Sequence Variants and Haplotype Analysis of Cat ERBB2 Gene: A Survey on Spontaneous Cat Mammary Neoplastic and Non-Neoplastic Lesions

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Extraction of Genomic DNA from FFPET and Frozen Samples
2.2. Analysis of Cat ERBB2 Gene DNA Fragments from Exons 17 to 20: Structural Features
2.3. Sequence Variant Detection in Cat-ERBB2_17-18 DNA Sequences
2.4. Sequence Variants Detection in the Cat ERBB2_19-20 DNA Sequences
2.5. Cat ERBB2_19-20 Haplotype Determination in Normal and Mammary Lesion Populations
2.6. Statistical Analysis of Cat Mammary Lesions, Clinicopathological Classifiers and Its Association with Haplotypes
2.7. Discussion
3. Experimental Section
3.1. Biological Material
3.2. Genomic DNA Extraction
3.3. PCR Amplification of Two Fragments from Cat ERBB2 Gene Comprising Exons 17 to 20
3.4. Sequencing and Analysis of DNA Fragments
3.5. Bioinformatic Splice Prediction Based on the SVs Detected
3.6. Statistic Analysis in Normal and Mammary Lesion Cat Populations. Testing Through DNA Variant Sequences and Clinicopathological Features
4. Conclusions
Supplementary Information
ijms-13-02783-s001.docAcknowledgments
References
- MacEwen, E.G. Spontaneous tumors in dogs and cats: Models for the study of cancer biology and treatment. Cancer Metastasis Rev 1990, 9, 125–136. [Google Scholar]
- Vail, D.M.; MacEwen, E.G. Spontaneously occurring tumors of companion animals as models for human cancer. Cancer Invest 2000, 18, 781–792. [Google Scholar]
- Zappulli, V.; de Zan, G.; Cardazzo, B.; Bargelloni, L.; Castagnaro, M. Feline mammary tumors in comparative oncology. J. Dairy Res 2005, 72, 98–106. [Google Scholar]
- Weijer, K.; Hart, A.A.M. Prognostic Factors in feline mammary carcinoma. J. Natl. Cancer Inst 1983, 70, 709–716. [Google Scholar]
- Hahn, K.A.; Bravo, L.; Avenell, J.S. Feline breast carcinoma as a pathologic and therapeutic model for human breast cancer. In Vivo 1994, 8, 825–828. [Google Scholar]
- de Maria, R.; Olivero, M.; Iussich, S.; Nakaichi, M.; Murata, T.; Biolatti, B.; di Renzo, M.F. Spontaneous feline mammary carcinoma is a model of HER2 overexpressing poor prognosis human breast cancer. Cancer Res 2005, 65, 907–912. [Google Scholar]
- Burrai, G.P.; Mohammed, S.I.; Miller, M.A.; Marras, V.; Pirino, S.; Addis, M.F.; Uzzau, S.; Antuofermo, E. Spontaneous feline mammary intraepithelial lesions as a model for human estrogen receptor- and progesterone receptor-negative breast lesions. BMC Cancer 2010, 10. [Google Scholar] [CrossRef] [Green Version]
- Lana, S.E.; Rutteman, G.R.; Withrow, S.J. Tumors of the Mammary Gland. In Small Animal Clinical Oncology, 4th ed; Withrow, S.J., Vail, D.M., Eds.; Saunders Elsevier: Toronto, Canada, 2001; pp. 628–636. [Google Scholar]
