Sensing and Responding to UV-A in Cyanobacteria


{kind=link}
{kind=link}
{kind=link}
Abstract
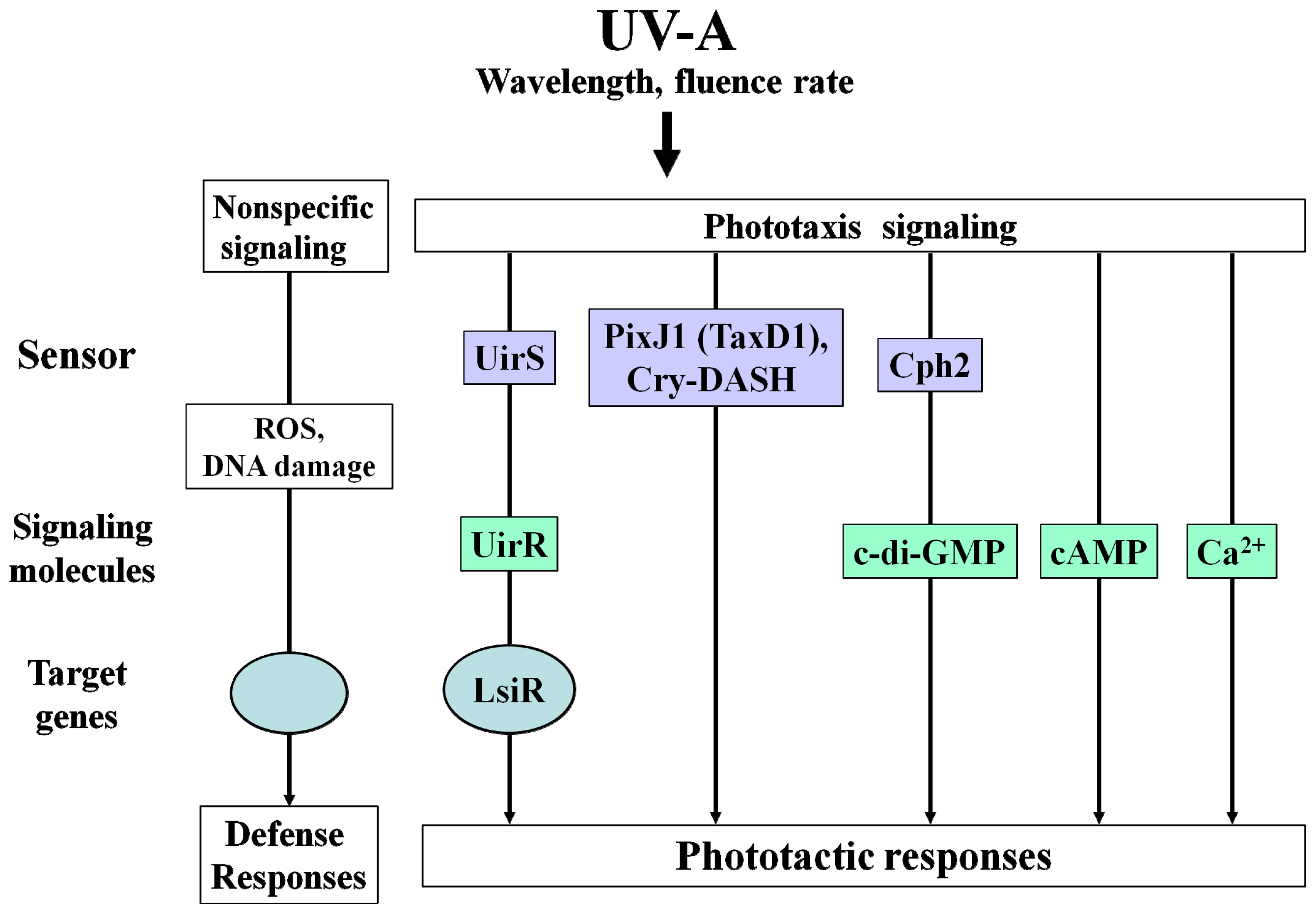
:1. Introduction
2. UV-Mediated Signaling Pathways in Cyanobacteria
2.1. High Fluence Rate UV-A Dependent Phototaxis Signaling Pathway
2.2. Low Fluence Rate UV-A Dependent Phototaxis Signaling Pathway
2.3. Second Messengers in Cellular Response to UV Radiation
3. Role of Pterin in Responses to UV Radiation
3.1. Cyanobacterial Pteridines
3.2. Evidences for the Involvement of Pterin in UV-Perception
3.3. The Role of Pterin in the Regulation of Cyanobacterial Phototaxis
3.4. Cyanobacterial Cryptochromes
3.5. Involvement of Cryptochrome in the Regulation of Phototactic Response to UV-A
4. Possible Role of Pterin as a UV-A-Protecting Compound
5. UV Stress and Programmed Cell Death in Cyanobacteria
6. Concluding Remarks
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
Abbreviations
| UV | Ultraviolet |
| ROS | Reactive Oxygen Species |
| Syn6803 | Synechocystis sp. PCC 6803 |
| RuBisCO | Ribulose 1,5-Bisphosphate Carboxylase/Oxygenase |
| CPDs | Cyclobutane Pyrimidine Dimers |
| CBCRs | Cyanobacteriochromes |
| PCB | Phycocyanobilin |
| UirS | UV intensity response Sensor |
| UirR | UV intensity response Regulator |
| LsiR | Light and stress integrating response Regulator |
| HK | Histidine Kinase |
| ETR | Ethylene Receptor |
| NAC | N-Acetyl Cysteine |
| c-di-GMP | 3′,5′-cyclic diguanylic acid |
| cAMP | 3′,5′-cyclic adenosine monophosphate |
| SYCRP | Synechocystis sp. cAMP Receptor Protein |
| PgtA | Pteridine glycosyltransferase |
| BGluT | UDP-glucose: BH4 α-Glucosyltransferase |
| BH4 | Tetrahydrobiopterin |
| FAD | Flavin Adenine Dinucleotide |
| GTPCH | GTP Cyclohydrolase I |
| NAS | N-Acetylserotonin |
| SR | Sepiapterin Reductase |
| DAHP | 2,4-Diamino-6-hydroxypyrimidine |
| MAA | Mycosporine-like Amino Acid |
| BLUF | Sensor of Blue Light using FAD |
| Cry-DASH | Cryptochrome-Drosophila, Arabidopsis, Synechocystis, Human |
| PHR | Photolyase-Homologous Region |
| MTHF | 5,10-Methenyl tetrahydrofolate |
| 8-HDF | 8-Hydroxy-5-deazaflavin |
| PCD | Programmed Cell Death |
| MCAs | Metacaspases |
References
- Hargreaves, A.; Taiwo, F.A.; Duggan, O.; Kirk, S.H.; Ahmad, S.I. Near-ultraviolet photolysis of beta-phenylpyruvic acid generates free radicals and results in DNA damage. J. Photochem. Photobiol. B 2007, 89, 110–116. [Google Scholar]
- Vincent, W.F.; Roy, S. Solar ultraviolet-B radiation and aquatic primary production: Damage, protection, and recovery. Environ. Rev 1993, 1, 1–12. [Google Scholar]
- Caldwell, M.M.; Bornman, J.F.; Ballare, C.L.; Flint, S.D.; Kulandaivelu, G. Terrestrial ecosystems, increased solar ultraviolet radiation, and interactions with other climate change factors. Photochem. Photobiol. Sci 2007, 6, 252–266. [Google Scholar]
- Sinha, R.P.; Klisch, M.; Gröniger, A.; Häder, D.-P. Responses of aquatic algae and cyanobacteria to solar UV-B. Plant Ecol 2001, 154, 219–236. [Google Scholar]
- Rozema, J.; Bjorn, L.O.; Bornman, J.F.; Gaberscik, A.; Häder, D.-P.; Trost, T.; Germ, M.; Klisch, M.; Groniger, A.; Sinha, R.P.; et al. The role of UV-B radiation in aquatic and terrestrial ecosystems—An experimental and functional analysis of the evolution of UV-absorbing compounds. J. Photochem. Photobiol. B 2002, 66, 2–12. [Google Scholar]
- Singh, S.P.; Häder, D.-P.; Sinha, R.P. Cyanobacteria and ultraviolet radiation (UVR) stress: Mitigation strategies. Ageing Res. Rev 2010, 9, 79–90. [Google Scholar]
