Ultraviolet C Irradiation Induces Different Expression of Cyclooxygenase 2 in NIH 3T3 Cells and A431 Cells: The Roles of COX-2 Are Different in Various Cell Lines

Abstract
:1. Introduction
2. Results and Discussion
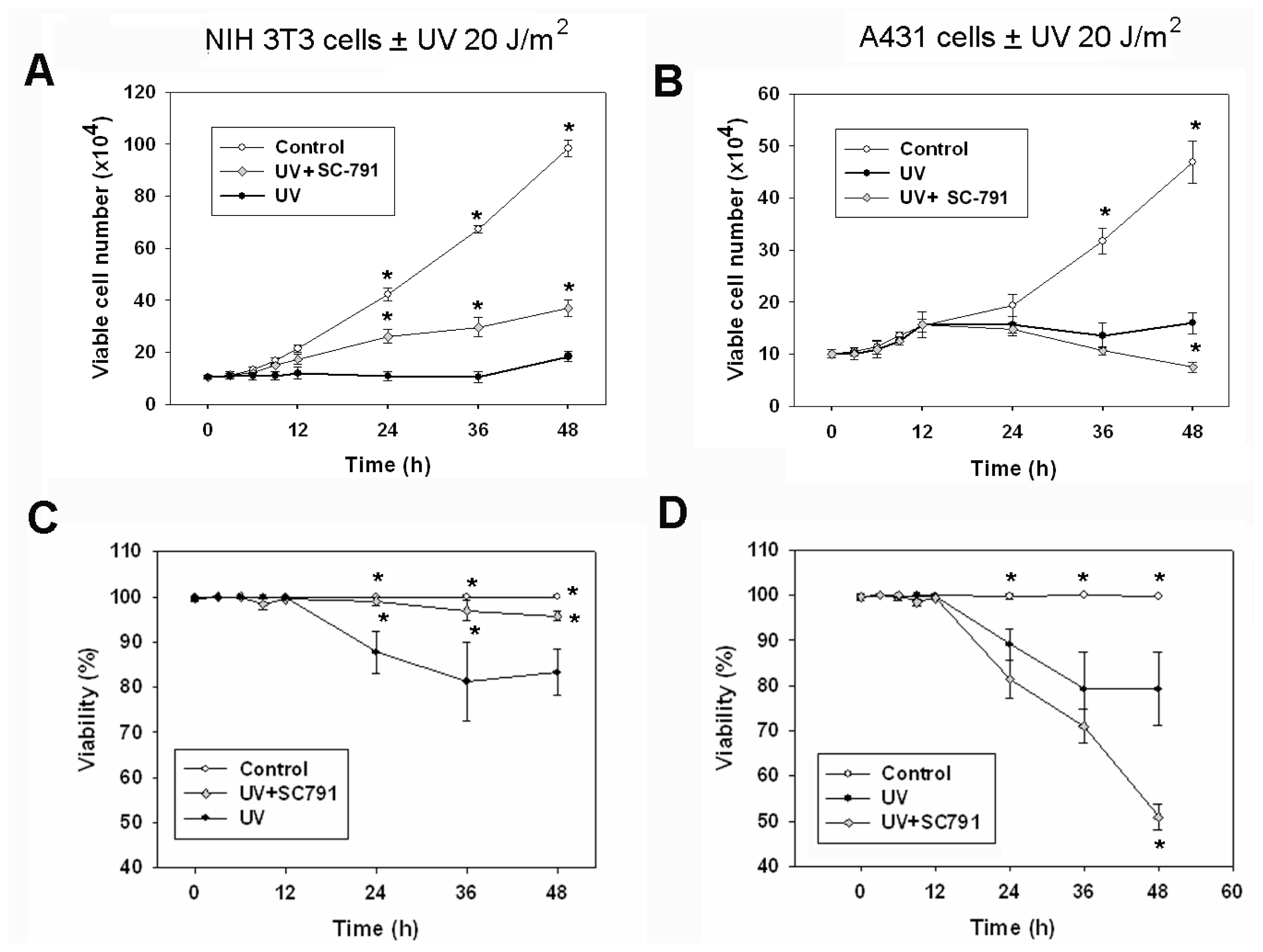
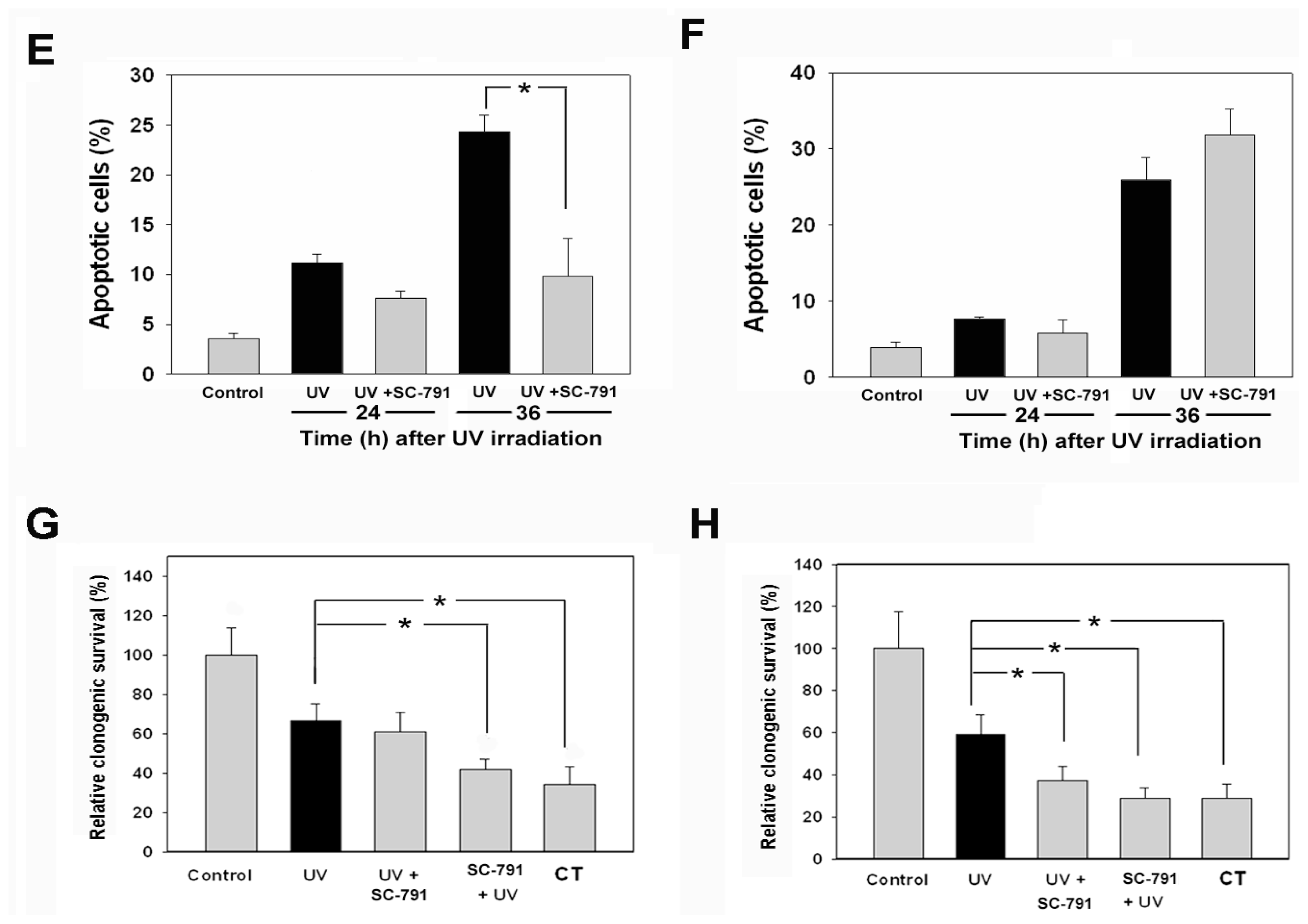
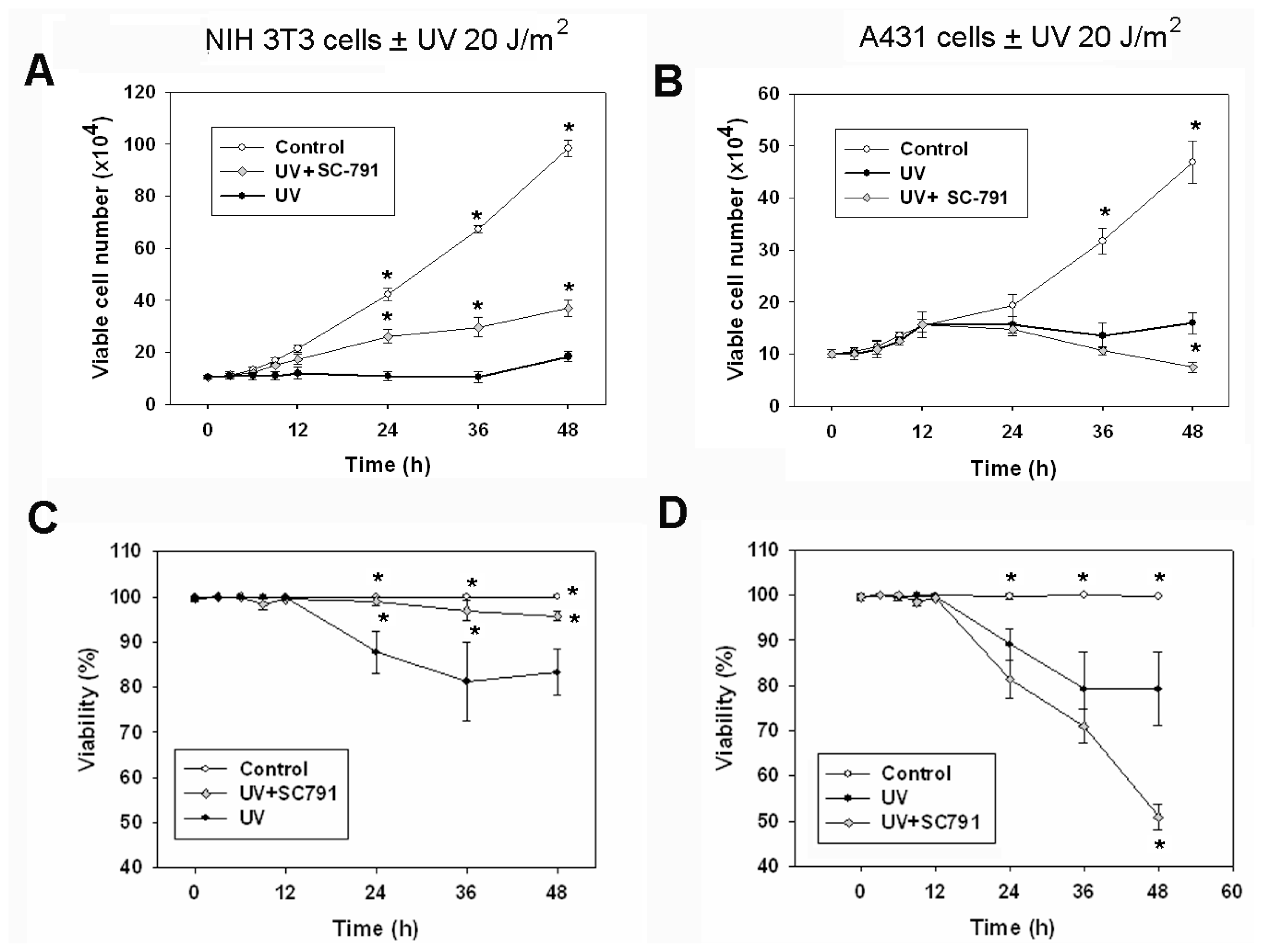
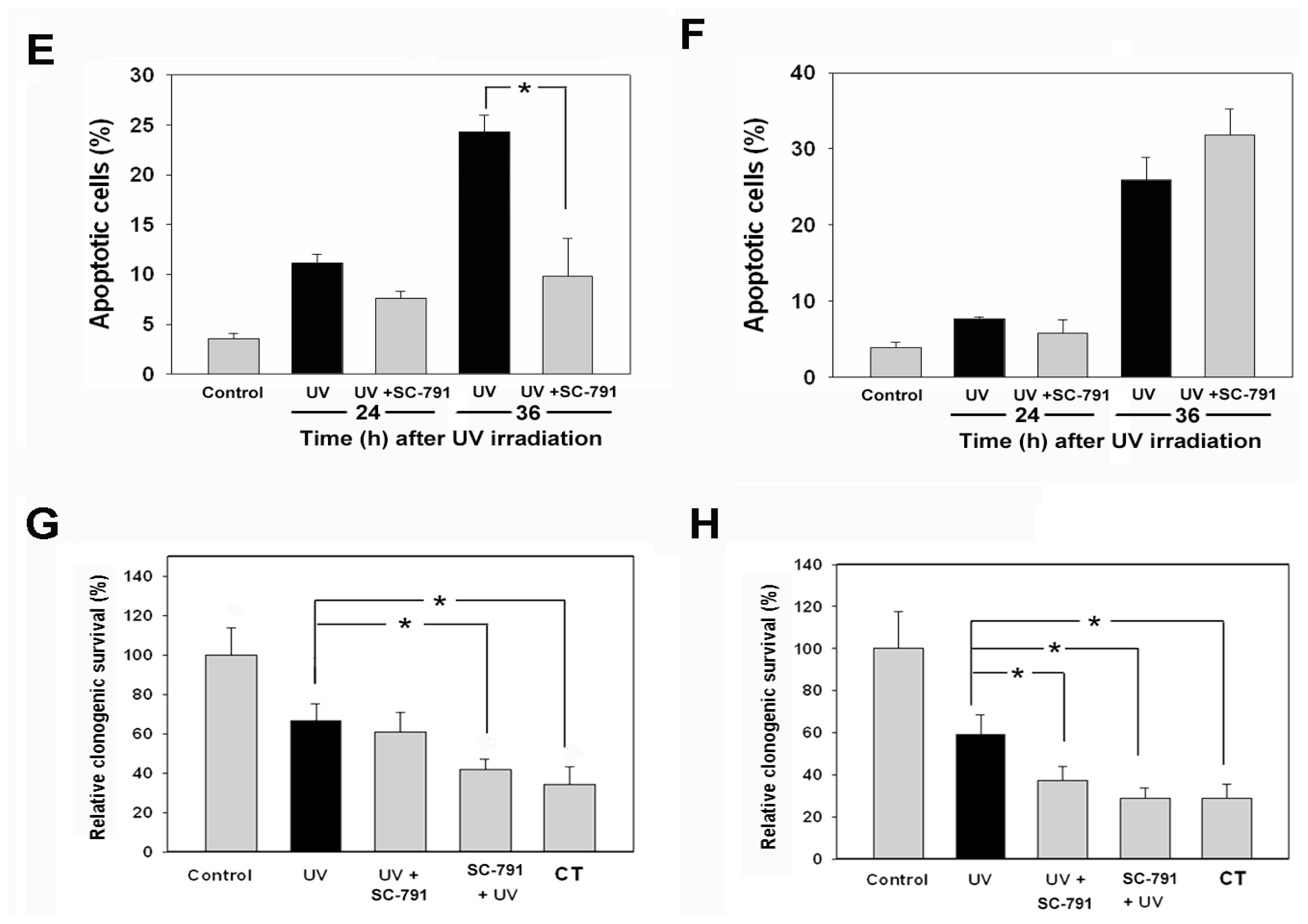
2.1. The COX-2 Inhibitor (SC-791) Attenuated UVC-Induced Apoptosis in NIH 3T3 Cells, but not in A431 Cells
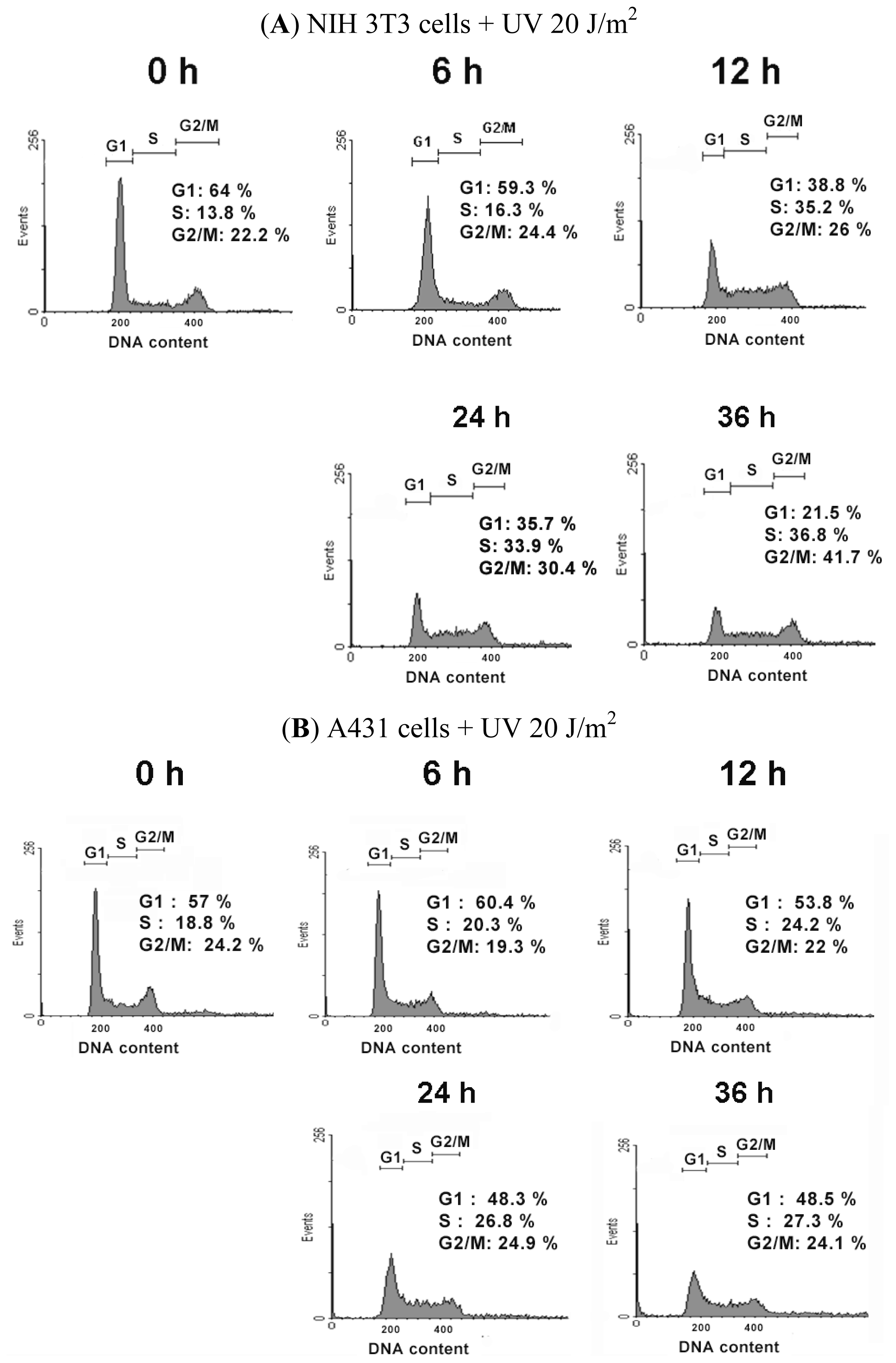
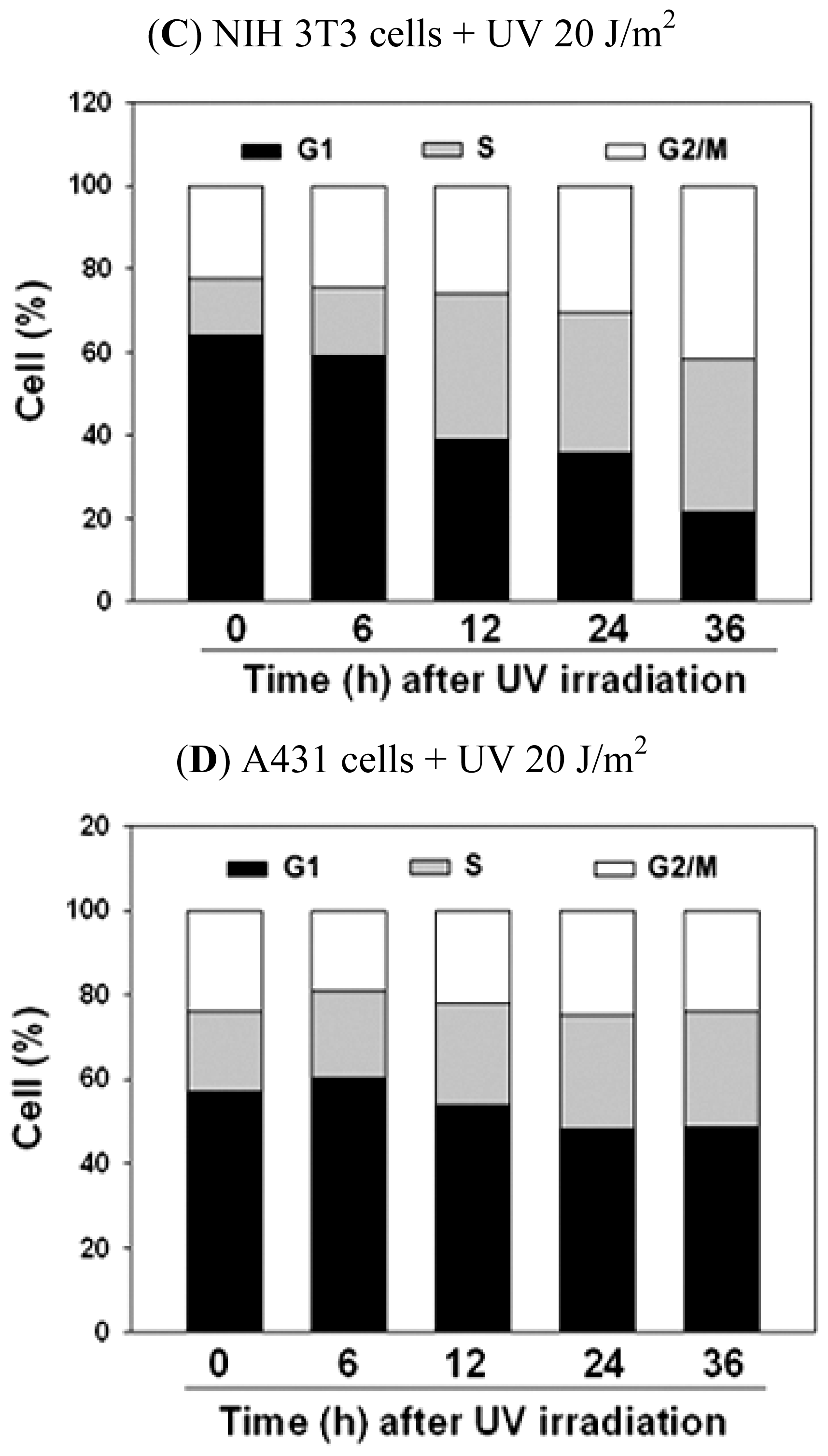
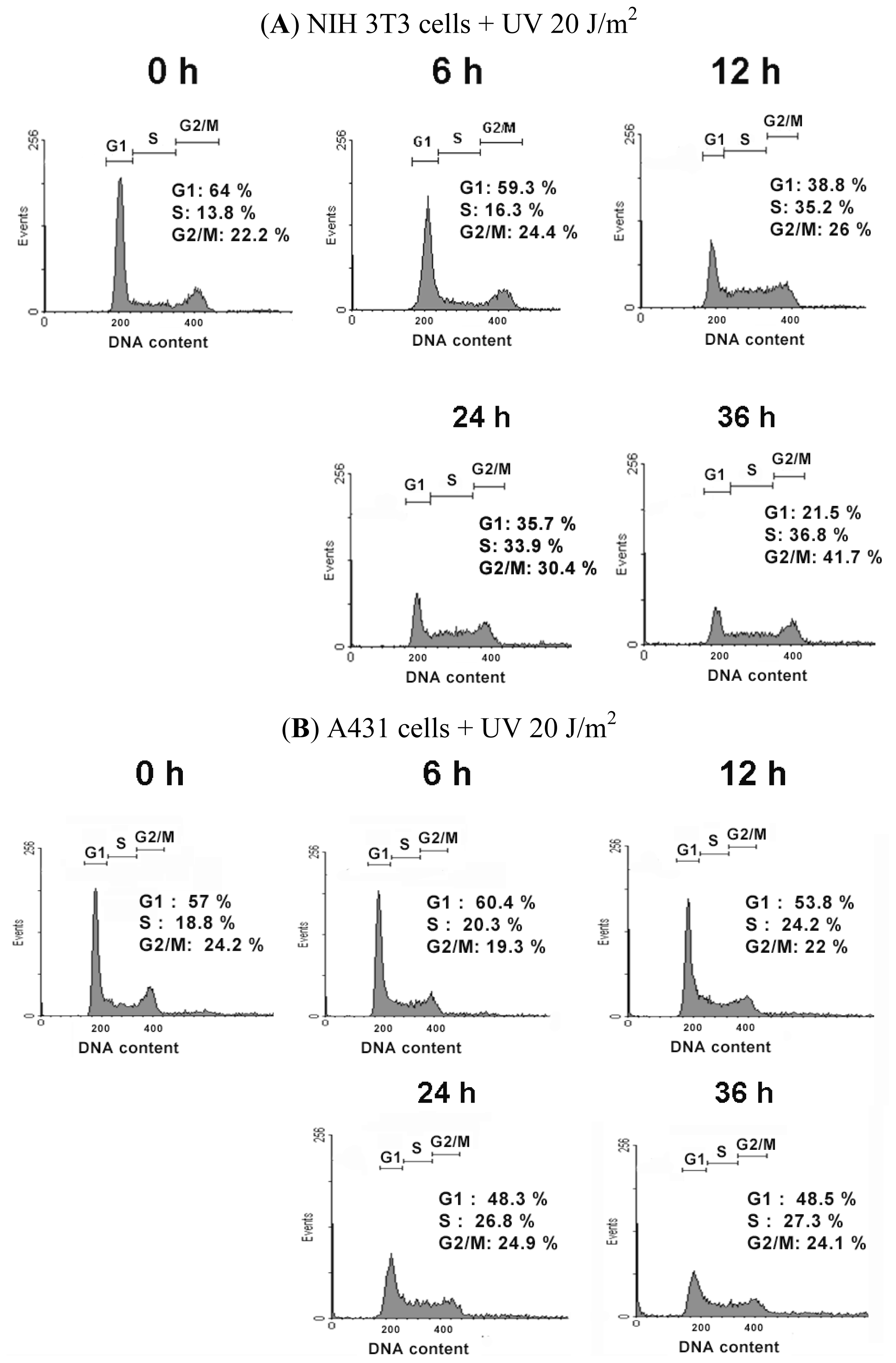
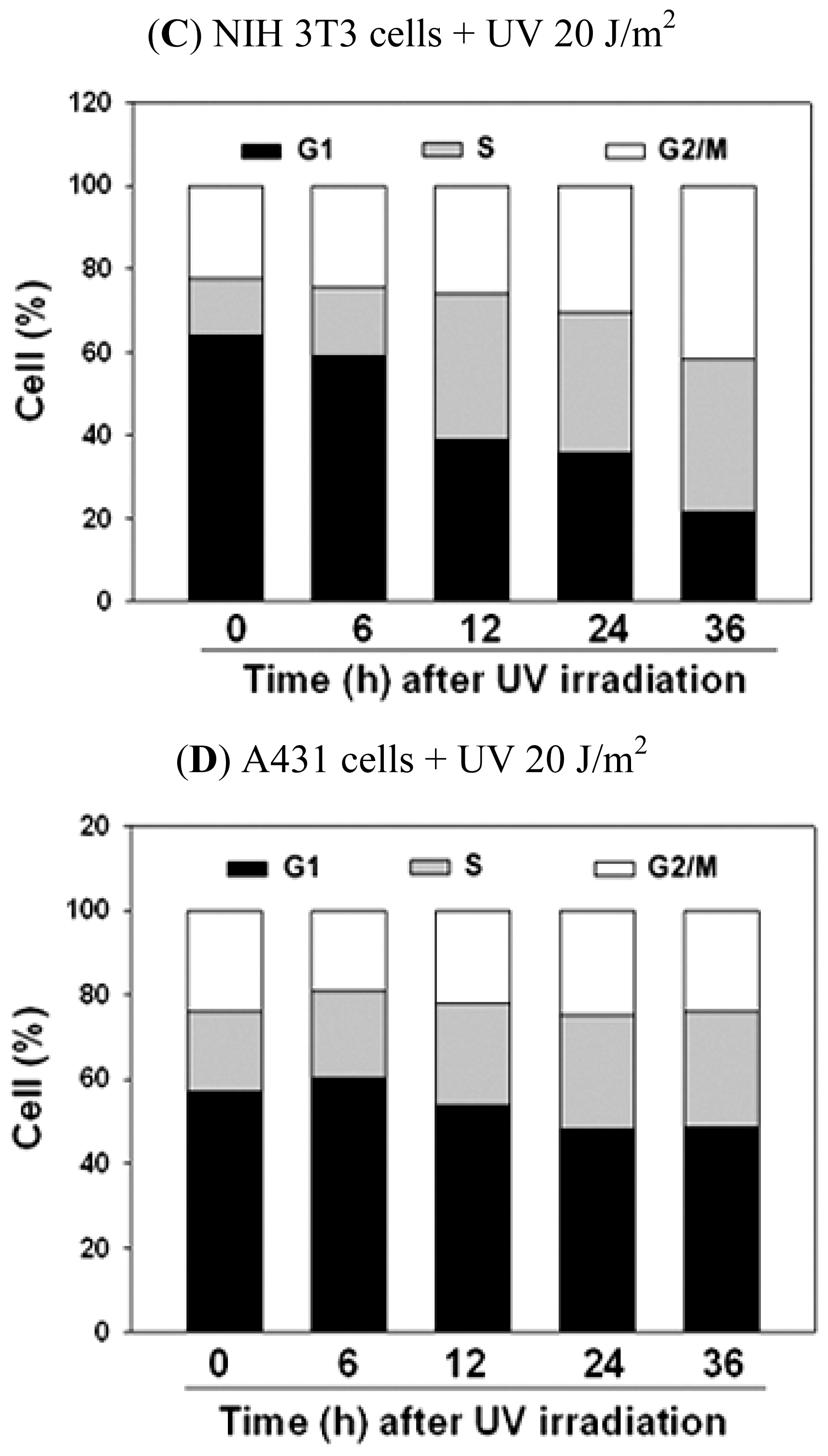
2.2. UVC-Induced Different Distributions of Cell Cycle in NIH 3T3 Cells and A431 Cells
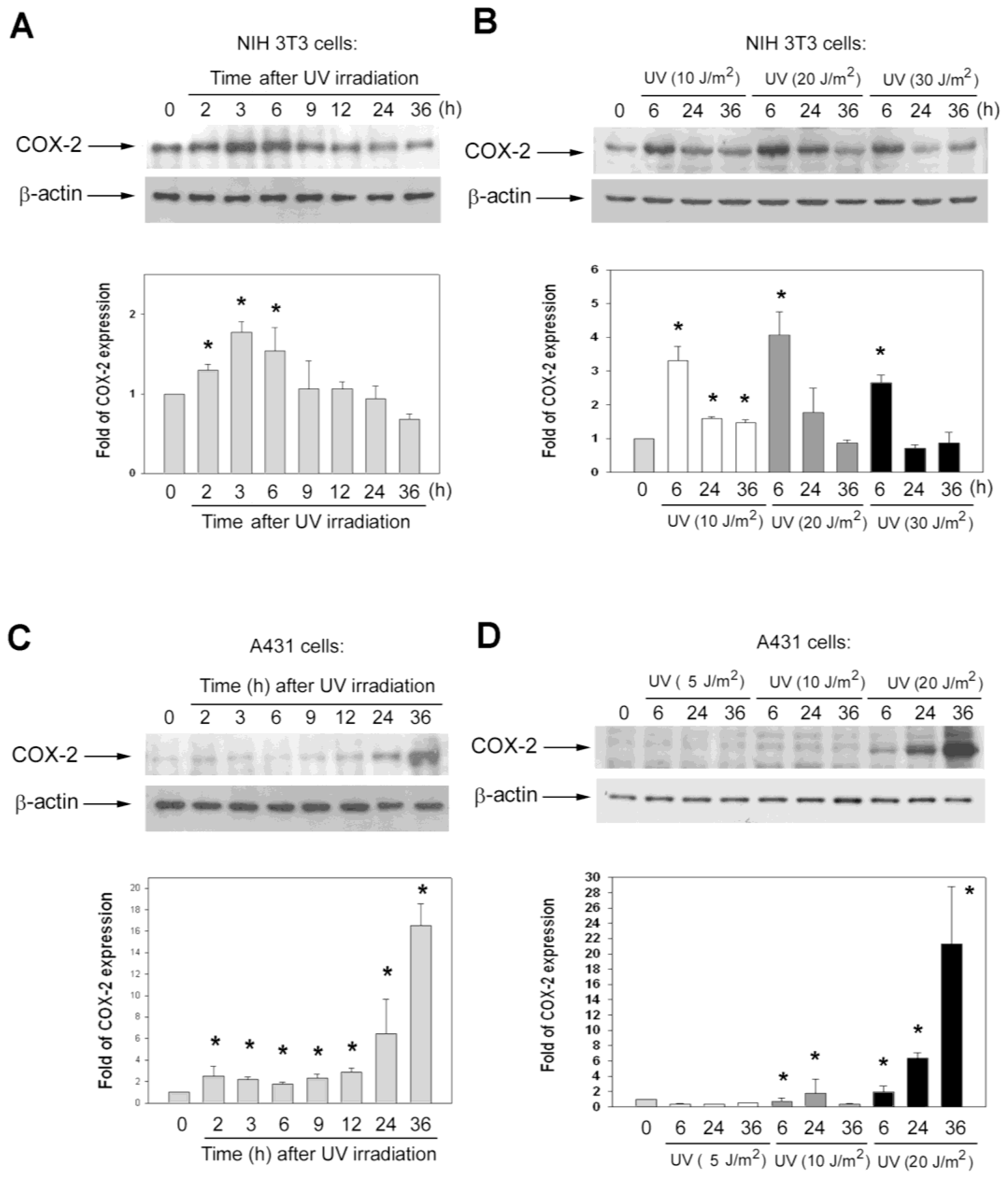
2.3. The Kinetics of COX-2 Protein Induction after UV Irradiation Were Different in Various Cell Lines
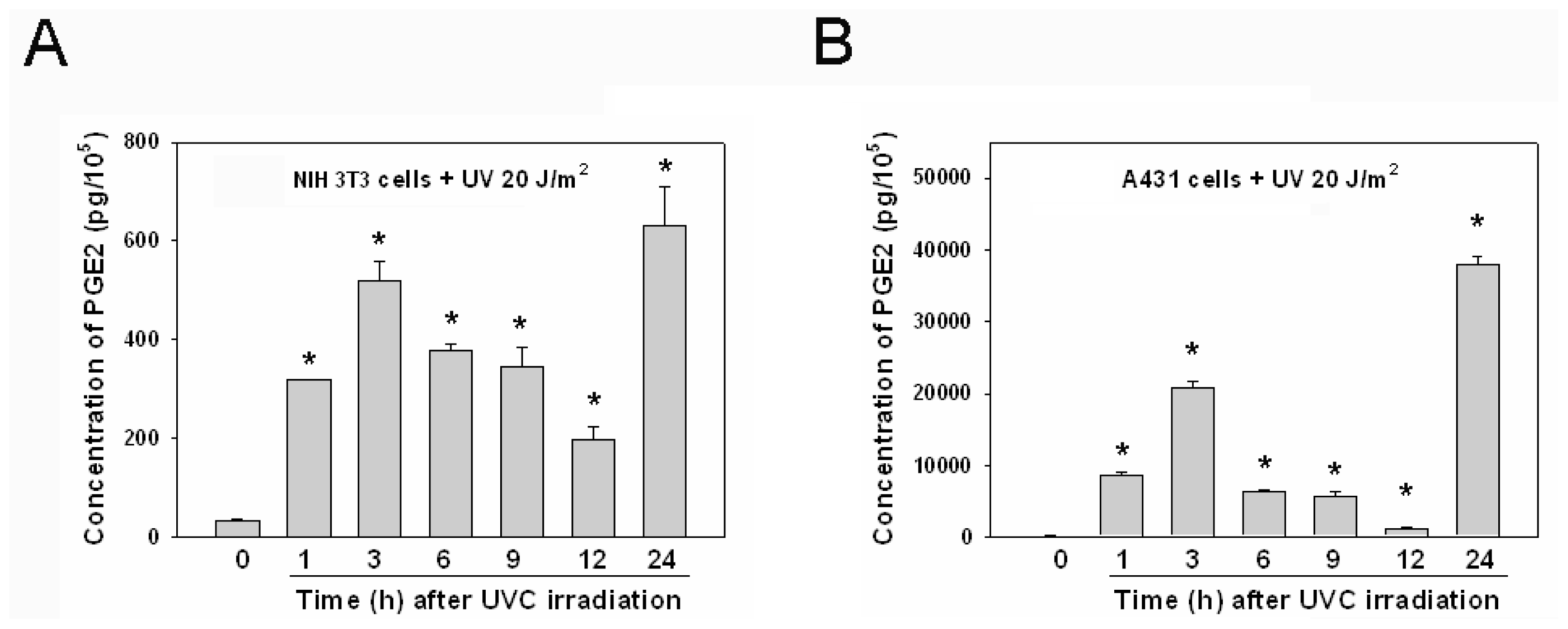
2.4. The Kinetics of PGE2 Induction after UVC Irradiation Was Similar in Various Cell Lines
2.5. Discussion
3. Experimental Section
3.1. Reagents and Antibodies
3.2. Cell Culture and UVC Treatments
3.3. Cell Growth Curve and Clonogenic Survival Assay
3.4. Immunoblotting
3.5. Prostaglandin E2 (PGE2) Measurement
3.6. Cell Cycle Analysis
3.7. Apoptotic Cell Analysis
