Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
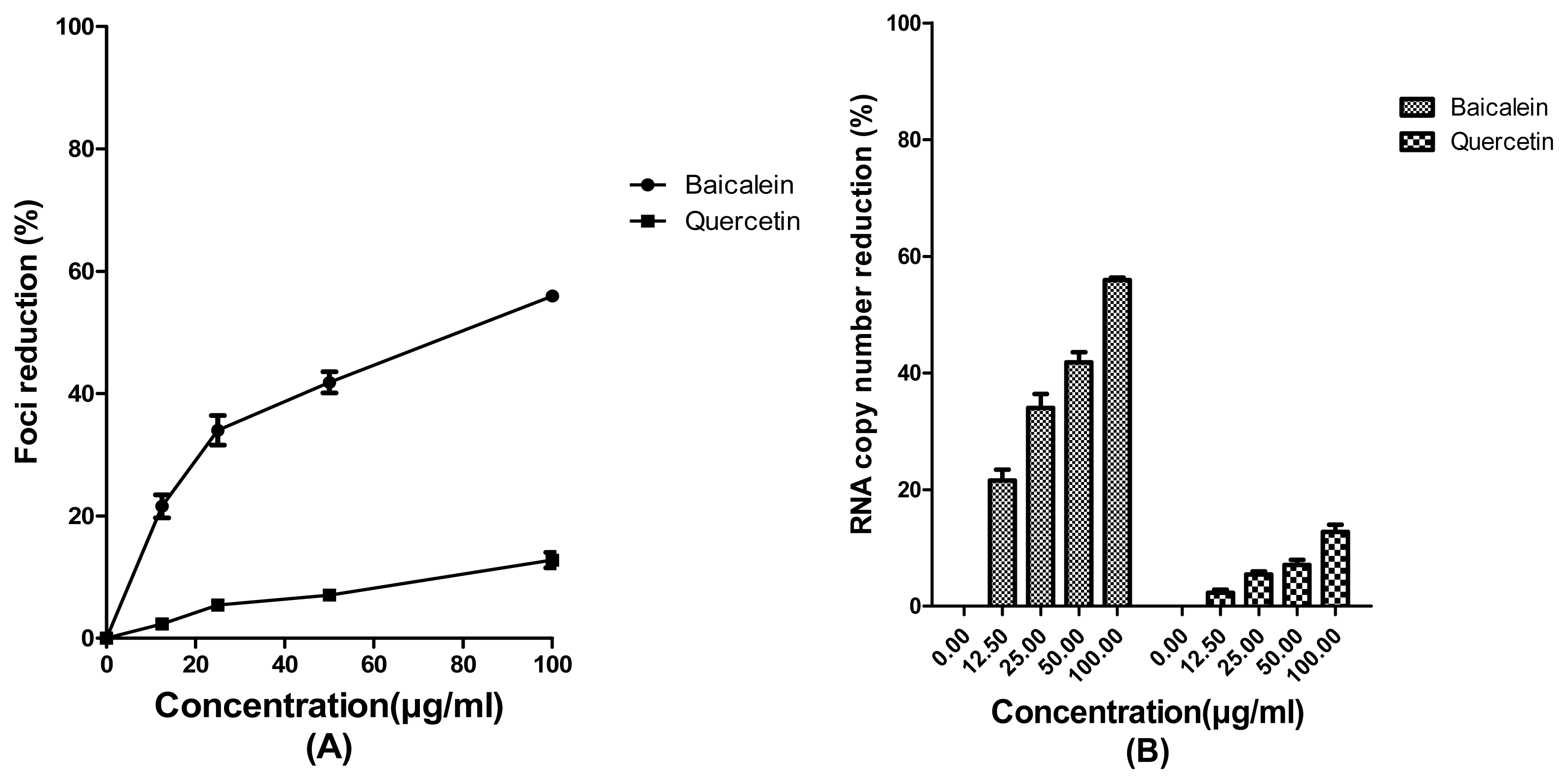
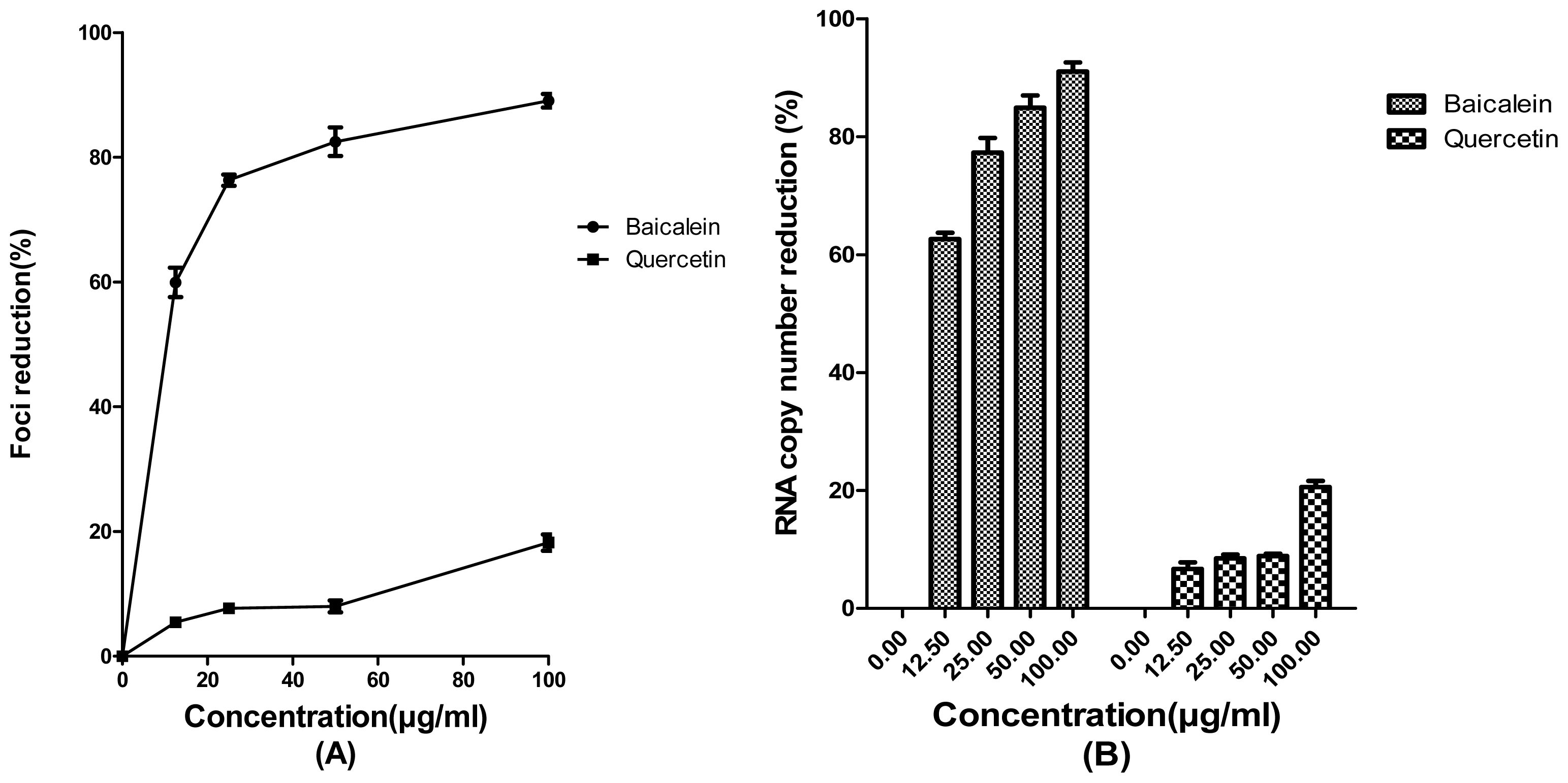
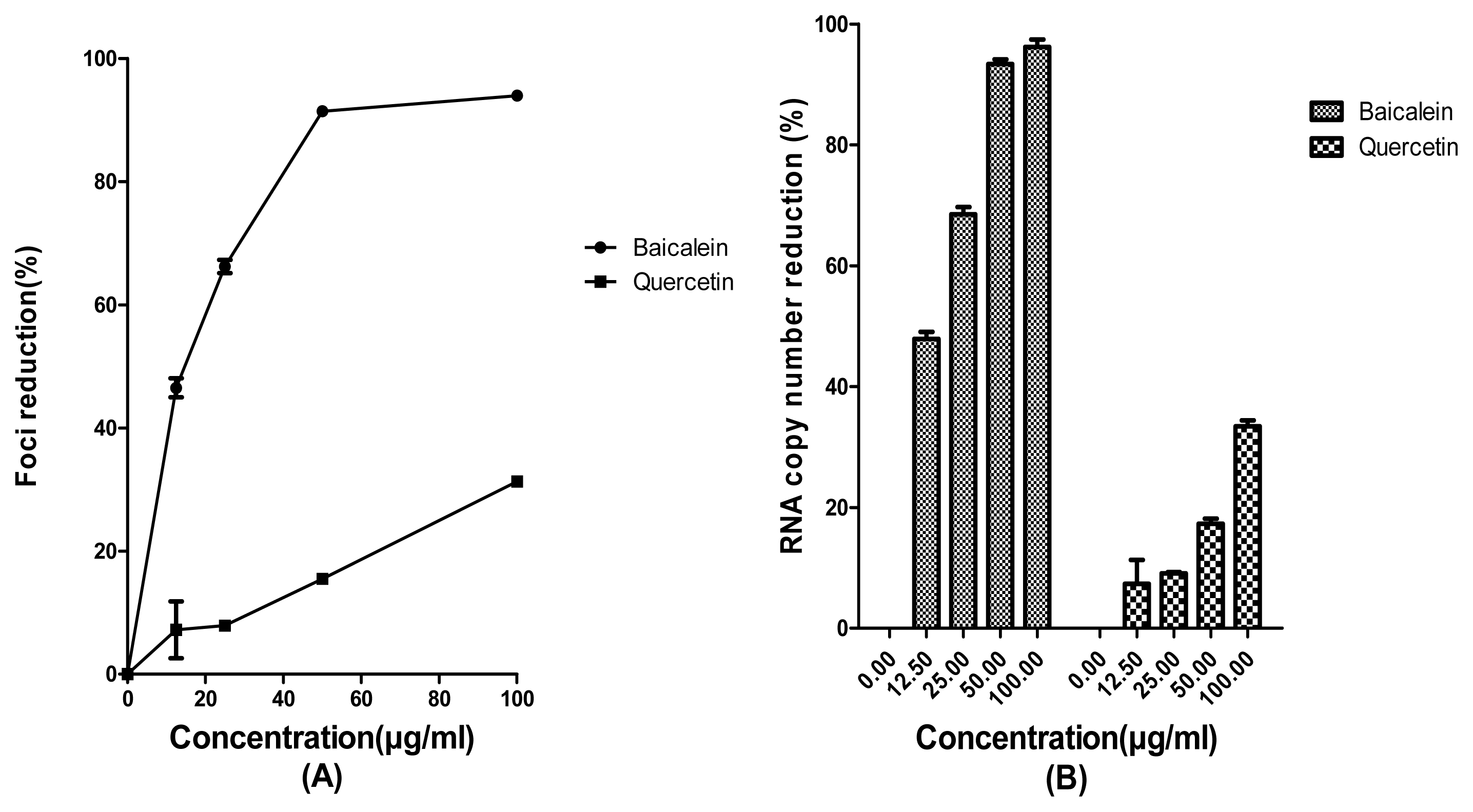
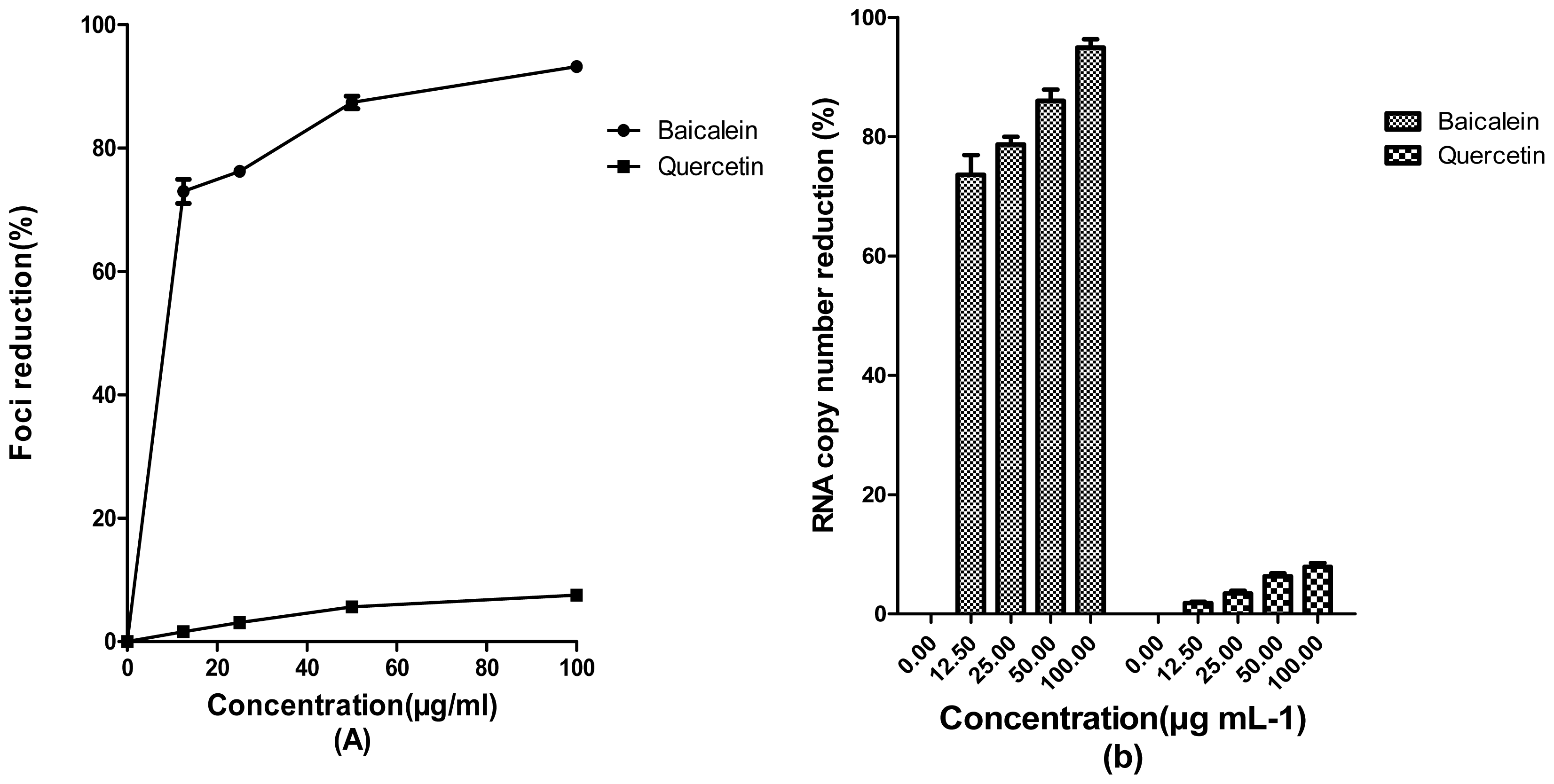
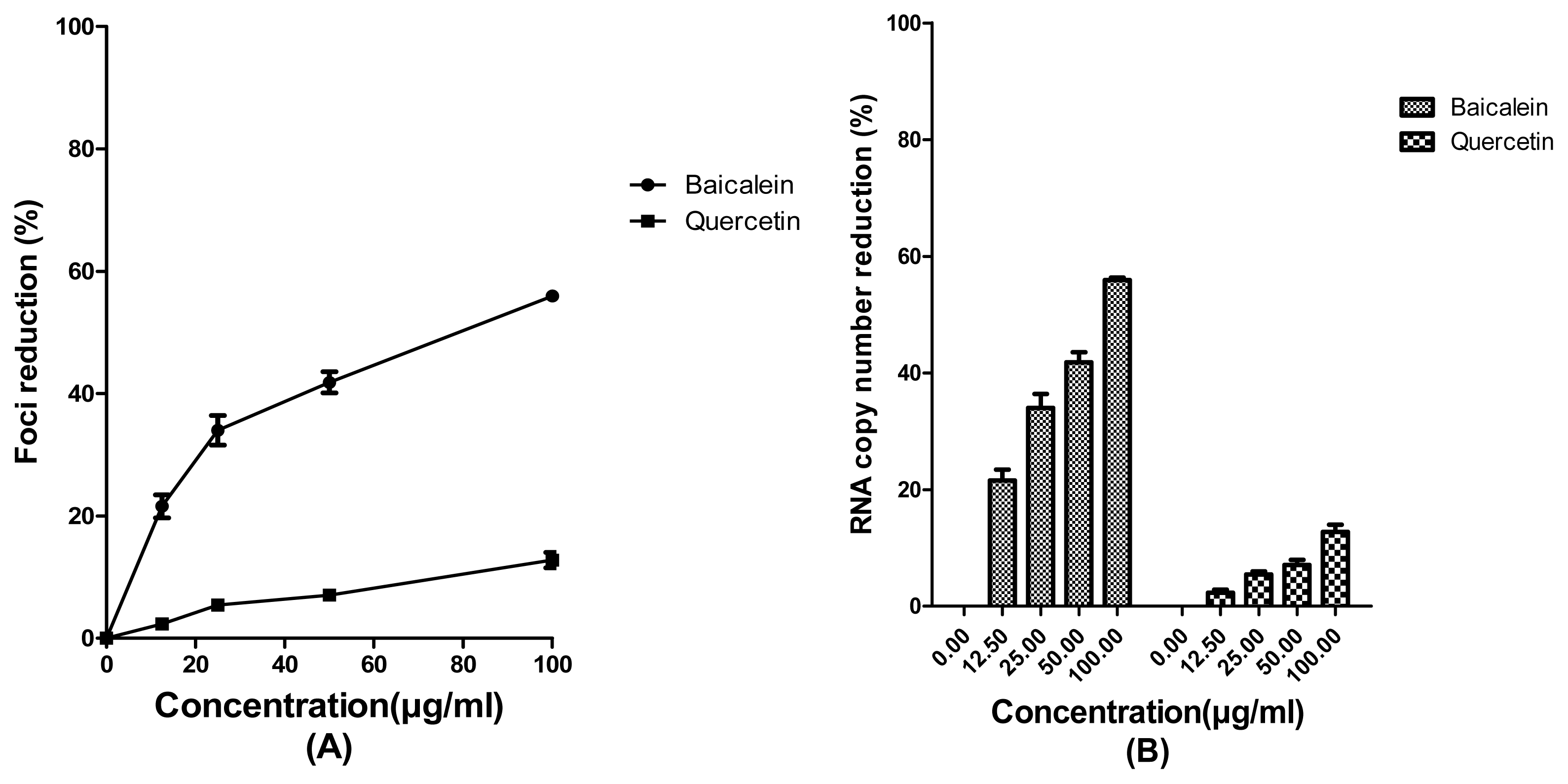
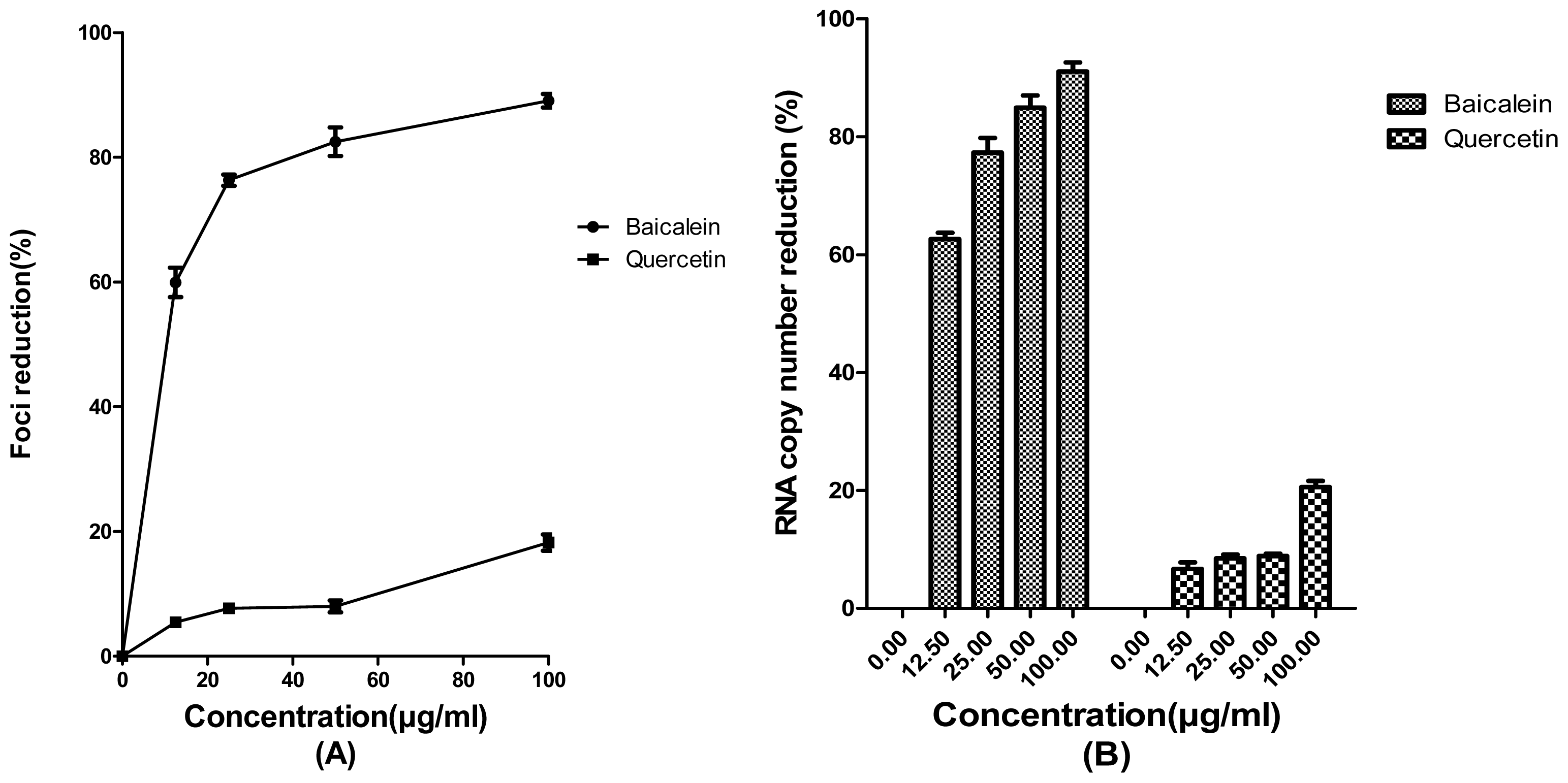
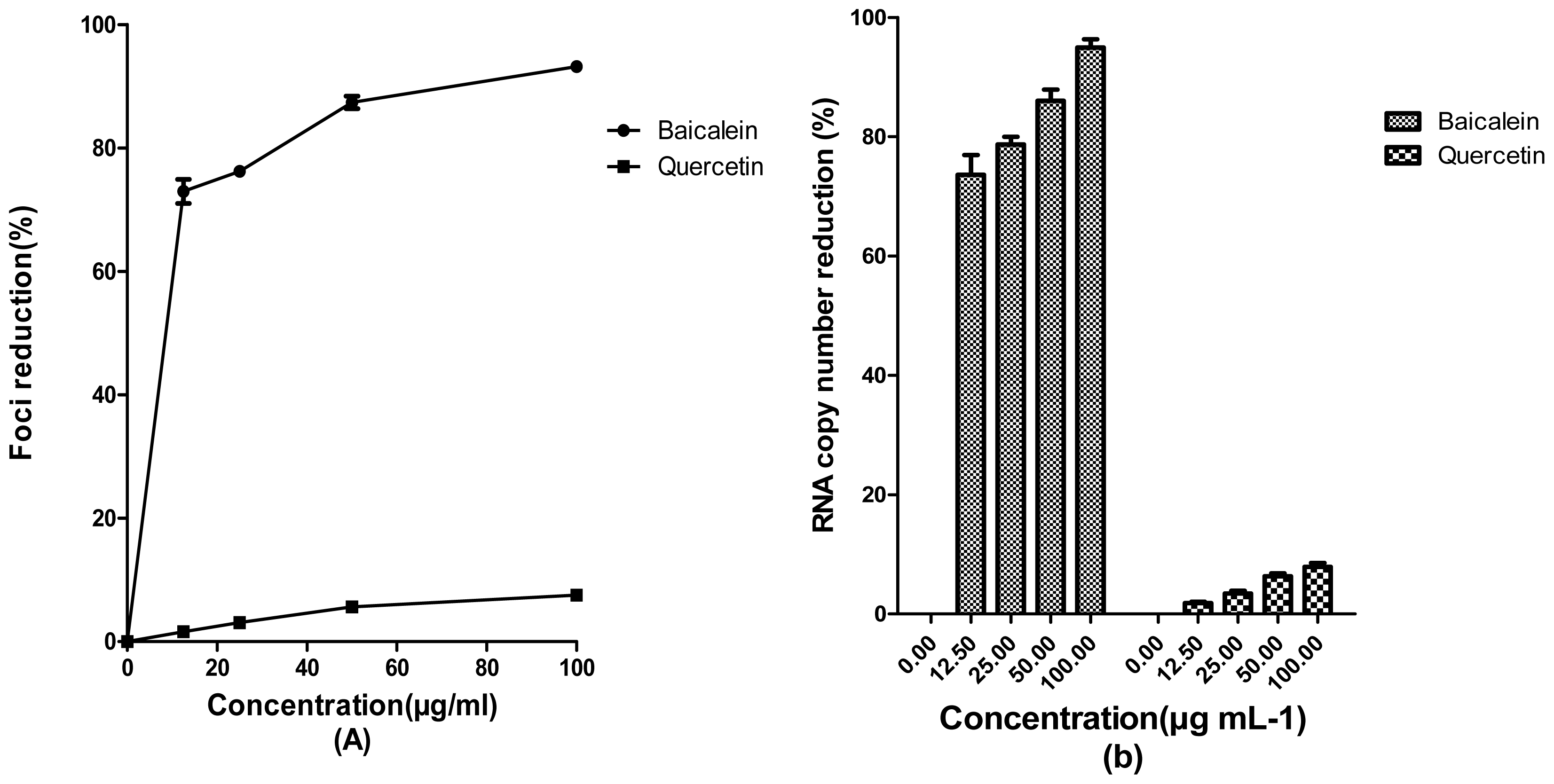
2. Results
2.1. Cytotoxicity of Bioflavonoids
3. Discussion
4. Experimental Section
4.1. Bioflavonoids
4.2. Cell and Virus
4.3. In Vitro Cytotoxicity Assay
4.4. Foci Forming Reduction Assay (FFURA)
4.6. Direct Virucidal Activity Assay
4.7. Quantitative RT-PCR (qRT-PCR)
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Hsiao, N.W.; Chen, J.W.; Yang, T.C.; Orloff, G.M.; Wu, Y.Y.; Lai, C.H.; Lan, Y.C.; Lin, C.W. ISG15 over-expression inhibits replication of the Japanese encephalitis virus in human medulloblastoma cells. Antivir. Res 2010, 85, 504–511. [Google Scholar]