- Castenholz, R.W.; Garcia-Pichel, F. Cyanobacterial Responses to UV-Radiation. In Ecology of Cyanobacteria II: Their Diversity in Time and Space; Whitton, B.A., Ed.; Springer press: Dordrecht, The Netherlands, 2012; pp. 481–499. [Google Scholar]
- Holzinger, A.; Lutz, C. Algae and UV irradiation: Effects on ultrastructure and related metabolic functions. Micron 2006, 37, 190–207. [Google Scholar]
- Xue, L.; Zhang, Y.; Zhang, T.; An, L.; Wang, X. Effects of enhanced ultraviolet-B radiation on algae and cyanobacteria. Crit. Rev. Microbiol 2005, 31, 79–89. [Google Scholar]
- Vincent, W.F.; Neale, P.J. Mechanisms of UV Damage to Aquatic Organisms. In The Effects of UV Radiation on Marine Ecosystems; de Mora, S.J., Demers, S., Vernet, M., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 149–176. [Google Scholar]
- He, Y.Y.; Häder, D.-P. Reactive oxygen species and UV-B: Effect on cyanobacteria. Photochem. Photobiol. Sci 2002, 1, 729–736. [Google Scholar]
- Lao, K.; Glazer, A.N. Ultraviolet-B photodestruction of a light-harvesting complex. Proc. Natl. Acad. Sci. USA 1996, 93, 5258–5263. [Google Scholar]
- Vass, I.; Kirilovsky, D.; Etienne, A.L. UV-B radiation-induced donor- and acceptor-side modifications of photosystem II in the cyanobacterium Synechocystis sp. PCC 6803. Biochemistry 1999, 38, 12786–12794. [Google Scholar]
- Singh, V.P.; Srivastava, P.K.; Prasad, S.M. Differential effect of UV-B radiation on growth, oxidative stress and ascorbate-glutathione cycle in two cyanobacteria under copper toxicity. Plant Physiol. Biochem 2012, 61, 61–70. [Google Scholar]
- Sinha, R.P.; Kumar, H.D.; Kumar, A.; Häder, D.-P. Effects of UV-B irradiation on growth, survival, pigmentation and nitrogen metabolism enzymes in cyanobacteria. Acta Protozool 1995, 34, 187–192. [Google Scholar]
- Donkor, V.A.; Häder, D.-P. Effects of ultraviolet irradiation on photosynthetic pigments in some filamentous cyanobacteria. Aquat. Microb. Ecol 1996, 11, 143–149. [Google Scholar]
- Kumar, A.; Sinha, R.P.; Häder, D.-P. Effect of UV-B on enzymes of nitrogen metabolism in the cynobacterium Nostoc calcicola. J. Phycol 1996, 148, 86–91. [Google Scholar]
- Sinha, R.P.; Singh, N.; Kumar, A.; Kumar, H.D.; Häder, M.; Häder, D.-P. Effects of UV irradiation on certain physiological and biochemical processes in cyanobacteria. J. Photochem. Photobiol. B 1996, 32, 107–113. [Google Scholar]
- Kumar, A.; Tyagi, M.B.; Singh, N.; Tyagi, R.; Jha, P.N.; Sinha, R.P.; Häder, D.-P. Role of white light in reversing UV-B-mediated effects in the N2-fixing cyanobacterium Anabaena BT2. J. Photochem. Photobiol. B 2003, 71, 35–42. [Google Scholar]
- Sinha, R.P.; Singh, N.; Kumar, A.; Kumar, H.D.; Häder, D.-P. Impacts of ultraviolet-B irradiation on nitrogen-fixing cyanobacteria of rice paddy fields. J. Plant Physiol 1997, 150, 188–193. [Google Scholar]
- Donkor, V.A.; Häder, D.-P. Effects of solar and ultraviolet radiation on motility, photomovement and pigmentation in filamentous, gliding cyanobacterium. FEMS Microbiol. Ecol 1991, 86, 159–168. [Google Scholar]
- Gao, K.; Yu, H.; Brown, M.T. Solar PAR and UV radiation affects the physiology and morphology of the cyanobacterium Anabaena sp. PCC 7120. J. Photochem. Photobiol. B 2007, 89, 117–124. [Google Scholar]
- Vass, I.; Turcsanyi, E.; Touloupakis, E.; Ghanotakis, D.; Petrouleas, V. The mechanism of UV-A radiation-induced inhibition of photosystem II electron transport studied by EPR and chlorophyll fluorescence. Biochemistry 2002, 41, 10200–10208. [Google Scholar]