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Tyrrell, R.M. Activation of mammalian gene expression by the UV component of sunlight—From models to reality. Bioessays 1996, 18, 139–148. [Google Scholar]
- Pourzand, C.; Tyrrell, R.M. Apoptosis, the role of oxidative stress and the example of solar UV radiation. Photochem. Photobiol 1999, 70, 380–390. [Google Scholar]
- Buckman, S.Y.; Gresham, A.; Hale, P.; Hruza, G.; Anast, J.; Masferrer, J.; Pentland, A.P. COX-2 expression is induced by UVB exposure in human skin: Implications for the development of skin cancer. Carcinogenesis 1998, 19, 723–729. [Google Scholar]
- Rundhaug, J.E.; Mikulec, C.; Pavone, A.; Fischer, S.M. A role for cyclooxygenase-2 in ultraviolet light-induced skin carcinogenesis. Mol. Carcinog 2007, 46, 692–698. [Google Scholar]
- Wu, M.H.; Chang, J.H.; Yung, B.Y. Resistance to UV-induced cell-killing in nucleophosmin/B23 over-expressed NIH 3T3 fibroblasts: Enhancement of DNA repair and up-regulation of PCNA in association with nucleophosmin/B23 over-expression. Carcinogenesis 2002, 23, 93–100. [Google Scholar]
- Wu, M.H.; Yung, B.Y. UV stimulation of nucleophosmin/B23 expression is an immediate-early gene response induced by damaged DNA. J. Biol. Chem 2002, 277, 48234–48240. [Google Scholar]