- Gimenez, F.; Hecht, S.; Craig, L.E.; Legendre, A.M. Early detection, aggressive therapy. Optimizing the management of feline mammary masses. J. Feline Med. Surg 2010, 12, 214–224. [Google Scholar]
- Owen, L.N. Classification of Tumors in Domestic Animals, 1st ed; World Health Organization: Geneva, Belgium, 1980. [Google Scholar]
- Ito, T.; Kadosawa, T.; Mochizuki, M.; Matsunaga, S.; Nishimura, R.; Sasaki, N. Prognosis of malignant mammary tumors in 53 cats. J. Vet. Med. Sci 1996, 58, 723–726. [Google Scholar]
- Winston, J.; Craft, D.M.; Scase, T.J.; Bergman, P.J. Immunohistochemical detection of erbB-2/neu expression in spontaneous feline mammary tumors. Vet. Comp. Oncol 2005, 3, 8–15. [Google Scholar]
- Ross, J.S.; Slodkowska, E.A.; Symmans, W.F.; Pusztai, L.; Ravnin, P.M.; Hortobagyi, G.N. The erbB-2 receptor and breast cancer: Ten years of targeted anti-erbB-2 therapy and personalized medicine. Oncologist 2009, 14, 320–368. [Google Scholar]
- Tao, W.; Wang, C.; Han, R.; Jiang, H. HER2 codon 655 polymorphism and breast cancer risk: A meta-analysis. Breast Cancer Res. Treat 2009, 114, 371–376. [Google Scholar]
- Penuel, E.; Schaefer, G.; Akita, R.W.; Sliwkowski, M.X. Structural requirements for ERBB2 transactivation. Semin. Oncol 2001, 28, 36–42. [Google Scholar]
- Benusiglio, P.R. Focus on ERBB2. Pharmaogenomics 2007, 9, 825–828. [Google Scholar]
- Slamon, D.J.; Godolphin, W.; Jones, L.A.; Holt, J.A.; Wong, S.G.; Keith, D.E.; Levin, W.J.; Stuart, S.G.; Udove, J.; Ullrich, A.; et al. Studies of the erbB-2/neu proto-oncogene in human breast and ovarian cancer. Science 1989, 244, 707–712. [Google Scholar]
- Kolibaba, K.S.; Druker, B.J. Protein tyrosine kinases and cancer. Biochim. Biophys. Acta 1997, 1333, 217–248. [Google Scholar]
- Ordás, J.; Millán, Y.; Dios, R.; Reymundo, C.; de las Mulas, J.M. Proto-oncogene erbB-2 in normal, dysplastic and tumorous feline mammary glands: An immunohistochemical and chromogenic in situ hybridization study. BMC Cancer 2007, 7. [Google Scholar] [CrossRef]
- Blume-Jensen, P.; Hunter, T. Oncogenic kinase signaling. Nature 2001, 411, 355–365. [Google Scholar]
- Borg, A.; Tandon, A.K.; Sigurdsson, H.; Clark, G.M.; Fernö, M.; Fuqua, S.A.W.; Killander, D.; McGuire, W.L. erbB-2/neu amplification predicts poor survival in node-positive breast cancer. Cancer Res 1990, 50, 4332–4337. [Google Scholar]
- Charpin, C.; Garcia, S.; Bouvier, C.; Martini, F.; Lavaut, M.N.; Allasia, C.; Bonnier, P.; Andrac, L. c-erbB-2 oncoprotein detected by automated quantitative immunocytochemistry in breast carcinomas correlates with patients’ overall and disease-free survival. Br. J. Cancer 1997, 75, 1667–1673. [Google Scholar]