- Cullen, J.J.; Neale, P.J.; Lesser, M.P. Biological weighting function for the inhibition of phytoplankton photosynthesis by ultraviolet radiation. Science 1992, 258, 646–650. [Google Scholar]
- Bebout, B.M.; Garcia-Pichel, F. UV B-Induced Vertical Migrations of Cyanobacteria in a Microbial Mat. Appl. Environ. Microbiol 1995, 61, 4215–4222. [Google Scholar]
- Sinha, R.P.; Häder, D.-P. UV-protectants in cyanobacteria. Plant Sci 2008, 174, 278–289. [Google Scholar]
- Gao, Q.; Garcia-Pichel, F. Microbial ultraviolet sunscreens. Nat. Rev. Microbiol 2011, 9, 791–802. [Google Scholar]
- Wang, G.; Chen, K.; Chen, L.; Hu, C.; Zhang, D.; Liu, Y. The involvement of the antioxidant system in protection of desert cyanobacterium Nostoc sp. against UV-B radiation and the effects of exogenous antioxidants. Ecotoxicol. Environ. Saf 2008, 69, 150–157. [Google Scholar]
- Sinha, R.P.; Häder, D.-P. UV-induced DNA damage and repair: A review. Photochem. Photobiol. Sci 2002, 1, 225–236. [Google Scholar]
- Sass, L.; Spetea, C.; Mate, Z.; Nagy, F.; Vass, I. Repair of UV-B-induced damage of photosystem II via de novo synthesis of the D1 and D2 reaction centre subunits in Synechocystis sp. PCC 6803. Photosynth. Res 1997, 54, 55–62. [Google Scholar]
- Richa Sinha, R.P. UV-mediated stress and its mitigation in cyanobacteria. Int. J. Plant Anim. Environ. Sci. 2011, 1, 155–166. [Google Scholar]
- Ehling-Schulz, M.; Scherer, S. UV protection in cyanobacteria. Eur. J. Phycol 1999, 34, 329–338. [Google Scholar]
- Castenholz, R.W.; Garcia-Pichel, F. Cyanobacterial Responses to UV-Radiation. In In the Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Kluwer Academic Publishers: Dordrecht, Netherlands, 2000; pp. 591–611. [Google Scholar]
- Sgarbossa, A.; Checcucci, G.; Lenci, F. Photoreception and photomovements of microorganisms. Photochem. Photobiol. Sci 2002, 1, 459–467. [Google Scholar]
- Häder, D.-P. Photosensory behavior in procaryotes. Microbiol. Rev 1987, 51, 1–21. [Google Scholar]
- Lebert, M. Phototaxis of Euglena gracilis—Flavins and pterins. In Photomovement; Häder, D.-P., Lebert, M., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2001; pp. 297–341. [Google Scholar]
- Choi, J.S.; Chung, Y.H.; Moon, Y.J.; Kim, C.; Watanabe, M.; Song, P.S.; Joe, C.O.; Bogorad, L.; Park, Y.M. Photomovement of the gliding cyanobacterium Synechocystis sp. PCC 6803. Photochem. Photobiol 1999, 70, 95–102. [Google Scholar]
- Bhaya, D.; Bianco, N.R.; Bryant, D.; Grossman, A. Type IV pilus biogenesis and motility in the cyanobacterium Synechocystis sp. PCC 6803. Mol. Microbiol 2000, 37, 941–951. [Google Scholar]
- Chung, Y.H.; Cho, M.S.; Moon, Y.J.; Choi, J.S.; Yoo, Y.C.; Park, Y.I.; Lee, K.M.; Kang, K.W.; Park, Y.M. ctr1, a gene involved in a signal transduction pathway of the gliding motility in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett 2001, 492, 33–38. [Google Scholar]
- Yoshihara, S.; Geng, X.; Okamoto, S.; Yura, K.; Murata, T.; Go, M.; Ohmori, M.; Ikeuchi, M. Mutational analysis of genes involved in pilus structure, motility and transformation competency in the unicellular motile cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol 2001, 42, 63–73. [Google Scholar]
- Mullineaux, C.W. How do cyanobacteria sense and respond to light? Mol. Microbiol 2001, 41, 965–971. [Google Scholar]
- Montgomery, B.L. Sensing the light: Photoreceptive systems and signal transduction in cyanobacteria. Mol. Microbiol 2007, 64, 16–27. [Google Scholar]