- Wu, M.H.; Chang, J.H.; Chou, C.C.; Yung, B.Y. Involvement of nucleophosmin/B23 in the response of HeLa cells to UV irradiation. Int. J. Cancer 2002, 97, 297–305. [Google Scholar]
- Simmons, D.L.; Botting, R.M.; Hla, T. Cyclooxygenase isozymes: The biology of prostaglandin synthesis and inhibition. Pharmacol. Rev 2004, 56, 387–437. [Google Scholar]
- Smith, W.L.; Dewitt, D.L. Prostaglandin endoperoxide H synthases-1 and -2. Adv. Immunol 1996, 62, 167–215. [Google Scholar]
- Rhodes, L.E.; Belgi, G.; Parslew, R.; McLoughlin, L.; Clough, G.F.; Friedmann, P.S. Ultraviolet-B-induced erythema is mediated by nitric oxide and prostaglandin E2 in combination. J. Invest. Dermatol 2001, 117, 880–885. [Google Scholar]
- Tong, X.; Van Dross, R.T.; Abu-Yousif, A.; Morrison, A.R.; Pelling, J.C. Apigenin prevents UVB-induced cyclooxygenase 2 expression: Coupled mRNA stabilization and translational inhibition. Mol. Cell. Biol 2007, 27, 283–296. [Google Scholar]
- Lallemand, D.; Ham, J.; Garbay, S.; Bakiri, L.; Traincard, F.; Jeannequin, O.; Pfarr, C.M.; Yaniv, M. Stress-activated protein kinases are negatively regulated by cell density. EMBO J 1998, 17, 5615–5626. [Google Scholar]
- Laszlo, C.F.; Fayad, S.; Carpenter, O.L.; George, K.S.; Lu, W.; Saad, A.A.; Wu, S. The role of translational regulation in ultraviolet C light-induced cyclooxygenase-2 expression. Life Sci 2009, 85, 70–76. [Google Scholar]
