Anti-Legionella dumoffii Activity of Galleria mellonella Defensin and Apolipophorin III


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
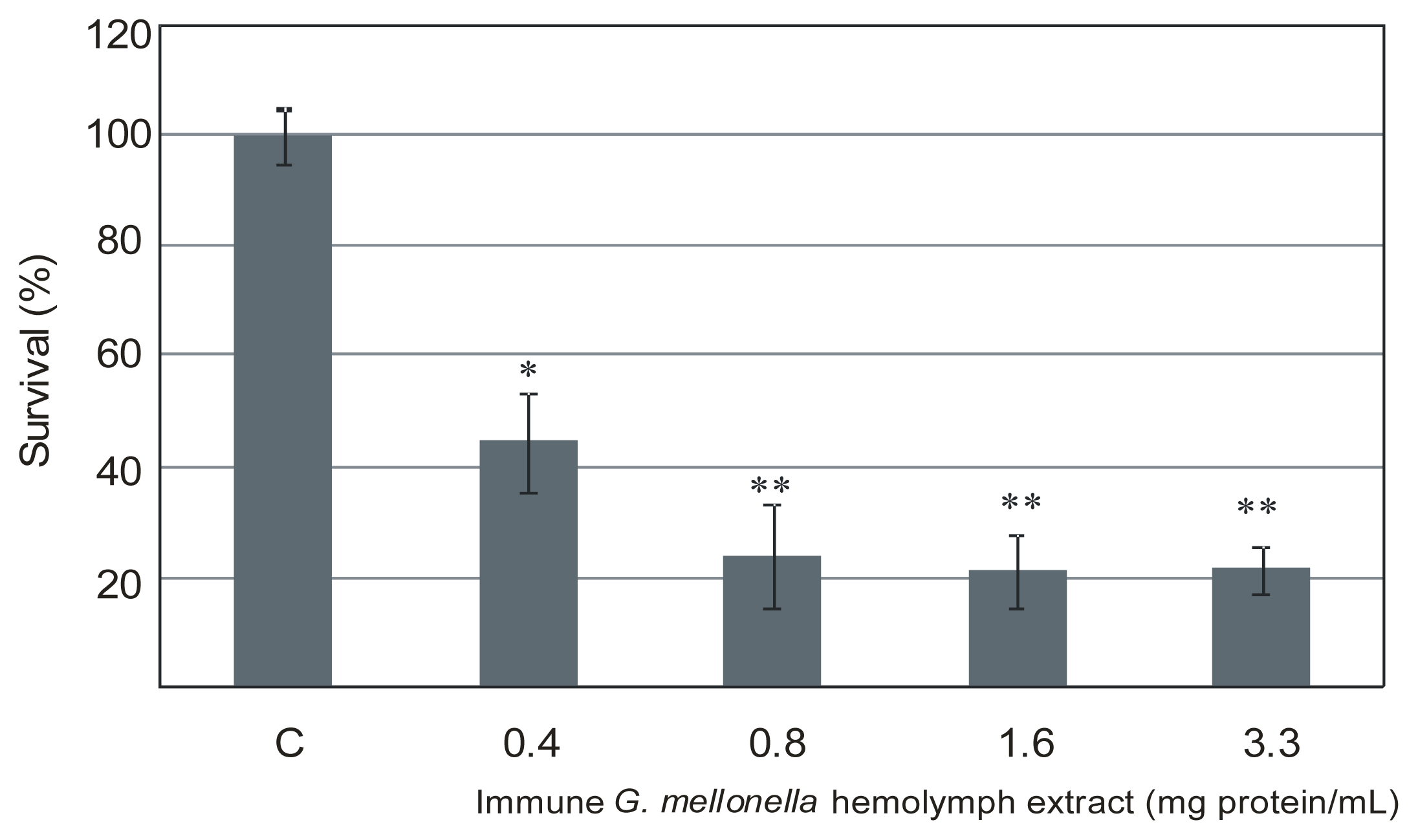
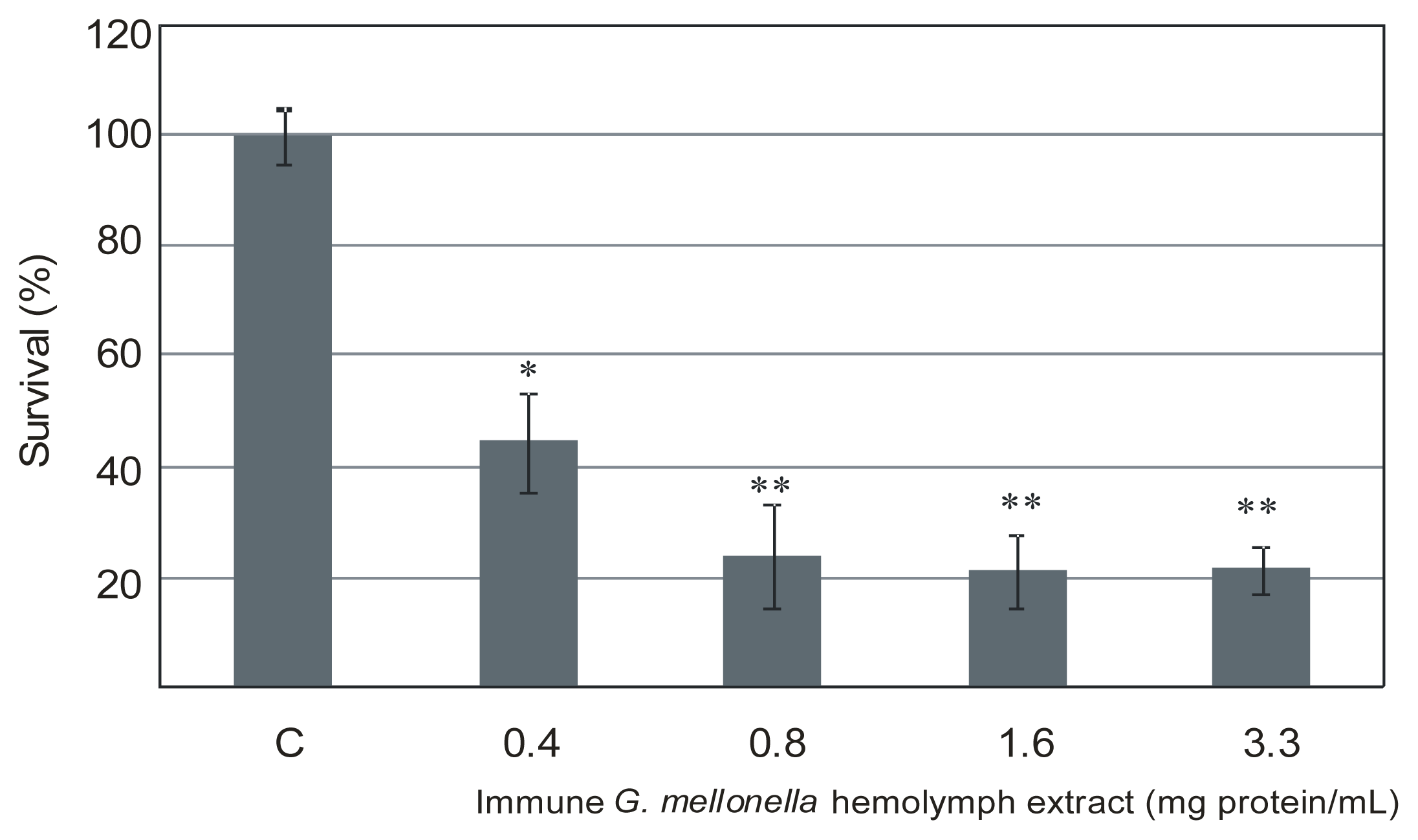
2.1. The Effect of Extracts of Immune G. mellonella Hemolymph on L. dumoffii Survival Rate