- Ikeuchi, M.; Ishizuka, T. Cyanobacteriochromes: A new superfamily of tetrapyrrole-binding photoreceptors in cyanobacteria. Photochem. Photobiol. Sci 2008, 7, 1159–1167. [Google Scholar]
- Yoshihara, S.; Katayama, M.; Geng, X.; Ikeuchi, M. Cyanobacterial phytochrome-like PixJ1 holoprotein shows novel reversible photoconversion between blue- and green-absorbing forms. Plant Cell Physiol 2004, 45, 1729–1737. [Google Scholar]
- Ishizuka, T.; Shimada, T.; Okajima, K.; Yoshihara, S.; Ochiai, Y.; Katayama, M.; Ikeuchi, M. Characterization of cyanobacteriochrome TePixJ from a thermophilic cyanobacterium Thermosynechococcus elongatus strain BP-1. Plant Cell Physiol 2006, 47, 1251–1261. [Google Scholar]
- Rockwell, N.C.; Njuguna, S.L.; Roberts, L.; Castillo, E.; Parson, V.L.; Dwojak, S.; Lagarias, J.C.; Spiller, S.C. A second conserved GAF domain cysteine is required for the blue/green photoreversibility of cyanobacteriochrome Tlr0924 from Thermosynechococcus elongatus. Biochemistry 2008, 47, 7304–7316. [Google Scholar]
- Terauchi, K.; Montgomery, B.L.; Grossman, A.R.; Lagarias, J.C.; Kehoe, D.M. RcaE is a complementary chromatic adaptation photoreceptor required for green and red light responsiveness. Mol. Microbiol 2004, 51, 567–577. [Google Scholar]
- Hirose, Y.; Shimada, T.; Narikawa, R.; Katayama, M.; Ikeuchi, M. Cyanobacteriochrome CcaS is the green light receptor that induces the expression of phycobilisome linker protein. Proc. Natl. Acad. Sci. USA 2008, 105, 9528–9533. [Google Scholar]
- Hirose, Y.; Narikawa, R.; Katayama, M.; Ikeuchi, M. Cyanobacteriochrome CcaS regulates phycoerythrin accumulation in Nostoc punctiforme, a group II chromatic adapter. Proc. Natl. Acad. Sci. USA 2010, 107, 8854–8859. [Google Scholar]
- Narikawa, R.; Fukushima, Y.; Ishizuka, T.; Itoh, S.; Ikeuchi, M. A novel photoactive GAF domain of cyanobacteriochrome AnPixJ that shows reversible green/red photoconversion. J. Mol. Biol 2008, 380, 844–855. [Google Scholar]
- Rockwell, N.C.; Martin, S.S.; Feoktistova, K.; Lagarias, J.C. Diverse two-cysteine photocycles in phytochromes and cyanobacteriochromes. Proc. Natl. Acad. Sci. USA 2011, 108, 11854–11859. [Google Scholar]
- Narikawa, R.; Kohchi, T.; Ikeuchi, M. Characterization of the photoactive GAF domain of the CikA homolog (SyCikA, Slr1969) of the cyanobacterium Synechocystis sp. PCC 6803. Photochem. Photobiol. Sci 2008, 7, 1253–1259. [Google Scholar]
- Kehoe, D.M.; Grossman, A.R. Similarity of a chromatic adaptation sensor to phytochrome and ethylene receptors. Science 1996, 273, 1409–1412. [Google Scholar]
- Yoshihara, S.; Suzuki, F.; Fujita, H.; Geng, X.X.; Ikeuchi, M. Novel putative photoreceptor and regulatory genes required for the positive phototactic movement of the unicellular motile cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol 2000, 41, 1299–1304. [Google Scholar]
- Bhaya, D.; Takahashi, A.; Grossman, A.R. Light regulation of type IV pilus-dependent motility by chemosensor-like elements in Synechocystis PCC 6803. Proc. Natl. Acad. Sci. USA 2001, 98, 7540–7545. [Google Scholar]
- Wilde, A.; Fiedler, B.; Borner, T. The cyanobacterial phytochrome Cph2 inhibits phototaxis towards blue light. Mol. Microbiol 2002, 44, 981–988. [Google Scholar]
- Moon, Y.J.; Kim, S.Y.; Jung, K.H.; Choi, J.S.; Park, Y.M.; Chung, Y.H. Cyanobacterial phytochrome Cph2 is a negative regulator in phototaxis toward UV-A. FEBS Lett 2011, 585, 335–340. [Google Scholar]