- Olsen, B.B.; Neves-Petersen, M.T.; Klitgaard, S.; Issinger, O.G.; Petersen, S.B. UV light blocks EGFR signalling in human cancer cell lines. Int. J. Oncol 2007, 30, 181–185. [Google Scholar]
- Ibuki, Y.; Goto, R. Antiapoptotic effects induced by different wavelengths of ultraviolet light. Photochem. Photobiol 2002, 75, 495–502. [Google Scholar]
- Nunez, G.; Benedict, M.A.; Hu, Y.; Inohara, N. Caspases: The proteases of the apoptotic pathway. Oncogene 1998, 17, 3237–3245. [Google Scholar]
- Takasawa, R.; Nakamura, H.; Mori, T.; Tanuma, S. Differential apoptotic pathways in human keratinocyte HaCaT cells exposed to UVB and UVC. Apoptosis 2005, 10, 1121–1130. [Google Scholar]
- Chun, K.S.; Akunda, J.K.; Langenbach, R. Cyclooxygenase-2 inhibits UVB-induced apoptosis in mouse skin by activating the prostaglandin E2 receptors, EP2 and EP4. Cancer Res 2007, 67, 2015–2021. [Google Scholar]
- Liou, J.Y.; Ellent, D.P.; Lee, S.; Goldsby, J.; Ko, B.S.; Matijevic, N.; Huang, J.C.; Wu, K.K. Cyclooxygenase-2-derived prostaglandin e2 protects mouse embryonic stem cells from apoptosis. Stem Cells 2007, 25, 1096–1103. [Google Scholar]
