Plant Regeneration and Somatic Embryogenesis from Immature Embryos Derived through Interspecific Hybridization among Different Carica Species

Abstract
:1. Introduction
2. Results and Discussion
2.1. Fruit Set and Embryo Rescue
2.2. Plant Regeneration from 90-Day-Old Embryos
2.3. Somatic Embryogenesis from 90-Day-Old Embryos
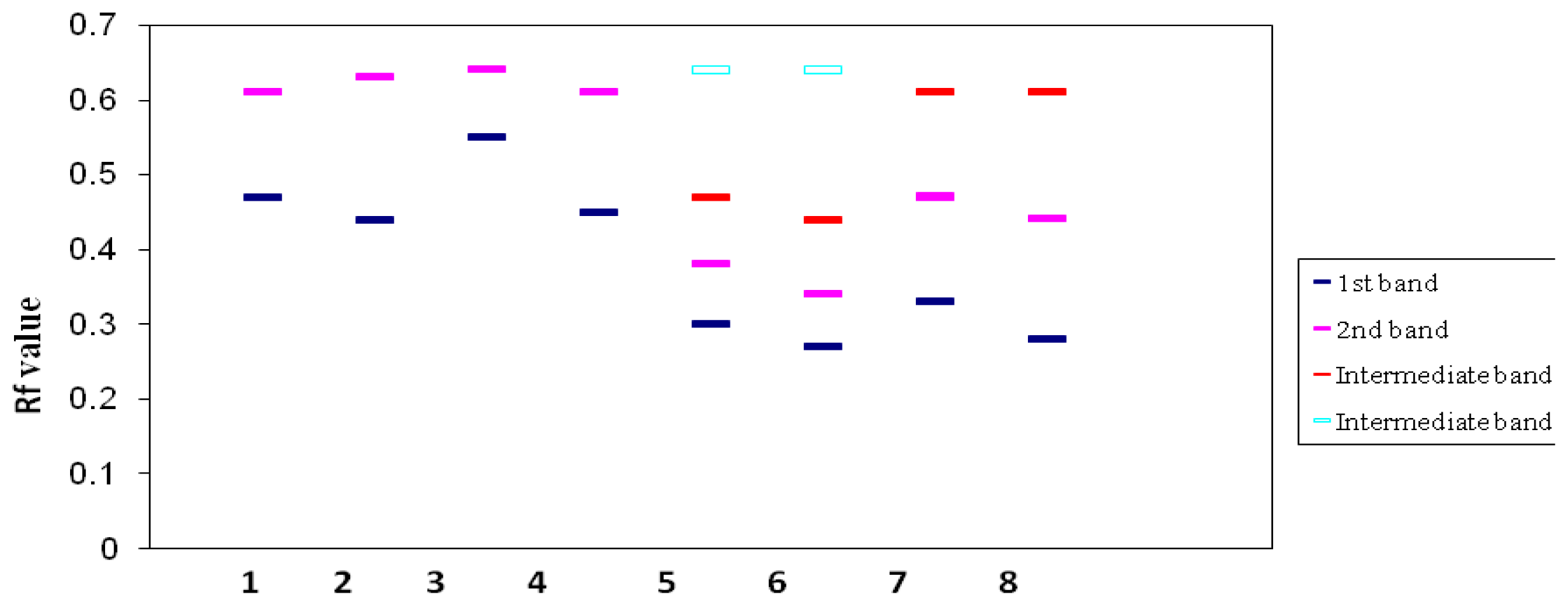
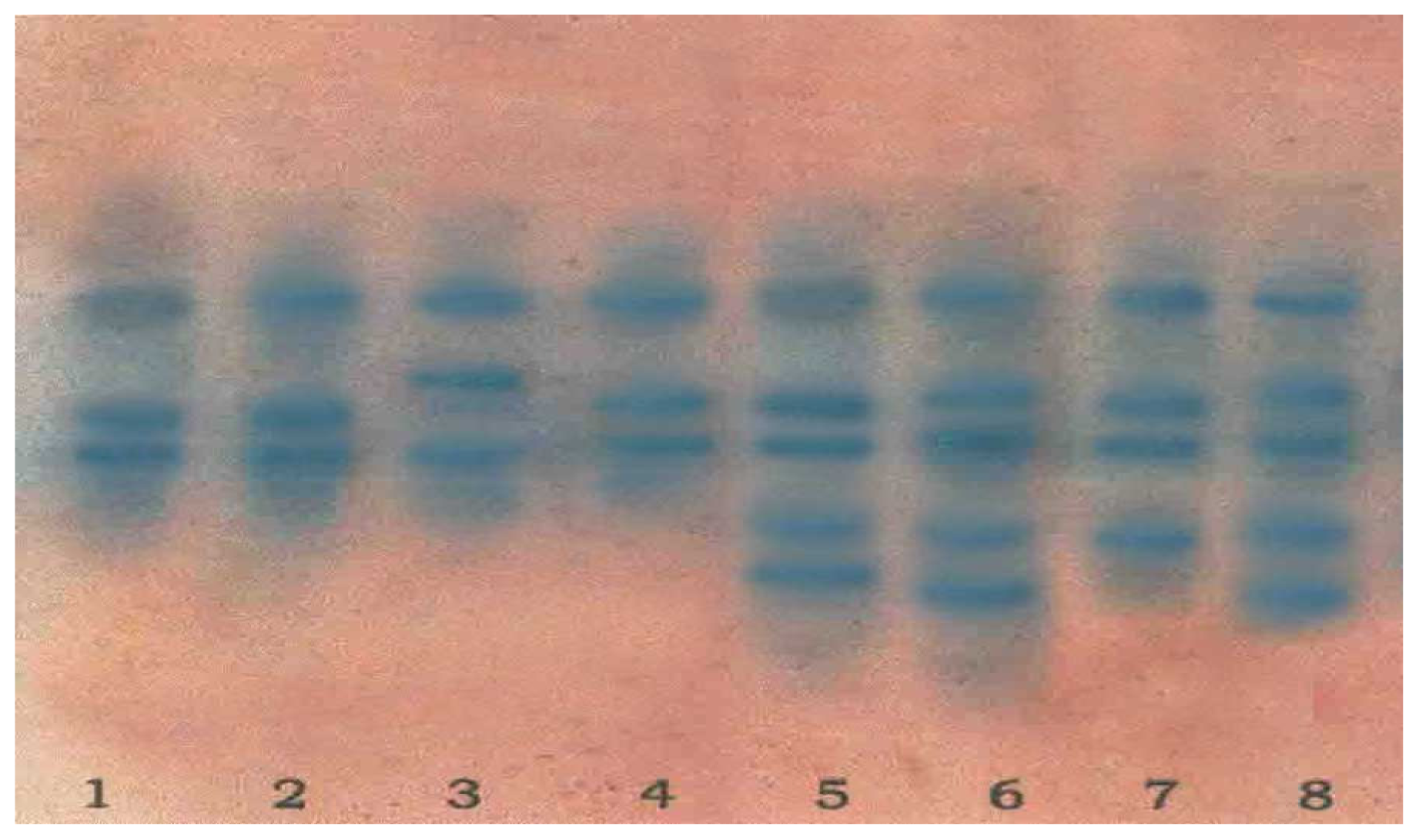
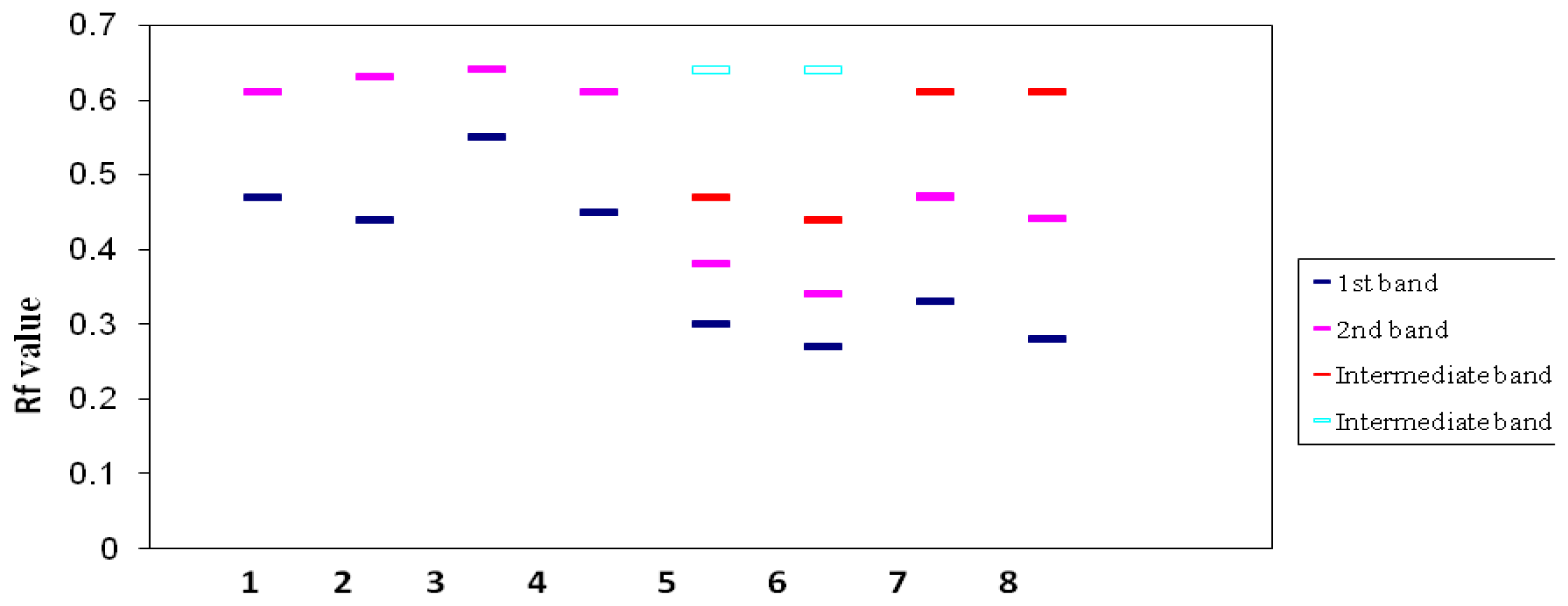
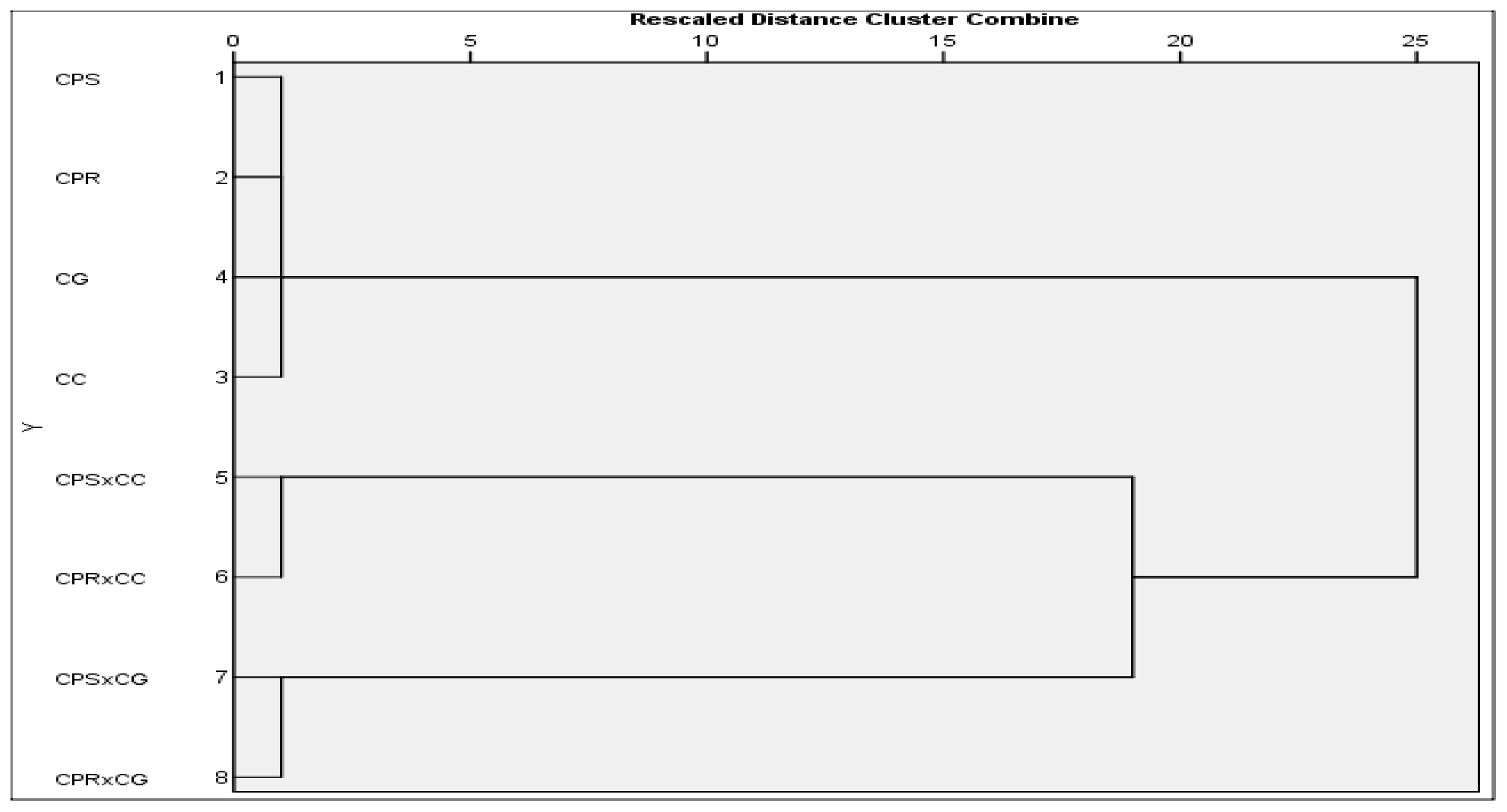
2.4. Isoenzyme Analysis
3. Experimental Section
3.1. Plant Materials and Hybridization Procedure
3.2. Plant Regeneration and Somatic Embryogenesis
3.3. Acclimatization of Plantlets
3.4. Isoenzyme Analysis
3.5. Data Collection and Analysis
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Bhattacharya, J.; Khuspe, S.S. In vitro and in vivo germination of papaya (Carica papaya L.) seeds. Sci. Hortic 2001, 91, 39–49. [Google Scholar]
- De Candolle, A. Origin of Cultivated Plants; John Wiley and Sons, Inc: New York, NY, USA, 1984; p. 281. [Google Scholar]
- Jayavalli, R.; Balamahon, T.N.; Manivannan, N.; Govindaraj, M. Breaking the inergeneric hybridization barrier in Carica papaya and Vasconcella cauliflora. Sci. Hortic 2011, 130, 787–794. [Google Scholar]
- Litz, R.E. Papaya. In Handbook of Plant Cell Culture; Evans, D.A., Sharp, W.R., Eds.; Macmillan: New York, NY, USA, 1984; Volume 2, pp. 349–368. [Google Scholar]