- Moon, Y.J.; Lee, E.M.; Park, Y.M.; Park, Y.S.; Chung, W.I.; Chung, Y.H. The role of cyanopterin in UV/blue light signal transduction of cyanobacterium Synechocystis sp. PCC 6803 phototaxis. Plant Cell Physiol 2010, 51, 969–980. [Google Scholar]
- Moon, Y.J.; Kim, S.J.; Park, Y.M.; Chung, Y.H. Sensing UV/blue: Pterin as a UV-A absorbing chromophore of cryptochrome. Plant Signal. Behav 2010, 5, 1127–1130. [Google Scholar]
- Song, J.Y.; Cho, H.S.; Cho, J.I.; Jeon, J.S.; Lagarias, J.C.; Park, Y.I. Near-UV cyanobacteriochrome signaling system elicits negative phototaxis in the cyanobacterium Synechocystis sp. PCC 6803. Proc. Natl. Acad. Sci. USA 2011, 108, 10780–10785. [Google Scholar]
- He, Y.Y.; Klisch, M.; Häder, D.-P. Adaptation of cyanobacteria to UV-B stress correlated with oxidative stress and oxidative damage. Photochem. Photobiol 2002, 76, 188–196. [Google Scholar]
- He, Y.Y.; Häder, D.-P. Involvement of reactive oxygen species in the UV-B damage to the cyanobacterium Anabaena sp. J. Photochem. Photobiol. B 2002, 66, 73–80. [Google Scholar]
- Shibata, H.; Noda, T.; Ogura, Y.; Suginaka, K.; Matsui, Y.; Ozoe, Y.; Sawa, Y.; Kono, Y. A soluble-form of pro-oxidant lumazine isolated from cyanobacterial cells generates superoxide anion under near-UV irradiation. Biochim. Biophys. Acta 1996, 1274, 129–134. [Google Scholar]
- Frohnmeyer, H.; Staiger, D. Ultraviolet-B radiation-mediated responses in plants. Balancing damage and protection. Plant Physiol 2003, 133, 1420–1428. [Google Scholar]
- Jenkins, G.I. Signal transduction in responses to UV-B radiation. Annu. Rev. Plant Biol 2009, 60, 407–431. [Google Scholar]
- Rockwell, N.C.; Lagarias, J.C. A brief history of phytochromes. Chemphyschem 2010, 11, 1172–1280. [Google Scholar]
- Rodriguez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A copper cofactor for the ethylene receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar]
- Ulijasz, A.T.; Cornilescu, G.; von Stetten, D.; Cornilescu, C.; Velazquez Escobar, F.; Zhang, J.; Stankey, R.J.; Rivera, M.; Hildebrandt, P.; Vierstra, R.D. Cyanochromes are blue/green light photoreversible photoreceptors defined by a stable double cysteine linkage to a phycoviolobilin-type chromophore. J. Biol. Chem 2009, 284, 29757–29772. [Google Scholar]
- Wang, H.L.; Postier, B.L.; Burnap, R.L. Alterations in global patterns of gene expression in Synechocystis sp. PCC 6803 in response to inorganic carbon limitation and the inactivation of ndhR, a LysR family regulator. J. Biol. Chem 2004, 279, 5739–5751. [Google Scholar]
- Singh, A.K.; McIntyre, L.M.; Sherman, L.A. Microarray analysis of the genome-wide response to iron deficiency and iron reconstitution in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol 2003, 132, 1825–1839. [Google Scholar]
- Li, H.; Singh, A.K.; McIntyre, L.M.; Sherman, L.A. Differential gene expression in response to hydrogen peroxide and the putative PerR regulon of Synechocystis sp. strain PCC 6803. J. Bacteriol 2004, 186, 3331–3345. [Google Scholar]
- Singh, A.K.; Elvitigala, T.; Bhattacharyya-Pakrasi, M.; Aurora, R.; Ghosh, B.; Pakrasi, H.B. Integration of carbon and nitrogen metabolism with energy production is crucial to light acclimation in the cyanobacterium Synechocystis. Plant Physiol 2008, 148, 467–478. [Google Scholar]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol 2004, 55, 373–399. [Google Scholar]
- Hancock, J.; Desikan, R.; Harrison, J.; Bright, J.; Hooley, R.; Neill, S. Doing the unexpected: Proteins involved in hydrogen peroxide perception. J. Exp. Bot 2006, 57, 1711–1718. [Google Scholar]