- Choi, K.H.; Hama-Inaba, H.; Wang, B.; Haginoya, K.; Odaka, T.; Yamada, T.; Hayata, I.; Ohyama, H. UVC-induced apoptosis in human epithelial tumor A431 cells: Sequence of apoptotic changes and involvement of caspase (-8 and -3) cascade. J. Radiat. Res 2000, 41, 243–258. [Google Scholar]
- Manke, I.A.; Nguyen, A.; Lim, D.; Stewart, M.Q.; Elia, A.E.; Yaffe, M.B. MAPKAP kinase-2 is a cell cycle checkpoint kinase that regulates the G2/M transition and S phase progression in response to UV irradiation. Mol. Cell 2005, 17, 37–48. [Google Scholar]
- Orren, D.K.; Petersen, L.N.; Bohr, V.A. Persistent DNA damage inhibits S-phase and G2 progression, and results in apoptosis. Mol. Biol. Cell 1997, 8, 1129–1142. [Google Scholar]
- Serini, S.; Donato, V.; Piccioni, E.; Trombino, S.; Monego, G.; Toesca, A.; Innocenti, I.; Missori, M.; De Spirito, M.; Celleno, L.; et al. Docosahexaenoic acid reverts resistance to UV-induced apoptosis in human keratinocytes: Involvement of COX-2 and HuR. J. Nutr. Biochem 2011, 22, 874–885. [Google Scholar]
- Judson, B.L.; Miyaki, A.; Kekatpure, V.D.; Du, B.; Gilleaudeau, P.; Sullivan-Whalen, M.; Mohebati, A.; Nair, S.; Boyle, J.O.; Granstein, R.D.; et al. UV radiation inhibits 15-hydroxyprostaglandin dehydrogenase levels in human skin: Evidence of transcriptional suppression. Cancer Prev. Res. (Phila) 2010, 3, 1104–1111. [Google Scholar]