- Ding, L.; Getz, G.; Wheeler, D.A.; Mardis, E.R.; McLellan, M.D.; Cibulskis, K.; Sougnez, C.; Greulich, H.; Muzny, D.M.; Morgan, M.B.; et al. Somatic mutations affect key pathways in lung adenocarcinoma. Nature 2008, 455, 1069–1075. [Google Scholar]
- Leary, R.J.; Lin, J.C.; Cummins, J.; Boca, S.; Wood, L.D.; Parsons, D.W.; Jones, S.; Sjöblom, T.; Park, B.H.; Parsons, R.; et al. Integrated analysis of homozygous deletions, focal amplifications, and sequence alterations in breast and colorectal cancers. Proc. Natl. Acad. Sci. USA 2008, 105, 16224–16229. [Google Scholar]
- Benusiglio, P.R.; Lesueur, F.; Luccarini, C.; Conroy, D.M.; Shah, M.; Easton, D.F.; Day, N.E.; Dunning, A.M.; Pharoah, P.D.; Ponder, B.A. Common ERBB2 polymorphisms and risk of breast cancer in a white British population: A case-control study. Breast Cancer Res 2005, 7, 204–209. [Google Scholar]
- Han, W.; Kang, D.; Lee, J.E.; Park, I.A.; Choi, J.Y.; Lee, K.M.; Bae, J.Y.; Kim, S.; Shin, E.S.; Lee, J.E.; et al. A haplotype analysis of erbB-2 gene polymorphisms: Association with breast cancer risk, erbB-2 protein expression in the tumor, and disease recurrence in Korea. Clin. Cancer Res 2005, 11, 4775–4778. [Google Scholar]
- Breyer, J.P.; Sanders, M.E.; Airey, D.C.; Cai, Q.; Yaspan, B.L.; Schuyler, P.A.; Dai, Q.; Boulos, F.; Olivares, M.G.; Bradley, K.M.; et al. Heritable variation of ERBB2 and breast cancer risk. Cancer Epidemiol. Biomarkers Prev 2009, 18, 1252–1258. [Google Scholar]
- Desmet, F.O.; Hamroun, D.; Lalande, M.; Collod-Béroud, G.; Claustres, M.; Béroud, C. Human splicing finder: An online bioinformatics tool to predict splicing signals. Nucleic Acids Res 2009, 37. [Google Scholar] [CrossRef]
- Mayr, B.; Schaffner, G.; Kurzbauer, R.; Reifinger, M.; Schellander, K. Sequence of an exon of tumor suppressor p53 gene—A comparative study in domestic animals: Mutation in a feline solid mammary carcinoma. Br. Vet. J 1995, 151, 325–329. [Google Scholar]
- Mayr, B.; Reifinger, M.; Alton, K.; Schaffner, G. Novel p53 tumor suppressor mutations in cases of spindle cell sarcoma, pleomorphic sarcoma and fibrosarcoma in cats. Vet. Res. Commun 1998, 22, 249–255. [Google Scholar]
- Mayr, B.; Blauensteiner, J.; Edlinger, A.; Reifinger, M.; Alton, K.; Schaffner, G.; Brem, G. Presence of p53 mutations in feline neoplasms. Res. Vet. Sci 2000, 68, 63–70. [Google Scholar]
- Baptista, C.S.; Santos, S.; Laso, A.; Bastos, E.; Ávila, S.; Guedes-Pinto, H.; Gärtner, F.; Gut, I.G.; Castrillo, J.L.; Chaves, R. Sequence variation and mRNA expression of the TWIST1 gene in cats with mammary hyperplasia and neoplasia. Vet. J 2011, 191, 203–207. [Google Scholar]