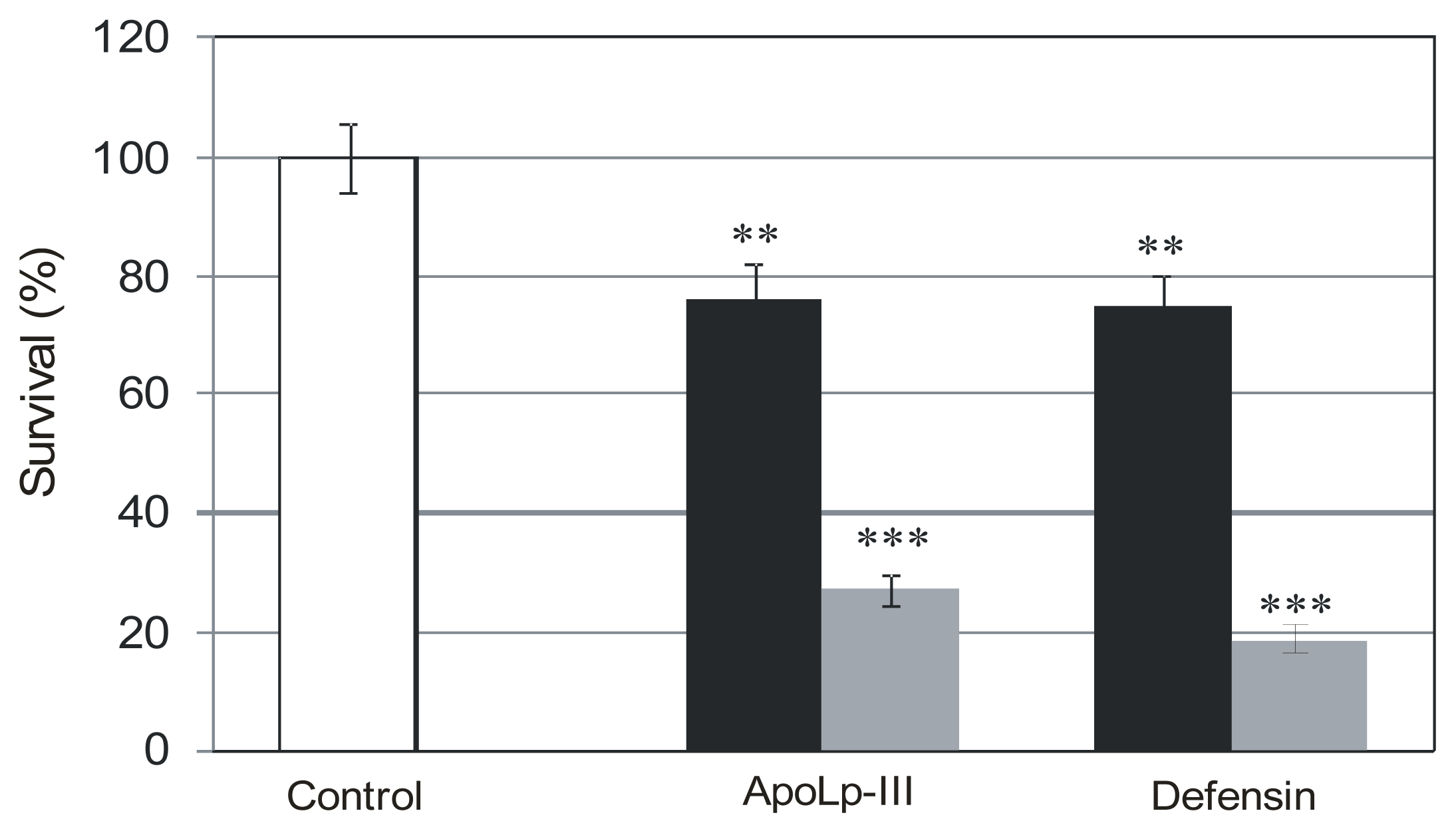
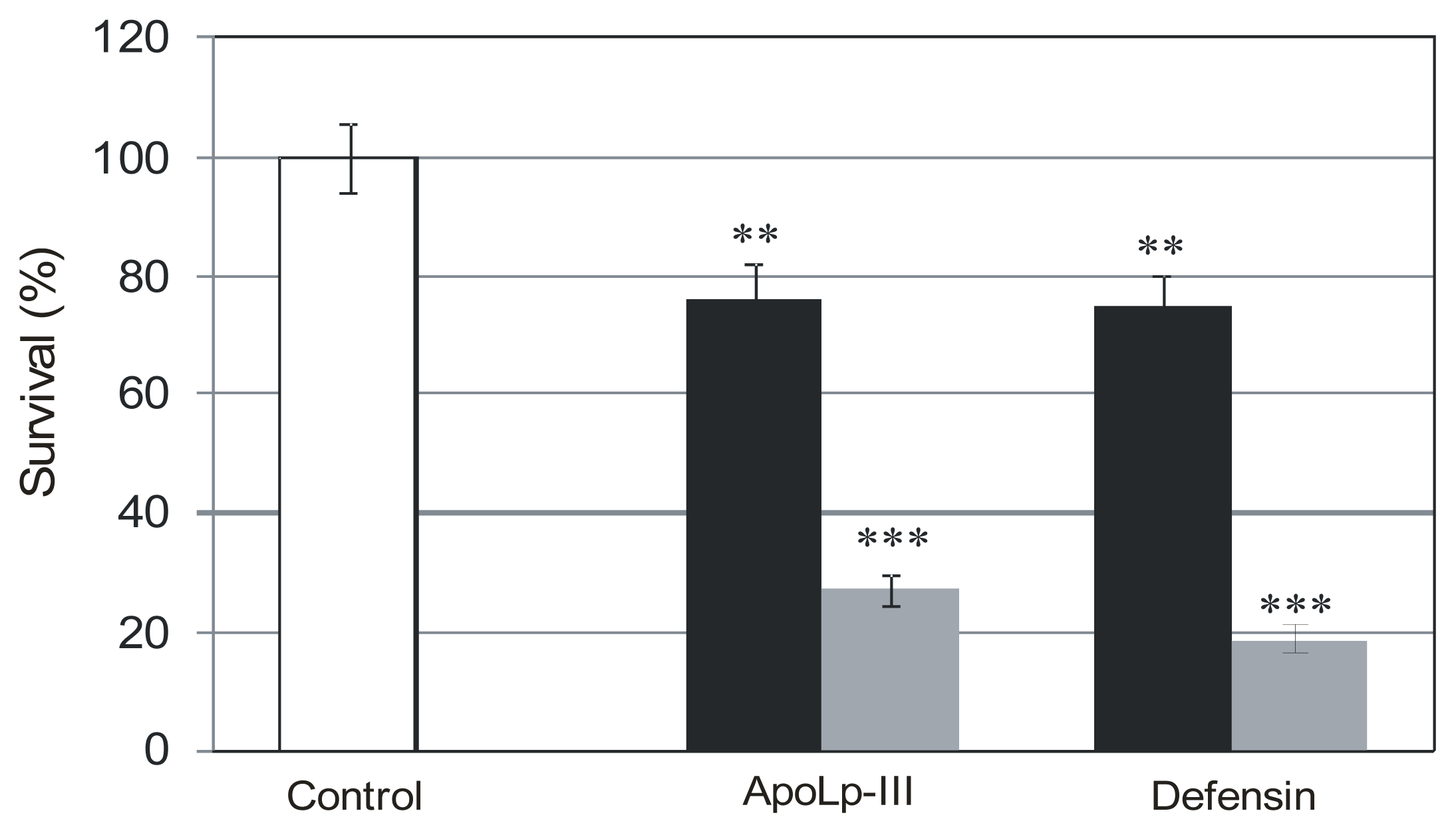
2.2. The Effect of G. mellonella Defensin and apoLp-III on Survival Rate of L. dumoffii Cultured with and without Choline Supplementation
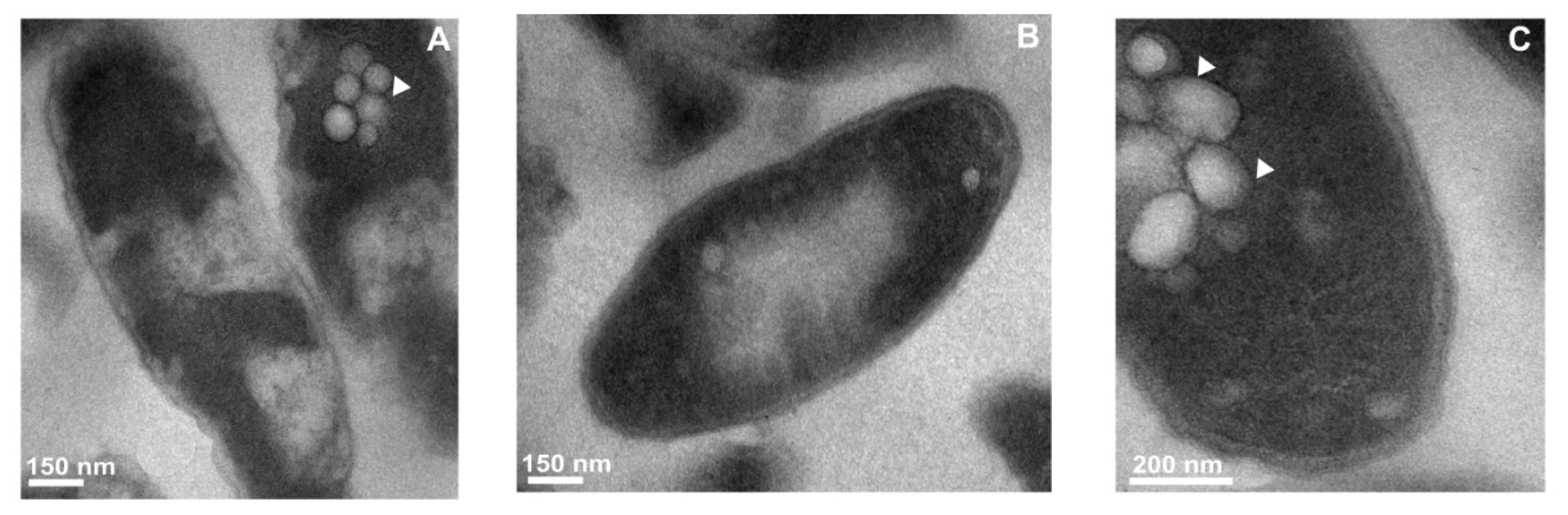