- Desikan, R.; Hancock, J.T.; Bright, J.; Harrison, J.; Weir, I.; Hooley, R.; Neill, S.J. A role for ETR1 in hydrogen peroxide signaling in stomatal guard cells. Plant Physiol 2005, 137, 831–834. [Google Scholar]
- Desikan, R.; Last, K.; Harrett-Williams, R.; Tagliavia, C.; Harter, K.; Hooley, R.; Hancock, J.T.; Neill, S.J. Ethylene-induced stomatal closure in Arabidopsis occurs via AtrbohF-mediated hydrogen peroxide synthesis. Plant J 2006, 47, 907–916. [Google Scholar]
- Cooper, C.E.; Patel, R.P.; Brookes, P.S.; Darley-Usmar, V.M. Nanotransducers in cellular redox signaling: Modification of thiols by reactive oxygen and nitrogen species. Trends Biochem. Sci 2002, 27, 489–492. [Google Scholar]
- Grefen, C.; Harter, K. Plant two-component systems: Principles, functions, complexity and cross talk. Planta 2004, 219, 733–742. [Google Scholar]
- Sweere, U.; Eichenberg, K.; Lohrmann, J.; Mira-Rodado, V.; Baurle, I.; Kudla, J.; Nagy, F.; Schafer, E.; Harter, K. Interaction of the response regulator ARR4 with phytochrome B in modulating red light signaling. Science 2001, 294, 1108–1111. [Google Scholar]
- Mira-Rodado, V.; Sweere, U.; Grefen, C.; Kunkel, T.; Fejes, E.; Nagy, F.; Schafer, E.; Harter, K. Functional cross-talk between two-component and phytochrome B signal transduction in Arabidopsis. J. Exp. Bot 2007, 58, 2595–2607. [Google Scholar]
- Brown, B.A.; Cloix, C.; Jiang, G.H.; Kaiserli, E.; Herzyk, P.; Kliebenstein, D.J.; Jenkins, G.I. A UV-B-specific signaling component orchestrates plant UV protection. Proc. Natl. Acad. Sci. USA 2005, 102, 18225–18230. [Google Scholar]
- Green, R.; Fluhr, R. UV-B-induced PR-1 accumulation is mediated by active oxygen species. Plant Cell 1995, 7, 203–212. [Google Scholar]
- Ulm, R.; Nagy, F. Signalling and gene regulation in response to ultraviolet light. Curr. Opin. Plant Biol 2005, 8, 477–482. [Google Scholar]
- Ng, W.O.; Grossman, A.R.; Bhaya, D. Multiple light inputs control phototaxis in Synechocystis sp. strain PCC6803. J. Bacteriol 2003, 185, 1599–1607. [Google Scholar]
- Masuda, S.; Ono, T.A. Biochemical characterization of the major adenylyl cyclase, Cya1, in the cyanobacterium Synechocystis sp. PCC 6803. FEBS Lett 2004, 577, 255–258. [Google Scholar]
- Okajima, K.; Yoshihara, S.; Fukushima, Y.; Geng, X.; Katayama, M.; Higashi, S.; Watanabe, M.; Sato, S.; Tabata, S.; Shibata, Y.; Itoh, S.; Ikeuchi, M. Biochemical and functional characterization of BLUF-type flavin-binding proteins of two species of cyanobacteria. J. Biochem 2005, 137, 741–750. [Google Scholar]
- Savakis, P.; de Causmaecker, S.; Angerer, V.; Ruppert, U.; Anders, K.; Essen, L.O.; Wilde, A. Light-induced alteration of c-di-GMP level controls motility of Synechocystis sp. PCC 6803. Mol. Microbiol 2012, 85, 239–251. [Google Scholar]
- Boehm, A.; Kaiser, M.; Li, H.; Spangler, C.; Kasper, C.A.; Ackermann, M.; Kaever, V.; Sourjik, V.; Roth, V.; Jenal, U. Second messenger-mediated adjustment of bacterial swimming velocity. Cell 2010, 141, 107–116. [Google Scholar]
- Jenal, U.; Malone, J. Mechanisms of cyclic-di-GMP signaling in bacteria. Annu. Rev. Genet 2006, 40, 385–407. [Google Scholar]
- Hengge, R. Principles of c-di-GMP signalling in bacteria. Nat. Rev. Microbiol 2009, 7, 263–273. [Google Scholar]
- Cadoret, J.C.; Rousseau, B.; Perewoska, I.; Sicora, C.; Cheregi, O.; Vass, I.; Houmard, J. Cyclic nucleotides, the photosynthetic apparatus and response to a UV-B stress in the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem 2005, 280, 33935–33944. [Google Scholar]
