J.; Im, S.U.; Ahn, J.S.; Kim, M.K.; Kim, J.C. Dihydrotestosterone-inducible dickkopf 1 from balding dermal papilla cells causes apoptosis in follicular keratinocytes. J. Invest. Dermatol 2008, 128, 262–269. [Google Scholar]
- Heo, S.J.; Jeon, Y.J.; Lee, J.; Kim, H.T.; Lee, K.W. Antioxidant effect of enzymatic hydrolyzates from a Kelp, Ecklonia cava. Algae 2003, 18, 341–347. [Google Scholar]
- Li, Y.; Qian, Z.J.; Ryu, B.M.; Lee, S.H.; Kim, M.N.; Kim, S.K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga, Ecklonia cava. Bioorg. Med. Chem 2009, 17, 1963–1973. [Google Scholar]
- Ohnemus, U.; Uenalan, M.; Conrad, F.; Handjiski, B.; Mecklenburg, L.; Nakamura, M.; Inzunza, J.; Gustafsson, J.A.; Paus, R. Hair cycle control by estrogens: Catagen induction via estrogen receptor (ER)-alpha is checked by ER beta signaling. Endocrinology 2005, 146, 1214–1225. [Google Scholar]
- Filsell, W.; Little, J.C.; Stones, A.J.; Granger, S.P.; Bayley, S.A. Transfection of rat dermal papilla cells with a gene encoding a temperature-sensitive polyomavirus large T antigen generates cell lines a differentiated phenotype. J. Cell Sci 1994, 107, 1761–1772. [Google Scholar]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res 1987, 47, 936–942. [Google Scholar]
- Hirosumi, J.; Nakayama, O.; Fagan, T.; Sawada, K.; Chida, N.; Inami, M.; Takahashi, S.; Notsu, Y.; Okuhara, M.J. FK143, a novel nonsteroidal inhibitor of steroid 5-reductase: (1) In vitro effects on human and animal prostatic enzymes. Steroid. Biochem. Mol. Biol 1995, 52, 357–363. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (μg/mL) | Compounds | |||
|---|---|---|---|---|
| Eckol | Dieckol | Phloroglucinol | Triphlorethol-A | |
| 0.005 | 100.8 ± 1.6 * | 100.5 ± 4.7 ** | 102.0 ± 8.5 | 101.7 ± 7.9 |
| 0.01 | 106.1 ± 4.5 * | 103.9 ± 6.5 | 100.4 ± 6.8 | 102.9 ± 3.5 |
| 0.05 | 120.3 ± 9.9 | 113.5 ± 6.2 * | 96.4 ± 5.3 | 100.4 ± 8.2 |
| 0.1 | 108.5 ± 7.7 | 106.1 ± 5.4 | 99.6 ± 4.7 | 99.3 ± 5.2 |
| 0.5 | 107.8 ± 6.7 | 108.1 ± 6.2 | 99.2 ± 4.0 | 99.4 ± 7.7 |
| 1 | 105.4 ± 6.2 | 98.5 ± 6.5 | 98.3 ± 2.3 | 103.3 ± 7.5 |
| 10 | 104.1 ± 9.7 | 97.8 ± 3.9 | 76.8 ± 5.6 | 100.6 ± 4.5 |
| 1 μM Minoxidil sulfate | 119.0 ± 8.1 *** | |||
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kang, J.-I.; Kim, S.-C.; Kim, M.-K.; Boo, H.-J.; Jeon, Y.-J.; Koh, Y.-S.; Yoo, E.-S.; Kang, S.-M.; Kang, H.-K. Effect of Dieckol, a Component of Ecklonia cava, on the Promotion of Hair Growth. Int. J. Mol. Sci. 2012, 13, 6407-6423. https://doi.org/10.3390/ijms13056407
Kang J-I, Kim S-C, Kim M-K, Boo H-J, Jeon Y-J, Koh Y-S, Yoo E-S, Kang S-M, Kang H-K. Effect of Dieckol, a Component of Ecklonia cava, on the Promotion of Hair Growth. International Journal of Molecular Sciences. 2012; 13(5):6407-6423. https://doi.org/10.3390/ijms13056407
Chicago/Turabian StyleKang, Jung-Il, Sang-Cheol Kim, Min-Kyoung Kim, Hye-Jin Boo, You-Jin Jeon, Young-Sang Koh, Eun-Sook Yoo, Sung-Myung Kang, and Hee-Kyoung Kang. 2012. "Effect of Dieckol, a Component of Ecklonia cava, on the Promotion of Hair Growth" International Journal of Molecular Sciences 13, no. 5: 6407-6423. https://doi.org/10.3390/ijms13056407
APA StyleKang, J.-I., Kim, S.-C., Kim, M.-K., Boo, H.-J., Jeon, Y.-J., Koh, Y.-S., Yoo, E.-S., Kang, S.-M., & Kang, H.-K. (2012). Effect of Dieckol, a Component of Ecklonia cava, on the Promotion of Hair Growth. International Journal of Molecular Sciences, 13(5), 6407-6423. https://doi.org/10.3390/ijms13056407