- Paul, W.S.; Moore, P.S.; Karabatsos, N.; Flood, S.P.; Yamada, S.; Jackson, T.; Tsai, T.F. Outbreak of Japanese encephalitis on the island of Saipan, 1990. J. Infect. Dis 1993, 167, 1053–1058. [Google Scholar]
- Wakai, S. Scourge of Japanese encephalitis in southwestern Nepal. Lancet 1998, 351, 759. [Google Scholar]
- Unni, S.K.; Růžek, D.; Chhatbar, C.; Mishra, R.; Johri, M.K.; Singh, S.K. Japanese encephalitis virus: From genome to infectome. Microb. Infect 2011, 13, 312–321. [Google Scholar]
- Solomon, T.; Dung, N.M.; Kneen, R.; Gainsborough, M.; Vaughn, D.W.; Khanh, V.T. Japanese encephalitis. J. Neurol. Neurosurg. Psychiatr 2000, 68, 405–415. [Google Scholar]
- Nazmi, A.; Dutta, K.; Basu, A. Antiviral and neuroprotective role of octaguanidinium dendrimer-conjugated morpholino oligomers in Japanese encephalitis. PLoS Negl. Trop. Dis 2010, 4, e892. [Google Scholar]
- Gubler, D.J. Emerging vector-borne flavivirus diseases: Are vaccines the solution. Expert Rev. Vaccines 2011, 10, 563–565. [Google Scholar]