- Terauchi, K.; Ohmori, M. Blue light stimulates cyanobacterial motility via a cAMP signal transduction system. Mol. Microbiol 2004, 52, 303–309. [Google Scholar]
- Yoshimura, H.; Hisabori, T.; Yanagisawa, S.; Ohmori, M. Identification and characterization of a novel cAMP receptor protein in the cyanobacterium Synechocystis sp. PCC 6803. J. Biol. Chem 2000, 275, 6241–6245. [Google Scholar]
- Yoshimura, H.; Yoshihara, S.; Okamoto, S.; Ikeuchi, M.; Ohmori, M. A cAMP receptor protein, SYCRP1, is responsible for the cell motility of Synechocystis sp. PCC 6803. Plant Cell Physiol 2002, 43, 460–463. [Google Scholar]
- Terauchi, K.; Ohmori, M. An adenylate cyclase, Cya1, regulates cell motility in the cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol 1999, 40, 248–251. [Google Scholar]
- Bhaya, D.; Nakasugi, K.; Fazeli, F.; Burriesci, M.S. Phototaxis and impaired motility in adenylyl cyclase and cyclase receptor protein mutants of Synechocystis sp. strain PCC 6803. J. Bacteriol 2006, 188, 7306–7310. [Google Scholar]
- Iseki, M.; Matsunaga, S.; Murakami, A.; Ohno, K.; Shiga, K.; Yoshida, K.; Sugai, M.; Takahashi, T.; Hori, T.; Watanabe, M. A blue-light-activated adenylyl cyclase mediates photoavoidance in Euglena gracilis. Nature 2002, 415, 1047–1051. [Google Scholar]
- Ohmori, M.; Okamoto, S. Photoresponsive cAMP signal transduction in cyanobacteria. Photochem. Photobiol. Sci 2004, 3, 503–511. [Google Scholar]
- Arora, A.; Sairam, R.K.; Srivastava, G.C. Oxidative stress and antioxidative system in plants. Curr. Sci 2002, 82, 1227–1234. [Google Scholar]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen peroxide in plants: A versatile molecule of the reactive oxygen species network. J. Integr. Plant Biol 2008, 50, 2–18. [Google Scholar]
- Nishiyama, Y.; Yamamoto, H.; Allakhverdiev, S.I.; Inaba, M.; Yokota, A.; Murata, N. Oxidative stress inhibits the repair of photodamage to the photosynthetic machinery. EMBO J 2001, 20, 5587–5594. [Google Scholar]
- Blot, N.; Mella-Flores, D.; Six, C.; Le Corguille, G.; Boutte, C.; Peyrat, A.; Monnier, A.; Ratin, M.; Gourvil, P.; Campbell, D.A.; et al. Light history influences the response of the marine cyanobacterium Synechococcus sp. WH7803 to oxidative stress. Plant Physiol 2011, 156, 1934–1954. [Google Scholar]
- He, Y.Y.; Häder, D.-P. UV-B-induced formation of reactive oxygen species and oxidative damage of the cyanobacterium Anabaena sp.: Protective effects of ascorbic acid and N-acetyl-l-cysteine. J. Photochem. Photobiol. B 2002, 66, 115–124. [Google Scholar]
- Rastogi, R.P.; Singh, S.P.; Häder, D.-P.; Sinha, R.P. Detection of reactive oxygen species (ROS) by the oxidant-sensing probe 2′,7′-dichlorodihydrofluorescein diacetate in the cyanobacterium Anabaena variabilis PCC 7937. Biochem. Biophys. Res. Commun 2010, 397, 603–607. [Google Scholar]
- Huang, L.; McCluskey, M.P.; Ni, H.; LaRossa, R.A. Global gene expression profiles of the cyanobacterium Synechocystis sp. strain PCC 6803 in response to irradiation with UV-B and white light. J. Bacteriol 2002, 184, 6845–6858. [Google Scholar]
- Gao, Y.; Xiong, W.; Li, X.B.; Gao, C.F.; Zhang, Y.L.; Li, H.; Wu, Q.Y. Identification of the proteomic changes in Synechocystis sp. PCC 6803 following prolonged UV-B irradiation. J. Exp. Bot 2009, 60, 1141–1154. [Google Scholar]