- Nelson, S.E.; Gould, M.N.; Hampton, J.M. A case-control study of the HER2 Ile655Val polymorphism in relation to risk of invasive breast cancer. Breast Cancer Res 2005, 7, R357–R364. [Google Scholar]
- Beauclair, S.; Formento, P.; Fischel, J.L.; Lescaut, W.; Largillier, R.; Chamorey, E.; Hofman, P.; Ferrero, J.M.; Pagès, G.; Milano, G. Role of the HER2 [Ile655Val] genetic polymorphism in tumorogenesis and in the risk of trastuzumab-related cardiotoxicity. Ann. Oncol 2007, 18, 1335–1341. [Google Scholar]
- Puputti, M.; Sihto, H.; Isola, J.; Butzow, R.; Joensuu, H.; Nupponen, N.N. Allelic imbalance of HER2 variant in sporadic breast and ovarian cancer. Cancer Genet. Cytogenet 2006, 167, 32–38. [Google Scholar]
- Jo, U.H.; Han, S.G.L.; Seo, J.H.; Park, K.H.; Lee, J.W.; Lee, H.J.; Ryu, J.S.; Kim, Y.H. The genetic polymorphisms of HER-2 and the risk of lung cancer in a Korean population. BMC Cancer 2008, 8. [Google Scholar] [CrossRef]
- Fleishman, S.J.; Schlessinger, J.; Ben-Tal, N. A putative molecular-activation switch in the transmembrane domain of ERBB2. Proc. Natl. Acad. Sci. USA 2002, 99, 15937–15940. [Google Scholar]
- Bennasroune, A.; Fickova, M.; Gardin, A.; Dirrig-Grosch, S.; Aunis, D.; Cremel, G.; Hubert, P. Transmembrane peptides as inhibitors of ErbB receptor signaling. Mol. Biol. Cell 2004, 15, 3464–3474. [Google Scholar]
- Lengauer, C.; Kinzler, K.W.; Vogelstein, B. Genetic instabilities in human cancers. Nature 1998, 396, 643–649. [Google Scholar]
- Hartwell, L. Defects in a cell cycle checkpoint may be responsible for the genomic instability of cancer cells. Cell 1992, 71, 543–546. [Google Scholar]
- Bauer-Mehren, A.; Furlong, L.I.; Rautschka, M.; Sanz, F. From SNPs to pathways: Integration of functional effect of sequence variations on models of cell signaling pathways. BMC Bioinform 2009, 10. [Google Scholar] [CrossRef]
- Venables, J.P. Aberrant and alternative splicing in cancer. Cancer Res 2004, 64, 7647–7654. [Google Scholar]
- Loretti, A.P.; Ilha, M.R.; Ordas, J.; Martin de las Mulas, J. Clinical, pathological and immunohistochemical study of feline mammary fibro epithelial hyperplasia following a single injection of depot medroxyprogesterone acetate. J. Feline Med. Surg 2005, 7, 43–52. [Google Scholar]
- Rutteman, G.R.; Misdorp, W. Hormonal background of canine and feline mammary tumors. J. Reprod. Fertil. Suppl 1993, 47, 483–487. [Google Scholar]
- Hahn, K.A.; Bravo, L.; Avenell, J.S. Feline breast carcinoma as a pathologic and therapeutic model for human breast cancer. In Vivo 1994, 8, 825–828. [Google Scholar]