- Gould, E.A.; Solomon, T.; Mackenzie, J.S. Does antiviral therapy have a role in the control of Japanese encephalitis. Antivir. Res 2008, 78, 140–149. [Google Scholar]
- Chiu, Y.H.; Chan, Y.L.; Li, T.L.; Wu, C.J. Inhibition of Japanese encephalitis virus infection by the sulfated polysaccharide extracts from Ulva lactuca. Mar. Biotechnol 2011, 14, 468–478. [Google Scholar]
- Lee, E.; Pavy, M.; Young, N.; Freeman, C.; Lobigs, M. Antiviral effect of the heparan sulfate mimetic, PI-88, against dengue and encephalitic flaviviruses. Antivir. Res 2006, 69, 31–38. [Google Scholar]
- Zhang, T.; Wu, Z.; Du, J.; Hu, Y.; Liu, L.; Yang, F.; Jin, Q. Anti-Japanese-encephalitis-viral effects of kaempferol and daidzin and their RNA-binding characteristics. PLoS One 2012, 7, e30259. [Google Scholar]
- Hendra, R.; Ahmad, S.; Sukari, A.; Shukor, M.Y.; Oskoueian, E. Flavonoid analyses and antimicrobial activity of various parts of Phaleria macrocarpa (Scheff.) boerl fruit. Int. J. Mol. Sci 2011, 12, 3422–3431. [Google Scholar]