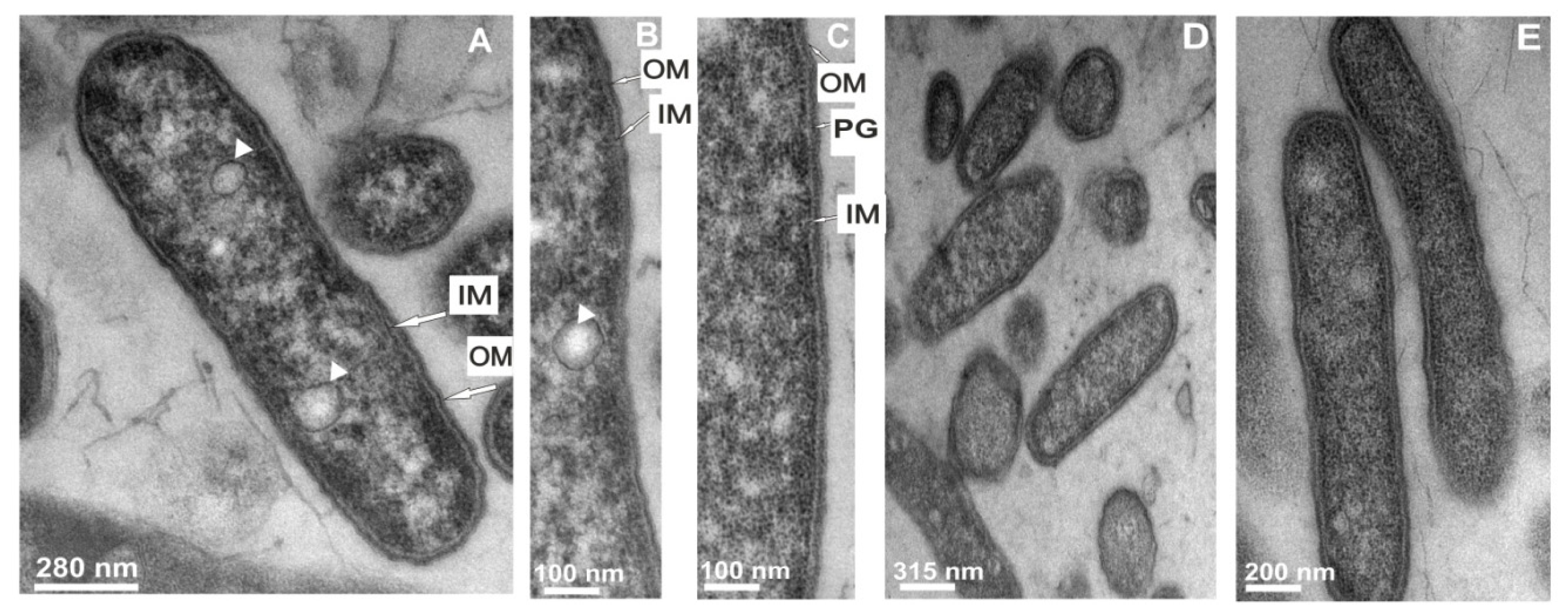
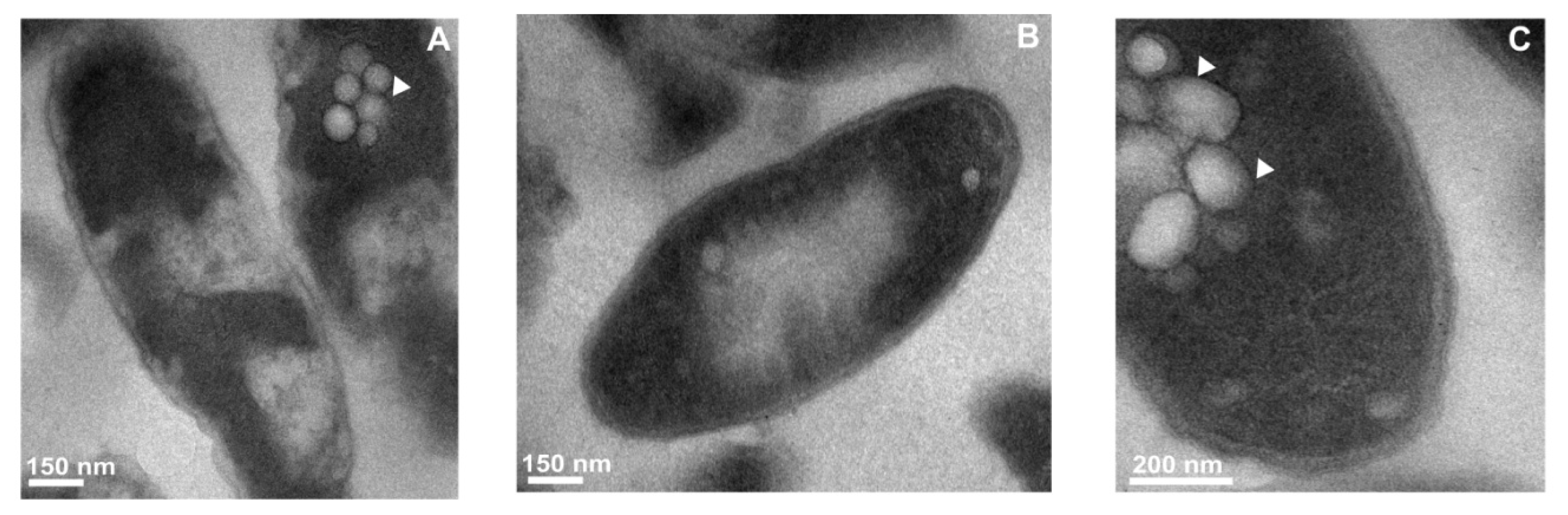
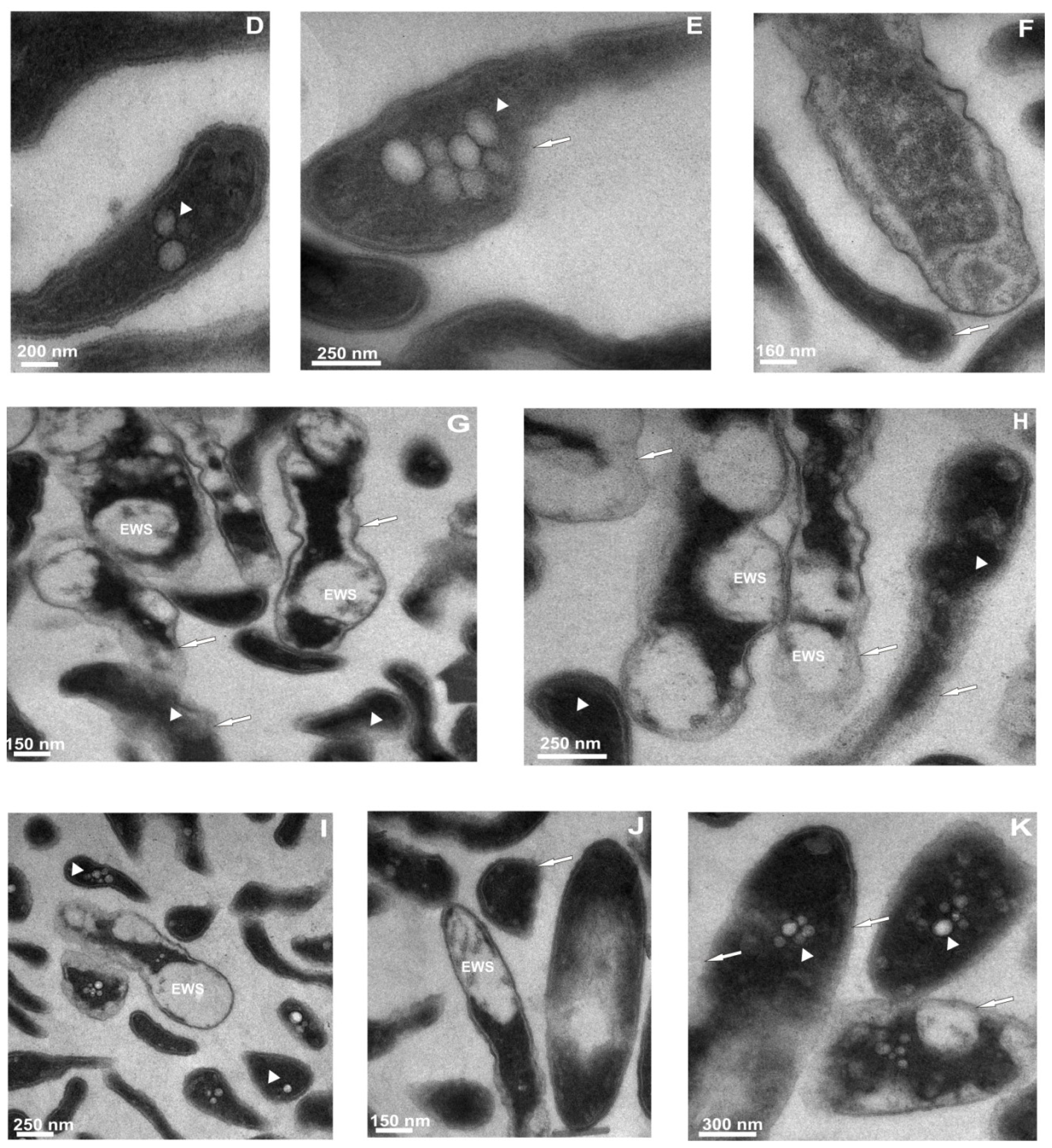
2.3. Transmission Electron Microscope Observations of Legionella dumoffii
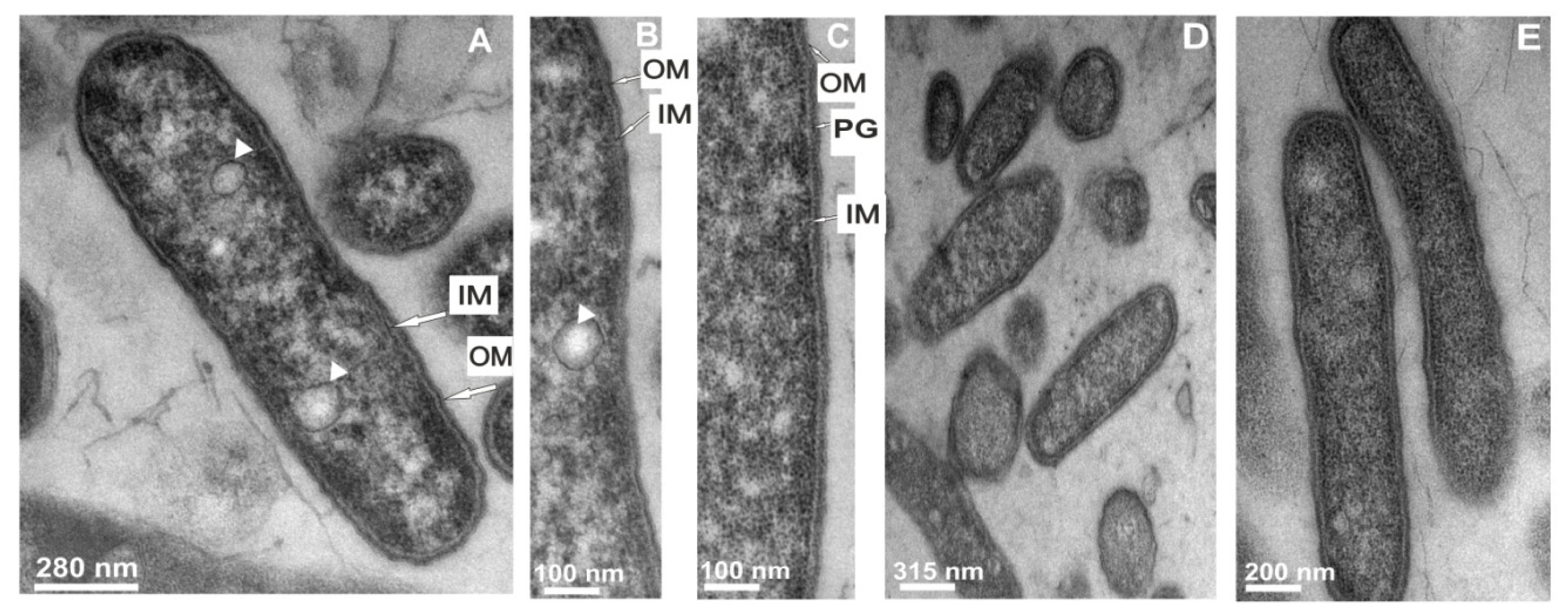
2.3.1. The Changes in the L. dumoffii Cell Morphology under the Influence