- Hampe, J.F.; Misdorp, W. Tumours and dysplasias of the mammary gland. Bull. World Health Organ 1974, 50, 111–133. [Google Scholar]
- MacEwen, E.G.; Hayes, A.A.; Harvey, H.J.; Patnaik, A.K.; Mooney, S.; Passe, S. Prognostic factors for feline mammary tumors. J. Am. Vet. Med. Assoc 1984, 2, 201–204. [Google Scholar]
- Viste, J.R.; Myers, S.L.; Singh, B.; Simko, E. Feline mammary adenocarcinoma: Tumor size as a prognostic indicator. Can. Vet. J 2002, 43, 33–37. [Google Scholar]
- Misdorp, W.; Else, R.W.; Helmén, E.; Lipscomb, T.P. Histological Classification of Mammary Tumors of the Dog and Cat; Armed Forces Institute of Pathology and World Health Organization: Washington, DC, USA, 1999. [Google Scholar]
- Santos, S.; Sá, D.; Bastos, E.; Guedes-Pinto, H.; Gut, I.; Gärtner, F.; Chaves, R. An efficient protocol for genomic DNA extraction from formalin-fixed paraffin-embedded tissues. Res. Vet. Sci 2009, 86, 421–426. [Google Scholar]
- den Dunnen, J.T.; Antonarakis, S.E. Mutation nomenclature extensions and suggestions to describe complex mutations: A Discussion. Hum. Mutat 2000, 15, 7–12. [Google Scholar]
- den Dunnen, J.T.; Antonarakis, S.E. Nomenclature for the description of human sequence variations. Hum Genet 2001, 109, 121–124. [Google Scholar]
- Ogino, S.; Gulley, M.L.; den Dunnen, J.T.; Wilson, R.B. Association for molecular pathology training and education committee. Standard mutation nomenclature in molecular diagnostics. practical and educational challenges. J. Mol. Diagn 2007, 9, 1–6. [Google Scholar]
- Tommasi, S.; Fedele, V.; Lacalamita, R.; Crapolicchioa, A.; Perlinob, E.; Bellizia, A.; Paradiso, A. Molecular and functional characteristics of ERBB2 in normal and cancer breast cells. Cancer Lett 2004, 209, 215–222. [Google Scholar]

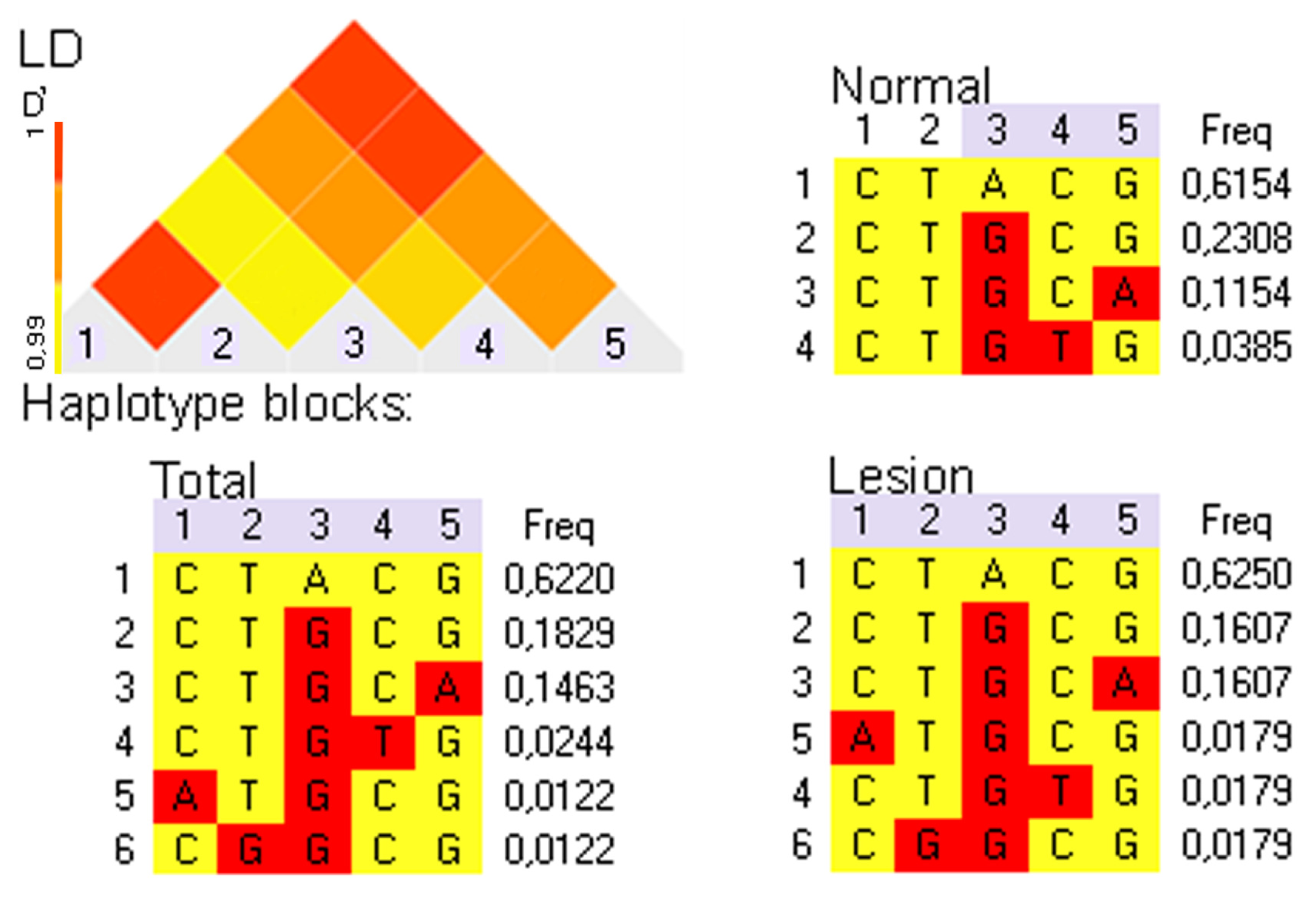

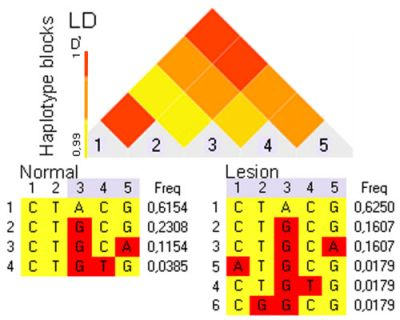{kind=link}
{kind=link}
{kind=link}
| A: cat-ERBB2_19-20 genomic sequence variants presence (+) or absence (0) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Normal DNA sequences (n = 16): | ||||||||||||||
| SVs | n = 7 | n = 1 | n = 3 | n = 2 | n = 1 | n = 2 | ||||||||
| g.[224A>G] + [wt] | 0 | 0 | + | 0 | + | + | ||||||||
| g.[224A>G] + [224A>G] | 0 | + | 0 | + | 0 | 0 | ||||||||
| g.[360C>T] + [wt] | 0 | 0 | 0 | 0 | + | 0 | ||||||||
| g.[371G>A] + [wt] | 0 | 0 | 0 | + | 0 | + | ||||||||
| Mammary Lesions DNA sequences (n = 22): | ||||||||||||||
| SVs | n = 8 | n = 5 | n = 1 | n = 1 | n = 1 | n = 4 | n = 1 | n = 1 | ||||||
| g.[153C>A] + [wt] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | + | ||||||
| g.[156T>G] + [wt] | 0 | 0 | + | 0 | 0 | 0 | 0 | 0 | ||||||