- Dou, J.; Chen, L.; Xu, G.; Zhang, L.; Zhou, H.; Wang, H.; Zhenzhen, S.; Ke, M.; Guo, Q.; Zhou, C. Effects of baicalein on Sendai virus in vivo are linked to serum baicalin and its inhibition of hemagglutinin-neuraminidase. Arch. Virol 2011, 156, 793–801. [Google Scholar]
- Zandi, K.; Lani, R.; Wong, P.F.; Teoh, B.T.; Sam, S.S.; Johari, J.; Mustafa, M.R.; AbuBakar, S. Flavone enhances dengue virus type-2 (NGC strain) infectivity and replication in Vero cells. Molecules 2012, 17, 2437–2445. [Google Scholar]
- Evers, D.L.; Chao, C.F.; Wang, X.; Zhang, Z.; Huong, S.M.; Huang, E.S. Human cytomegalovirus-inhibitory flavonoids: Studies on antiviral activity and mechanism of action. Antivir. Res 2005, 68, 124–134. [Google Scholar]
- Lyu, S.Y.; Rhim, J.Y.; Park, W.B. Antiherpetic activities of flavonoids against herpes simplex virus type 1 (HSV-1) and type 2 (HSV-2) in vitro. Arch. Pharm. Res 2005, 28, 1293–1301. [Google Scholar]