of G. mellonella Immune Hemolymph Extract
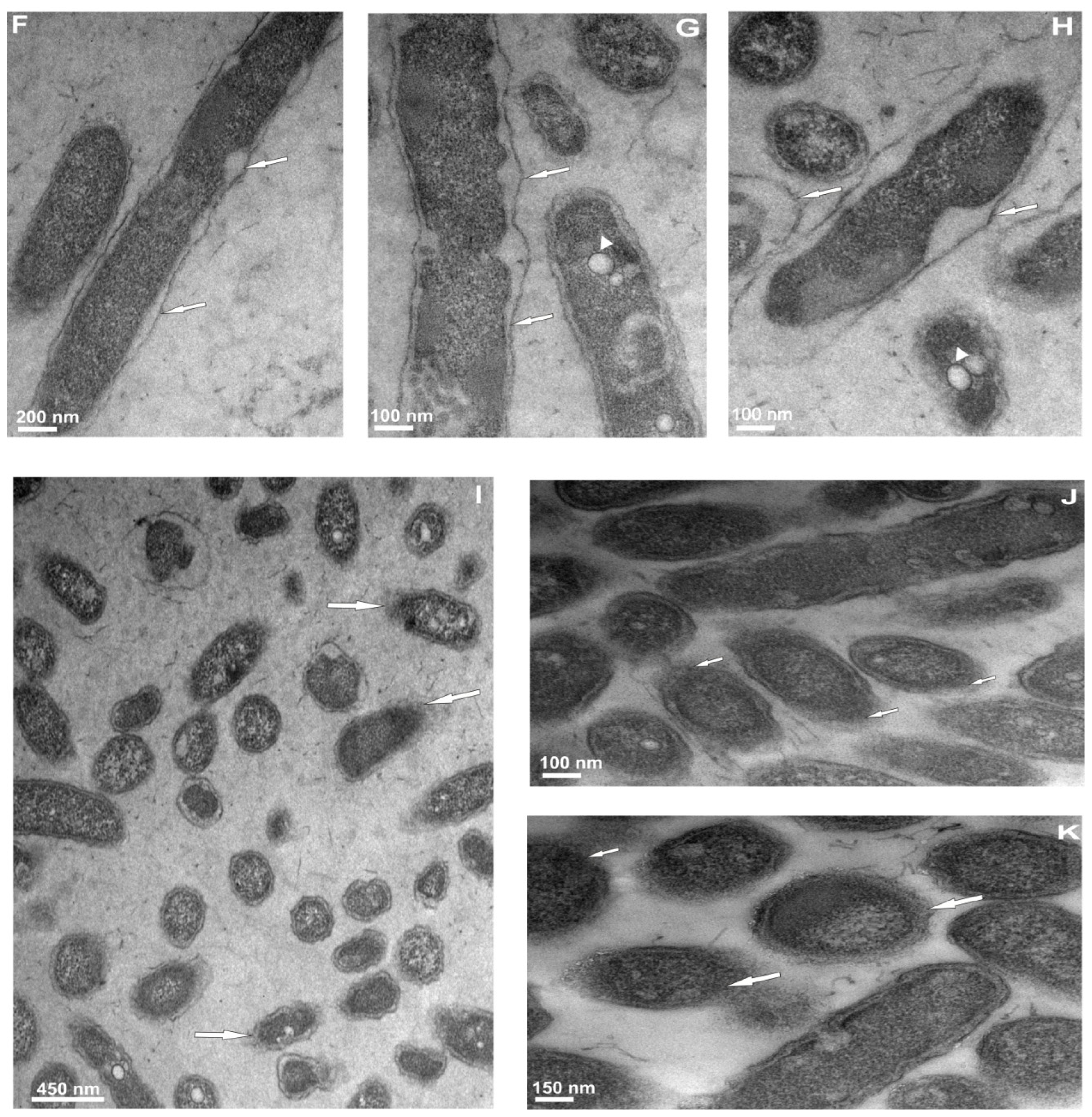
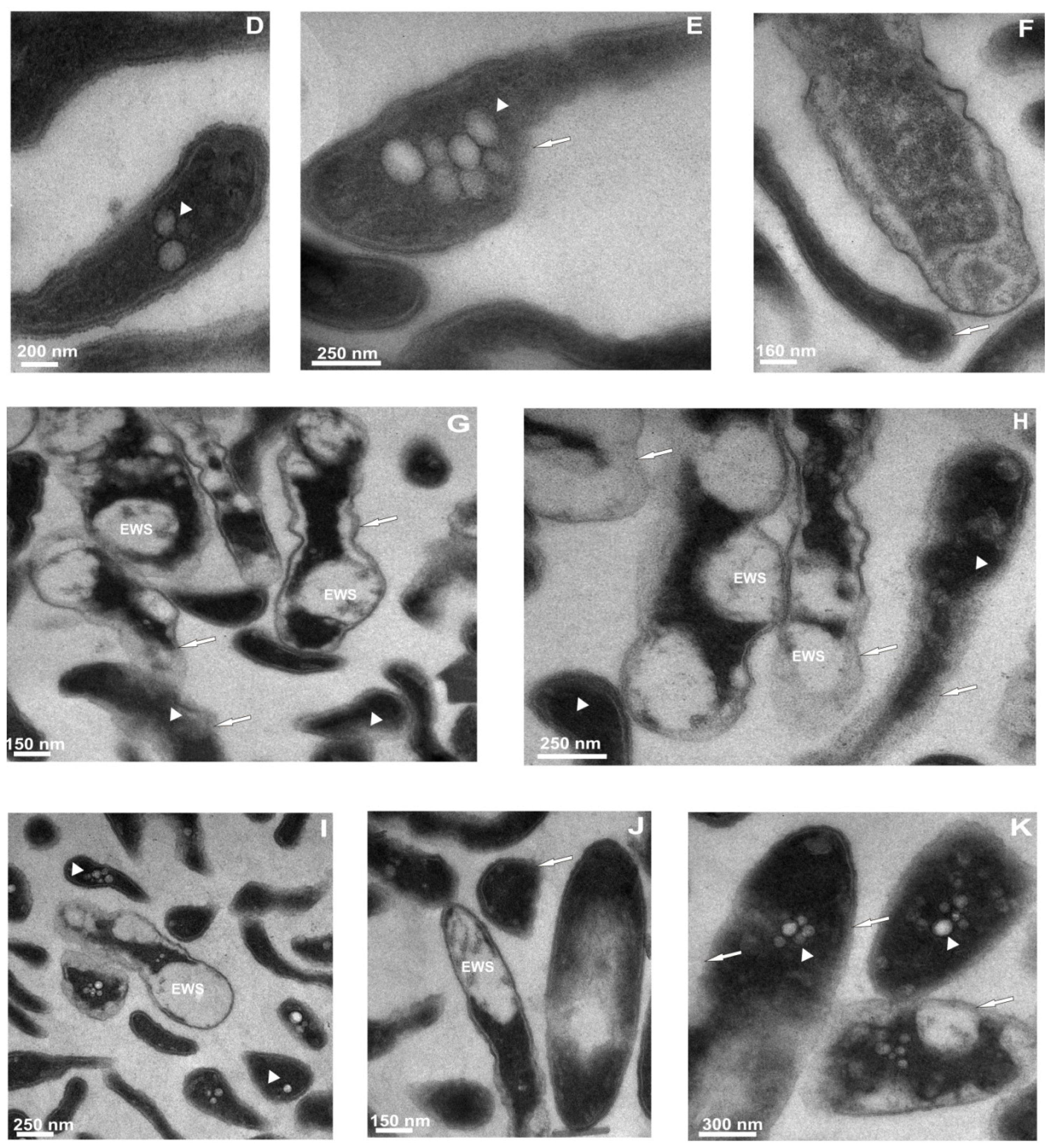
2.3.2. Ultrastructural Changes in the Cells of L. dumoffii after Treatment with Defensin and apoLp-III Isolated from G. mellonella Hemolymph
3. Discussion
4. Experimental Section
4.1. Bacterial Strain and Culture Conditions
4.2. Insect Culture and Immune Challenge
4.3. Preparation of Methanolic Extracts of G. mellonella Hemolymph
4.4. Purification of G. mellonella Defense Peptides and Proteins
4.5. Anti-Legionella Activity Assays
4.5.1. Radial Diffusion Assay
4.5.2. Colony Counting Assay
4.6. Preparation of Samples for Microscopic Analysis
4.7. Transmission Electron Microscopy Analysis of L. dumoffii Cells
4.8. Other Methods
4.9. Statistical Analysis
5. Conclusions
Acknowledgements
- Conflict of InterestThe authors declare no conflict of interest.