- Manshardt, R.M. Papaya. In Biotechnology of Perennial Fruit Crops; Hammerschlag, F.A., Litz, R.E., Eds.; CABI: London, UK, 1992; pp. 489–511. [Google Scholar]
- Gonsalves, D. Papaya Ringspot. In Compendium of Tropical Fruit Diseases; Ploetz, R.C., Ed.; APS Press: St. Paul, MN, USA, 1994; p. 67. [Google Scholar]
- Magdalia, P.M.; Village, V.N.; Pimentel, R.B.; Bayot, R.G. Reaction of papaya (Carica papaya L.) and related Carica species to ringspot virus. Philipp. J. Crop Sci 1988, 13, 129–132. [Google Scholar]
- Horovitz, S.; Jimenez, H. Cruzamientos interspecificos intergenericos en caricaceas y sus implicaciones fitotecnicas. Agron. Trop 1967, 17, 323–343. [Google Scholar]
- Manshardt, R.M.; Wenslaff, T.F. Zygotic polyembryony in interspecific hybrids of Carica papaya and Carica cauliflora. J. Am. Soc. Hortic. Sci. 1989, 114, 684–689. [Google Scholar]
- Khuspe, S.; Hendre, R.; Mascarenhas, A.; Jagannathan, V.; Thombre, M.; Joshi, A. Utilization of Tissue Culture Isolate in Interspecific Hybrids in Carica. In Plant Tissue Culture, Genetic Manipulation and Somatic hybridization of Plant Cells; Rao, P.S., Heble, M., Eds.; Bhaba Atomic Research Center: Bombay, India, 1980; pp. 198–205. [Google Scholar]
- Rabbani, M.G. Induction, Regeneration and Synchronisation of Somatic Embryos in Papaya (Carica papaya L.). In Ph.D. Thesis; University of London: London, UK, 1992; p. 268. [Google Scholar]