- Xu, G.; Dou, L.; Guo, Q.; Zhou, C. Inhibitory effects of baicalein on the influenza virus in vivo is determined by baicalein in the serum. Biol. Pharm. Bull 2010, 33, 238–243. [Google Scholar]
- Chiang, L.C.; Chiang, W.; Liu, M.C.; Lin, C.C. In vitro antiviral activities of Caesalpinia pulcherrima and its related flavonoids. J. Antimicrob. Chemother 2003, 52, 194–198. [Google Scholar]
- Choi, H.J.; Kim, J.H.; Lee, C.H.; Ahn, Y.J.; Song, J.H.; Baek, S.H.; Kwon, D.H. Antiviral activity of quercetin 7-rhamnoside against porcine epidemic diarrhea virus. Antivir. Res 2009, 81, 77–81. [Google Scholar]
- Kim, Y.; Narayanan, S.; Chang, K.O. Inhibition of influenza virus replication by plant derived isoquercetin. Antivir. Res 2010, 88, 227–235. [Google Scholar]
- Zandi, K.; Teoh, B.T.; Sam, S.S.; Wong, P.F.; Mustafa, M.R.; Abubakar, S. Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virol. J 2011, 8, 560. [Google Scholar]
- Shabrova, E.V.; Tarnopolsky, O.; Singh, A.P.; Plutzky, J.; Vorsa, N.; Quadro, L. Insights into the molecular mechanisms of the anti-atherogenic actions of flavonoids in normal and obese mice. PLoS One 2011, 6, e24634. [Google Scholar]