- Wang, X.; Zhu, J.; Zhao, P.; Jiao, Y.; Xu, N.; Grabinski, T.; Liu, C.; Miranti, C.K.; Fu, T.; Cao, B.B. In vitro efficacy of immuno-chemotherapy with anti-EGFR human Fab-Taxol conjugate on A431 epidermoid carcinoma cells. Cancer Biol. Ther 2007, 6, 980–987. [Google Scholar]
- Williams, C.S.; Mann, M.; DuBois, R.N. The role of cyclooxygenases in inflammation, cancer, and development. Oncogene 1999, 18, 7908–7916. [Google Scholar]
- Xu, Z.; Choudhary, S.; Voznesensky, O.; Mehrotra, M.; Woodard, M.; Hansen, M.; Herschman, H.; Pilbeam, C. Overexpression of COX-2 in human osteosarcoma cells decreases proliferation and increases apoptosis. Cancer Res 2006, 66, 6657–6664. [Google Scholar]
- Zahner, G.; Wolf, G.; Ayoub, M.; Reinking, R.; Panzer, U.; Shankland, S.J.; Stahl, R.A. Cyclooxygenase-2 overexpression inhibits platelet-derived growth factor-induced mesangial cell proliferation through induction of the tumor suppressor gene p53 and the cyclin-dependent kinase inhibitors p21waf-1/cip-1 and p27kip-1. J. Biol. Chem 2002, 277, 9763–9771. [Google Scholar]
- Ziegler, A.; Jonason, A.S.; Leffell, D.J.; Simon, J.A.; Sharma, H.W.; Kimmelman, J.; Remington, L.; Jacks, T.; Brash, D.E. Sunburn and p53 in the onset of skin cancer. Nature 1994, 372, 773–776. [Google Scholar]
- Young, A.R. Acute effects of UVR on human eyes and skin. Prog. Biophys. Mol. Biol 2006, 92, 80–85. [Google Scholar]