- Magdalita, P.M.; Adkins, S.W.; Godwin, I.D.; Drew, R.A. An improved embryo rescue protocol for a Carica interspecific hybrid. Aust. J. Bot 1996, 44, 343–353. [Google Scholar]
- Fitch, M.M.M.; Manshardt, R.M. Somatic embryogenesis and plant regeneration from immature zygotic embryos of papaya (Carica papaya L.). Plant Cell Rep 1990, 9, 320–324. [Google Scholar]
- Teixeira da Silva, J.A.; Rashid, Z.; tan Nut, D.; Sivakumar, D.; Gera, A.; Souza, M.T., Jr; Tennant, P.F. Papaya (Carica papaya L.) Biology and Biotechnology. Tree For. Sci. Biotech. 2007, 1, 47–73. [Google Scholar]
- Moore, G.A.; Litz, R.E. Biochemical markers of Carica papaya, C. cauliflora and plants from somatic embryos of their hybrid. J. Am. Soc. Hortic. Sci 1984, 109, 213–218. [Google Scholar]
- Malabadi, R.B.; Kumar, S.V.; Mulgund, G.S.; Nataraja, K. Induction of somatic embryogenesis in papaya (Carica papaya). Res. Biotech 2011, 2, 40–55. [Google Scholar]
- Anandan, R.; Sudhakar, D.; Balasubramanian, P.; Gutieirrez-Mora, A. In vitro somatic embryogenesis from suspension cultures of Carica papaya L. Sci. Hortic 2012, 136, 43–49. [Google Scholar]
- Farzana, A.R.F.; Palkadapala, P.G.V.N.; Meddegoda, K.M.M.N.; Samarajeewaand, P.K.; Eeswara, J.P. Somatic embryogenesis in Carica papaya L. cv. Rathna. J. Natn. Sci. Foundation Sri Lanka 2008, 36, 41–50. [Google Scholar]