References
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev 2002, 15, 506–526. [Google Scholar]
- Yu, V.L.; Plouffe, J.F.; Pastoris, M.C.; Stout, J.E.; Schousboe, M.; Widmer, A.; Summersgill, J.; File, T.; Heath, C.M.; Paterson, D.L.; et al. Distribution of Legionella species and serogroups isolated by culture in patients with sporadic community-acquired legionellosis: An international collaborative survey. J. Infect. Dis 2002, 186, 127–128. [Google Scholar]
- Fujita, I.; Tsuboi, H.; Ohotsuka, M.; Sano, I.; Murakami, Y.; Akioka, H.; Hayashi, M. Legionella dumoffii and Legionella pneumophila serogroup 5 isolated from 2 cases of fulminant pneumonia. J. Jpn. Assoc. Infect. Dis 1989, 63, 801–810. [Google Scholar]
- Flendrie, M.; Jeurissen, M.; Franssen, M.; Kwa, D.; Klaassen, C.; Vos, F. Septic arthritis caused by Legionella dumoffii in a patient with systemic lupus erythematosus-like disease. J. Clin. Microbiol 2011, 49, 746–749. [Google Scholar]
- Tompkins, L.S.; Roessler, B.J.; Redd, S.C.; Markowitz, L.E.; Cohen, M.L. Legionella prosthetic-valve endocarditis. N. Engl. J. Med 1988, 318, 530–535. [Google Scholar]
- Neumeister, B.; Schoniger, S.; Faigle, M.; Eichner, M.; Dietz, K. Multiplication of different Legionella species in Mono Mac 6 cells and in Acantamoeba castellanii. Appl. Environ. Microbiol 1997, 63, 1219–1224. [Google Scholar]
- Wadowsky, R.M.; Wilson, T.M.; Kapp, N.J.; West, A.J.; Kuchta, J.M.; States, S.J.; Dowling, J.N.; Yee, R.B. Multiplication of Legionella spp. in tap water containing Hartmannella vermiformis. Appl. Environ. Microbiol 1991, 57, 1950–1955. [Google Scholar]
- O’Connell, W.A.; Dhand, L.; Cianciotto, N.P. Infection of macrophage-like cells by Legionella species that have not been associated with disease. Infect. Immun 1996, 64, 4381–4384. [Google Scholar]
- Maruta, K.; Miyamoto, H.; Hamada, T.; Ogawa, M.; Taniguchi, H.; Yoshida, S.I. Entry and intracellular growth of Legionella dumoffii in alveolar epithelial cells. Am. Respir. Crit. Care Med 1998, 157, 1967–1974. [Google Scholar]
- Scharf, S.; Vardarova, K.; Lang, F.; Schmeck, B.; Opitz, B.; Flieger, A.; Heuner, K.; Hippenstiel, S.; Suttorp, N.; N’Guessan, P.D. Legionella pneumophila induces human beta Defensin-3 in pulmonary cells. Respir. Res 2010, 11, 93. [Google Scholar]
- Scharf, S.; Hippenstiel, S.; Flieger, A.; Suttorp, N.; N’Guessan, P.D. Induction of human β-defensin-2 in pulmonary epithelial cells by Legionella pneumophila: Involvement of TLR2 and TLR5, p38 MAPK, JNK, NF-κB, and AP-1. Am. J. Physiol. Lung Cell Mol. Physiol 2010, 298, 687–695. [Google Scholar]
- Bulet, P.; Stöcklin, R. Insect antimicrobial peptides: Structures, properties and gene regulation. Prot. Pept. Lett 2005, 12, 3–11. [Google Scholar]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev 2004, 198, 169–184. [Google Scholar]
- Casteels, P.; Romagnolo, J.; Castle, M.; Casteels-Josson, K.; Erdjument-Bromage, H.; Tempst, P. Biodiversity of apidaecin-type peptide antibiotics. J. Biol. Chem. 1994, 269, 26107–26115. [Google Scholar]
- Lee, Y.S.; Yun, E.K.; Jang, W.S.; Kim, I.; Lee, J.H.; Park, S.Y.; Ryu, K.S.; Seo, S.J.; Kim, C.H.; Lee, I.H. Purification, cDNA cloning and expression of an insect defensin from the great wax moth, Galleria mellonella. Insect. Mol. Biol 2004, 13, 65–72. [Google Scholar]
- Kim, C.H.; Lee, J.H.; Kim, I.; Seo, S.J.; Son, S.M.; Lee, K.Y. Purification and cDNA cloning of a cecropin-like peptide from the great wax moth, Galleria mellonella. Mol. Cells 2004, 17, 262–266. [Google Scholar]
- Cytryńska, M.; Mak, P.; Zdybicka-Barabas, A.; Suder, P.; Jakubowicz, T. Purification and characterization of eight peptides from Galleria mellonella immune hemolymph. Peptides 2007, 28, 533–546. [Google Scholar]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. The discovery and analysis of a diverged family of novel antifungal moricin-like peptides in the wax moth Galleria mellonella. Insect Biochem. Mol. Biol 2008, 38, 201–212. [Google Scholar]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. A peptidomic study reveals the impressive antimicrobial peptide arsenal of the wax moth Galleria mellonella. Insect Biochem. Mol. Biol 2009, 39, 792–800. [Google Scholar]
- Schuhmann, B.; Seitz, V.; Vilcinskas, A.; Podsiadlowski, L. Cloning and expression of gallerimycin, an antifungal peptide expressed in immune response of greater wax moth larvae, Galleria mellonella. Arch. Insect Biochem. Physiol 2003, 53, 125–133. [Google Scholar]
- Mak, P.; Chmiel, D.; Gacek, G.J. Antibacterial peptides of the moth Galleria mellonella. Acta Biochim. Pol 2001, 48, 1191–1195. [Google Scholar]
- Weers, P.M.; Ryan, R.O. Apolipophorin III: Role model apolipoprotein. Insect Biochem. Mol. Biol 2006, 36, 231–240. [Google Scholar]
- Zdybicka-Barabas, A.; Cytryńska, M. Involvement of apolipophorin III in antibacterial defense of Galleria mellonella larvae. Comp. Biochem. Physiol. B 2011, 158, 90–98. [Google Scholar]
- Zdybicka-Barabas, A.; Januszanis, B.; Mak, P.; Cytryńska, M. An atomic force microscopy study of Galleria mellonella apolipophorin III effect on bacteria. Biochim. Biophys. Acta 2011, 1808, 1896–1906. [Google Scholar]
- Hatters, D.M.; Peters-Libeu, C.A.; Weisgraber, K.H. Apolipoprotein E structure: Insights into function. Trends Biochem. Sci 2006, 31, 445–454. [Google Scholar]
- Mahley, R.W.; Rall, S.C., Jr. Apolipoprotein E: Far more than a lipid transport protein. Annu. Rev. Genomics Hum. Genet. 2000, 1, 507–537. [Google Scholar]