| g.[224A>G] + [wt] | 0 | + | + | 0 | + | + | + | |||||||
| g.[224A>G] + [224A>G] | + | + | ||||||||||||
| g.[360C>T] + [wt] | 0 | 0 | 0 | 0 | + | 0 | 0 | 0 | ||||||
| g.[371G>A] + [wt] | 0 | 0 | 0 | + | 0 | + | 0 | 0 | ||||||
| B: cat-ERBB2_19-20 genomic sequence variants frequency and Hardy-Weinberg equilibrium test | ||||||||||||||
| Allele frequencies (%): | ||||||||||||||
| allele | g.[153C>A] | allele | g.[156T>G] | allele | g.[224A>G] | allele | g.[360C>T] | allele | g.[371G>A] | |||||
| Lesion | Normal | Lesion | Normal | Lesion | Normal | Lesion | Normal | Lesion | Normal | |||||
| C | 97.7% | 100% | T | 97.7% | 100% | A | 36.4% | 62.5% | C | 97.7% | 96.9% | G | 88.6% | 87.5% |
| A | 2.3% | 0% | G | 2.3% | 0% | G | 63.6% | 37.5% | T | 2.3% | 3.1% | A | 11.3% | 12.5% |
| Hardy-Weinberg equilibrium: | ||||||||||||||
| p-value | g.[153C>A] | p-value | g.[156T>G] | p-value | g.[224A>G] | p-value | g.[360C>T] | p-value | g.[371G>A] | |||||
| Lesion | Normal | Lesion | Normal | Lesion | Normal | Lesion | Normal | Lesion | Normal | |||||
| 0.12E−6 | 0.12E−6 | 0.7 | 0.5 | 0.12E−6 | 0.31E−4 | 0.75 | 0.37 | |||||||
| Number of Masses | Primary Tumor Size | Diploid Genotype | |||
|---|---|---|---|---|---|
| T? | T1 | T2 | T3 | ||
| Multiple | 1 MaL * | 2 MaL | h1/h1 | ||
| 1 BeL * | h2/h2 | ||||
| 1 MaL | h1/h6 | ||||
| 1 BeL # | h1/h5 | ||||
| 1 MaL | h1/h4 | ||||
| 2 MaL * | h1/h3 | ||||
| 3 MaL | 1 MaL | 1 BeL | h1/h2 | ||
| 1 MaL | h2/h3 | ||||
| Simple | 1 MaL | 1 MaL | h1/h1 | ||
| 1 MaL | h1/h2 | ||||
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Santos, S.; Bastos, E.; Baptista, C.S.; Sá, D.; Caloustian, C.; Guedes-Pinto, H.; Gärtner, F.; Gut, I.G.; Chaves, R. Sequence Variants and Haplotype Analysis of Cat ERBB2 Gene: A Survey on Spontaneous Cat Mammary Neoplastic and Non-Neoplastic Lesions. Int. J. Mol. Sci. 2012, 13, 2783-2800. https://doi.org/10.3390/ijms13032783
Santos S, Bastos E, Baptista CS, Sá D, Caloustian C, Guedes-Pinto H, Gärtner F, Gut IG, Chaves R. Sequence Variants and Haplotype Analysis of Cat ERBB2 Gene: A Survey on Spontaneous Cat Mammary Neoplastic and Non-Neoplastic Lesions. International Journal of Molecular Sciences. 2012; 13(3):2783-2800. https://doi.org/10.3390/ijms13032783
Chicago/Turabian StyleSantos, Sara, Estela Bastos, Cláudia S. Baptista, Daniela Sá, Christophe Caloustian, Henrique Guedes-Pinto, Fátima Gärtner, Ivo G. Gut, and Raquel Chaves. 2012. "Sequence Variants and Haplotype Analysis of Cat ERBB2 Gene: A Survey on Spontaneous Cat Mammary Neoplastic and Non-Neoplastic Lesions" International Journal of Molecular Sciences 13, no. 3: 2783-2800. https://doi.org/10.3390/ijms13032783