- Romyanon, K.; Boonthum, M.; Attathom, S. Revised protocols for high efficient transformation and regeneration of somatic embryos of papaya (Carica papaya L.). Acta Hortic 2007, 740, 147–152. [Google Scholar]
- Litz, R.E.; Conover, R.A. Highfrequency somatic embryogenesis from Carica suspension cultures. Ann. Bot 1983, 51, 683–686. [Google Scholar]
- Chen, M.H.; Chen, C.C.; Wang, D.N.; Chen, F.C. Somatic embryogenesis and plant regeneration from immature embryos of Carica papaya × Carica cauliflora cultured in vitro. Can. J. Bot 1991, 69, 1913–1918. [Google Scholar]
- Drew, R.A.; Magdalita, P.M.; O’Brien, C.M. Development of Carica interspecific hybrids. Acta Hortic 1998, 461, 285–292. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar]
- Jobin-Décor, M.P.; Graham, G.C.; Henry, R.J.; Drew, R.A. RAPD and isozyme analysis of genetic relationships between C. papaya and wild relatives. Genet. Resour. Crop Ev 1997, 44, 471–477. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cross | Fruit set (%) (Mean ± SE) | Number of embryos per fruit | |||
|---|---|---|---|---|---|
| 60 days after pollination (Mean ± SE) | 90 days after pollination (Mean ± SE) | 120 days after pollination (Mean ± SE) | 150 days after pollination (Mean ± SE) | ||
| CPS × CC | 64.67 ± 2.27 a | 13.33 ± 0.63 bc | 18.30 ± 0.55ab | 3.30 ± 0.40 a | 0.00 ± 0.00 a |