- Learn, D.B.; Beard, J.; Moloney, S.J. The ultraviolet C energy emitted from FS lamps contributes significantly to the induction of human erythema and murine ear edema. Photodermatol. Photoimmunol. Photomed 1993, 9, 147–153. [Google Scholar]
- Jeong, H.S.; Kim, J.H.; Choi, H.Y.; Lee, E.R.; Cho, S.G. Induction of cell growth arrest and apoptotic cell death in human breast cancer MCF-7 cells by the COX-1 inhibitor FR122047. Oncol. Rep 2010, 24, 351–356. [Google Scholar]
- Hervouet, E.; Vallette, F.M.; Cartron, P.F. Dnmt1/Transcription factor interactions: An alternative mechanism of DNA methylation inheritance. Genes Cancer 2010, 1, 434–443. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The Cell Cycle Distribution after UV Irradiation in NIH 3T3 Cells | |||||
|---|---|---|---|---|---|
| Time | 0 h | 6 h | 12 h | 24 h | 36 h |
| Cell Cycle | |||||
| % of G1 phase | 63.50 ± 0.38 | 59.37 ± 1.54 | 38.86 ± 0.66 * | 36.37 ± 0.60 * | 28.95 ± 6.50 * |
| % of S phase | 15.41 ± 2.0 | 17.00 ± 1.08 | 35.99 ± 0.60 * | 37.36 ± 3.02 * | 37.68 ± 1.08 * |
| % of G2/M phase | 21.08 ± 1.77 | 23.62 ± 0.94 | 25.14 ± 1.16 * | 26.01 ± 3.14 | 33.36 ± 7.37 * |
| The Cell Cycle Distribution after UV Irradiation in A431 Cells | |||||
| Time | 0 h | 6 h | 12 h | 24 h | 36 h |
| Cell Cycle | |||||