- Chen, V.; Staub, R.E.; Baggett, S.; Chimmani, R.; Tagliaferri, M.; Cohen, I.; Shtivelman, E. Identification and analysis of the active phytochemicals from the anti-cancer botanical extract bezielle. PLoS One 2012, 7, e30107. [Google Scholar]
- Chen, L.; Dou, J.; Su, Z.; Zhou, H.; Wang, H.; Zhou, W.; Guo, Q.; Zhou, C. Synergistic activity of baicalein with ribavirin against influenza A (H1N1) virus infections in cell culture and in mice. Antivir. Res 2011, 91, 314–320. [Google Scholar]
- Ahn, H.C.; Lee, S.Y.; Kim, J.W.; Son, W.S.; Shin, C.G.; Lee, B.J. Binding aspects of baicalein to HIV-1 integrase. Mol. Cells 2001, 31, 127–130. [Google Scholar]
- Kitamura, K.; Honda, M.; Yoshizaki, H.; Yamamoto, S.; Nakane, H.; Fukushima, M.; Ono, K.; Tokunaga, T. Baicalin, an inhibitor of HIV-1 production in vitro. Antivir. Res 1998, 37, 131–140. [Google Scholar]
- Bachmetov, L.; Gal-Tanamy, M.; Shapira, A.; Vorobeychik, M.; Giterman-Galam, T.; Sathiyamoorthy, P.; Golan-Goldhirsh, A.; Benhar, I.; Tur-Kaspa, R.; Zemel, R. Suppression of hepatitis C virus by the flavonoid quercetin is mediated by inhibition of NS3 protease activity. J. Viral. Hepat 2012, 19, e81–e88. [Google Scholar]
- Santhosh, S.R.; Parida, M.M.; Dash, P.K.; Pateriya, A.; Pattnaik, B.; Pradhan, H.K.; Tripathi, N.K.; Ambuj, S.; Gupta, N.; Saxena, P.; et al. Development and evaluation of SYBR Green I-based one-step real-time RT-PCR assay for detection and quantification of Chikungunya virus. J. Clin. Virol 2007, 39, 188–193. [Google Scholar]
- Park, S.I.; Park, D.H.; Saif, L.J.; Jeong, Y.J.; Shin, D.J.; Chun, Y.H.; Park, S.J.; Kim, H.J.; Hosmillo, M.; Kwon, H.J.; et al. Development of SYBR Green real-time RT-PCR for rapid detection, quantitation and diagnosis of unclassified bovine enteric calicivirus. J. Virol. Methods 2009, 159, 64–68. [Google Scholar]

© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Johari, J.; Kianmehr, A.; Mustafa, M.R.; Abubakar, S.; Zandi, K. Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. Int. J. Mol. Sci. 2012, 13, 16785-16795. https://doi.org/10.3390/ijms131216785
Johari J, Kianmehr A, Mustafa MR, Abubakar S, Zandi K. Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. International Journal of Molecular Sciences. 2012; 13(12):16785-16795. https://doi.org/10.3390/ijms131216785
Chicago/Turabian StyleJohari, Jefree, Aynaz Kianmehr, Mohd Rais Mustafa, Sazaly Abubakar, and Keivan Zandi. 2012. "Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus" International Journal of Molecular Sciences 13, no. 12: 16785-16795. https://doi.org/10.3390/ijms131216785
APA StyleJohari, J., Kianmehr, A., Mustafa, M. R., Abubakar, S., & Zandi, K. (2012). Antiviral Activity of Baicalein and Quercetin against the Japanese Encephalitis Virus. International Journal of Molecular Sciences, 13(12), 16785-16795. https://doi.org/10.3390/ijms131216785