| CPS × CG | 59.73 ± 2.17 b | 18.36 ± 1.03 a | 20.53 ± 0.84 a | 3.76 ± 0.21 a | 0.00 ± 0.00 a |
| CPR × CC | 58.10 ± 1.36 b | 11.70 ± 0.94 c | 15.40 ± 1.07 b | 1.93 ± 0.37 b | 0.00 ± 0.00 a |
| CPR × CG | 54.36 ± 2.04 c | 15.43 ± 0.41 b | 17.07 ± 1.04 ab | 2.37 ± 0.17 b | 0.00 ± 0.00 a |
| Cross | Germination (%) (Mean ± SE) | Days to shooting (Mean ± SE) | Days to rooting (Mean ± SE) | Plant height (cm) (Mean ± SE) | No. of leaves per plant (Mean ± SE) | No. of roots per plant (Mean ± SE) |
|---|---|---|---|---|---|---|
| CPS × CC | 74.13 ± 2.25 a | 13.47 ± 0.61 c | 17.23 ± 0.83 b | 4.26 ± 0.17 a | 5.60 ± 0.17 a | 6.81 ± 0.24 a |
| CPS × CG | 5.80 ± 1.99 b | 16.57 ± 0.57 ab | 19.50 ± 0.63 ab | 3.50 ± 0.17 b | 5.50 ± 0.17 a | 6.13 ± 0.20 a |
| CPR × CC | 62.43 ± 1.64 b | 14.87 ± 0.82 bc | 17.47 ± 0.72 b | 3.43 ± 0.18 b | 5.43 ± 0.23 a | 6.48 ± 0.21 a |
| CPR × CG | 60.67 ± 1.04 b | 18.27 ± 0.54 a | 20.70 ± 0.60 a | 3.33 ± 0.08 b | 4.63 ± 0.08 b | 5.78 ± 0.09 bc |
| Cross | Hybrid embryo producing callus (%) (Mean ± SE) | Days required for callusing (Mean ± SE) | Hybrid embryo producing embryogenic callus (%) (Mean ± SE) | Days required for embryogenic callus (Mean ± SE) | Number of somatic embryo per hybrid embryo (Mean ± SE) | Number of plantlets per hybrid embryo (Mean ± SE) |
|---|---|---|---|---|---|---|