- De Bont, N.; Netea, M.G.; Demacker, P.N.M.; Verschueren, I.; Kullberg, B.J.; van Dijk, K.W.; van der Meer, J.W.; Stalenhoef, A.F. Apolipoprotein E knock-out mice are highly susceptible to endotoxemia and Klebsiella pneumoniae infection. J. Lipid Res 1999, 40, 680–685. [Google Scholar]
- Roselaar, S.E.; Daugherty, A. Apolipoprotein E-deficient mice have impaired innate immune responses to Listeria monocytogenes in vivo. J. Lipid Res 1998, 39, 1740–1743. [Google Scholar]
- Van Oosten, M.; Rensen, P.C.; van Amersfoort, E.S.; van Eck, M.; van Dam, A.M.; Brevé, J.J.; Vogel, T.; Panet, A.; van Berkel, T.J.; Kuiper, J. Apolipoprotein E protects against bacterial lipopolysaccharide-induced lethality. A new therapeutic approach to treat gram-negative sepsis. J. Biol. Chem 2001, 276, 8820–8824. [Google Scholar]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol 2011, 29, 464–472. [Google Scholar]
- Teixeira, V.; Feio, M.J.; Bastos, M. Role of lipids in the interaction of antimicrobial peptides with membranes. Prog. Lipid Res 2012, 51, 149–177. [Google Scholar]
- Mak, P.; Zdybicka-Barabas, A.; Cytryńska, M. A different repertoire of Galleria mellonella antimicrobial peptides in larvae challenged with bacteria and fungi. Dev. Comp. Immunol 2010, 34, 1129–1136. [Google Scholar]
- Harding, C.R.; Schroeder, G.N.; Reynolds, S.; Kosta, A.; Collins, J.W.; Mousnier, A.; Frankel, G. Legionella pneumophila pathogenesis in the Galleria mellonella infection model. Infect. Immun 2012, 80, 2780–2790. [Google Scholar]
- Palusinska-Szysz, M.; Kalitynski, R.; Russa, R.; Dawidowicz, A.L.; Drozanski, W.J. Cellular envelope phospholipids from Legionella lytica. FEMS Microbiol. Lett 2008, 283, 239–246. [Google Scholar]
- Palusinska-Szysz, M.; Janczarek, M.; Kalitynski, R.; Dawidowicz, A.L.; Russa, R. Legionella bozemanae synthesizes phosphatidylcholine from exogenous choline. Microbiol. Res 2011, 166, 87–98. [Google Scholar]
- Martínez-Morales, F.; Schobert, M.; López-Lara, I.M.; Geiger, O. Pathways for phosphatidylcholine biosynthesis in bacteria. Microbiology 2003, 149, 3461–3471. [Google Scholar]
- Wang, W.; Smith, D.K.; Moulding, K.; Chen, H.M. The dependence of membrane permeability by the antibacterial peptide cecropin B and its analogs, CB-1 and CB-3, on liposomes of different composition. J. Biol. Chem 1998, 273, 27438–27448. [Google Scholar]
- Lysenko, E.S.; Gould, J.; Bals, R.; Wilson, J.; Weiser, J. Bacterial phosphorylcholine decreases susceptibility to the antimicrobial peptide LL-37/hCAP18 expressed in the upper respiratory tract. Infect. Immun 2000, 68, 1664–1671. [Google Scholar]
- Verdon, J.; Labanowski, J.; Sahr, T.; Ferreira, T.; Lacombe, C.; Buchrieser, C.; Berjeaud, J.M.; Héchard, Y. Fatty acid composition modulates sensitivity of Legionella pneumophila to warnericin RK, an antimicrobial peptide. Biochim. Biophys. Acta 2011, 1808, 1146–1153. [Google Scholar]
- Koprivnjak, T.; Peschel, A. Bacterial resistance mechanisms against host defense peptides. Cell. Mol. Life Sci 2011, 68, 2243–2254. [Google Scholar]
- Skiada, A.; Markogiannakis, A.; Plachouras, D.; Daikos, G.L. Adaptive resistance to cationic compounds in Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2011, 37, 187–193. [Google Scholar]
- Richards, S.M.; Strandberg, K.L.; Conroy, M.; Gunn, J.S. Cationic antimicrobial peptides serve as activation signals for the Salmonella Typhimurium PhoPQ and PmrAB regulons in vitro and in vivo. Front. Cell. Infect. Microbiol. 2012, 2. [Google Scholar] [CrossRef]
- Al-Khodar, S.; Kalachikov, S.; Morozova, I.; Price, C.T.; Abu Kwaik, Y. The PmrA/PmrB two-component system of Legionella pneumophila is a global regulator required for intracellular replication within macrophages and protozoa. Infect. Immun 2009, 77, 374–386. [Google Scholar]
- Robey, M.; O’Connell, W.; Cianciotto, N.P. Identification of Legionella pneumophila rcp, a pagP-like gene that confers resistance to cationic antimicrobial peptides and promotes intracellular infection. Infect. Immun 2001, 69, 4276–4286. [Google Scholar]
- Edelstein, P.H. Improved semiselective medium for isolation of Legionella pneumophila from contaminated clinical and environmental specimens. J. Clin. Microbiol 1981, 14, 298–303. [Google Scholar]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem 1987, 166, 368–379. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- White, J.A.; Hart, R.J.; Fry, J.C. An evaluation of the Waters Pico-Tag system for the amino-acid analysis of food materials. J. Automat. Chem 1986, 8, 170–177. [Google Scholar]

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Palusińska-Szysz, M.; Zdybicka-Barabas, A.; Pawlikowska-Pawlęga, B.; Mak, P.; Cytryńska, M. Anti-Legionella dumoffii Activity of Galleria mellonella Defensin and Apolipophorin III. Int. J. Mol. Sci. 2012, 13, 17048-17064. https://doi.org/10.3390/ijms131217048
Palusińska-Szysz M, Zdybicka-Barabas A, Pawlikowska-Pawlęga B, Mak P, Cytryńska M. Anti-Legionella dumoffii Activity of Galleria mellonella Defensin and Apolipophorin III. International Journal of Molecular Sciences. 2012; 13(12):17048-17064. https://doi.org/10.3390/ijms131217048
Chicago/Turabian StylePalusińska-Szysz, Marta, Agnieszka Zdybicka-Barabas, Bożena Pawlikowska-Pawlęga, Pawel Mak, and Małgorzata Cytryńska. 2012. "Anti-Legionella dumoffii Activity of Galleria mellonella Defensin and Apolipophorin III" International Journal of Molecular Sciences 13, no. 12: 17048-17064. https://doi.org/10.3390/ijms131217048