| % of G1 phase | 53.05 ± 3.47 | 60.13 ± 0.51 * | 56.84 ± 2.66 | 53.22 ± 4.31 | 51.09 ± 2.25 |
| % of S phase | 22.93 ± 3.61 | 22.35 ± 1.91 | 24.64 ± 0.40 | 27.68 ± 0.82* | 29.40 ± 1.80 * |
| % of G2/M phase | 24.02 ± 0.20 | 17.52 ± 1.56 | 18.55 ± 3.04 | 19.10 ± 4.98 | 19.52 ± 4.02 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tai, M.-H.; Weng, C.-H.; Mon, D.-P.; Hu, C.-Y.; Wu, M.-H. Ultraviolet C Irradiation Induces Different Expression of Cyclooxygenase 2 in NIH 3T3 Cells and A431 Cells: The Roles of COX-2 Are Different in Various Cell Lines. Int. J. Mol. Sci. 2012, 13, 4351-4366. https://doi.org/10.3390/ijms13044351
Tai M-H, Weng C-H, Mon D-P, Hu C-Y, Wu M-H. Ultraviolet C Irradiation Induces Different Expression of Cyclooxygenase 2 in NIH 3T3 Cells and A431 Cells: The Roles of COX-2 Are Different in Various Cell Lines. International Journal of Molecular Sciences. 2012; 13(4):4351-4366. https://doi.org/10.3390/ijms13044351
Chicago/Turabian StyleTai, Ming-Hong, Chien-Hui Weng, Dir-Pu Mon, Chun-Yi Hu, and Ming-Hsiu Wu. 2012. "Ultraviolet C Irradiation Induces Different Expression of Cyclooxygenase 2 in NIH 3T3 Cells and A431 Cells: The Roles of COX-2 Are Different in Various Cell Lines" International Journal of Molecular Sciences 13, no. 4: 4351-4366. https://doi.org/10.3390/ijms13044351