| CPS × CC | 68.03 ± 1.46 a | 18.30 ± 0.60 a | 48.37 ± 2.22 a | 38.43 ± 1.01 a | 14.13 ± 0.40 a | 10.33 ± 0.52 a |
| CPS × CG | 54.80 ± 2.23 c | 15.47 ± 0.49 b | 38.40 ± 1.21 c | 29.47 ± 0.63 c | 9.80 ± 0.23 c | 6.60 ± 0.51 bc |
| CPR × CC | 64.40 ± 1.15 ab | 17.10 ± 0.62 ab | 47.20 ± 1.80 ab | 33.63 ± 178 b | 12.57 ± 1.21 ab | 9.41 ± 0.61 a |
| CPR × CG | 60.80 ± 1.43 b | 16.56 ± 0.52 ab | 41.47 ± 1.92 bc | 31.10 ± 0.81 bc | 11.40 ± 0.69 bc | 8.37 ± 0.52 ab |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Azad, M.A.K.; Rabbani, M.G.; Amin, L. Plant Regeneration and Somatic Embryogenesis from Immature Embryos Derived through Interspecific Hybridization among Different Carica Species. Int. J. Mol. Sci. 2012, 13, 17065-17076. https://doi.org/10.3390/ijms131217065
Azad MAK, Rabbani MG, Amin L. Plant Regeneration and Somatic Embryogenesis from Immature Embryos Derived through Interspecific Hybridization among Different Carica Species. International Journal of Molecular Sciences. 2012; 13(12):17065-17076. https://doi.org/10.3390/ijms131217065
Chicago/Turabian StyleAzad, Md. Abul Kalam, Md. Golam Rabbani, and Latifah Amin. 2012. "Plant Regeneration and Somatic Embryogenesis from Immature Embryos Derived through Interspecific Hybridization among Different Carica Species" International Journal of Molecular Sciences 13, no. 12: 17065-17076. https://doi.org/10.3390/ijms131217065