- Nultsch, W.; Schuchart, H. A model of the phototactic reaction chain of the cyanobacterium Anabaena variabilis. Arch. Microbiol 1985, 142, 180–184. [Google Scholar]
- Shibata, H.; Baba, K.; Ochiai, H. Near-UV irradiation induces shock proteins in Anacystis nidulans R-2; Possible role of active oxygen. Plant Cell Physiol 1991, 32, 771–776. [Google Scholar]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schafer, E.; Nagy, F.; Jenkins, G.I.; Ulm, R. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol 2010, 61, 593–620. [Google Scholar]
- Christie, J.M.; Jenkins, G.I. Distinct UV-B and UV-A/blue light signal transduction pathways induce chalcone synthase gene expression in Arabidopsis cells. Plant Cell 1996, 8, 1555–1567. [Google Scholar]
- Smith, R.J.; Wilkins, A. A correlation between intracellular calcium and incident irradiance in Nostoc 6720. New Phytol 1988, 109, 157–161. [Google Scholar]
- Pitta, T.P.; Sherwood, E.E.; Kobel, A.M.; Berg, H.C. Calcium is required for swimming by the nonflagellated cyanobacterium Synechococcus strain WH8113. J. Bacteriol 1997, 179, 2524–2528. [Google Scholar]
- Torrecilla, I.; Leganes, F.; Bonilla, I.; Fernandez-Pinas, F. A calcium signal is involved in heterocyst differentiation in the cyanobacterium Anabaena sp. PCC7120. Microbiology 2004, 150, 3731–3739. [Google Scholar]
- Shi, Y.; Zhao, W.; Zhang, W.; Ye, Z.; Zhao, J. Regulation of intracellular free calcium concentration during heterocyst differentiation by HetR and NtcA in Anabaena sp. PCC 7120. Proc. Natl. Acad. Sci. USA 2006, 103, 11334–11339. [Google Scholar]
- Moon, Y.J.; Park, Y.M.; Chung, Y.H.; Choi, J.S. Calcium is involved in photomovement of cyanobacterium Synechocystis sp. PCC 6803. Photochem. Photobiol 2004, 79, 114–119. [Google Scholar]
- Onek, L.A.; Lea, P.J.; Smith, R.J. Isolation and characterization of a calmodulin-like protein from the cyanobacterium Nostoc sp. PCC 6720. Arch. Microbiol 1994, 161, 352–358. [Google Scholar]
- Fujisawa, T.; Narikawa, R.; Okamoto, S.; Ehira, S.; Yoshimura, H.; Suzuki, I.; Masuda, T.; Mochimaru, M.; Takaichi, S.; Awai, K.; et al. Genomic structure of an economically important cyanobacterium, Arthrospira (Spirulina) platensis NIES-39. DNA Res 2010, 17, 85–103. [Google Scholar]
- Richter, P.; Krywult, M.; Sinha, R.P.; Häder, D.-P. Calcium signals from heterocysts of Anabaena sp. after UV irradiation. J. Plant Physiol 1999, 154, 137–139. [Google Scholar]
- Sinha, R.P.; Richter, P.; Faddoul, J.; Braun, M.; Häder, D.-P. Effects of UV and visible light on cyanobacteria at the cellular level. Photochem. Photobiol. Sci 2002, 1, 553–559. [Google Scholar]
- Galland, P.; Senger, H. The role of pterins in the photoreception and metabolism of plants. Photochem. Photobiol 1988, 48, 811–820. [Google Scholar]
- Maier, J.; Ninnemann, H. Biosynthesis of pteridines in Neurospora crassa, Phycomyces blakesleeanus and Euglena gracilis: Detection and characterization of biosynthetic enzymes. Photochem. Photobiol 1995, 61, 43–53. [Google Scholar]
- Maier, J.; Ninnemann, H. Inhibition of light-dependent photomorphogenesis of sporangiophores from Phycomyces blakesleeanus by application of pteridine biosynthesis inhibitors. Photochem. Photobiol 1995, 61, 206–209. [Google Scholar]
- Lee, S.W.; Lee, H.W.; Chung, H.J.; Kim, Y.A.; Kim, Y.J.; Hahn, Y.; Chung, J.H.; Park, Y.S. Identification of the genes encoding enzymes involved in the early biosynthetic pathway of pteridines in Synechocystis sp. PCC 6803. FEMS Microbiol. Lett 1999, 176, 169–176. [Google Scholar]
- Choi, Y.K.; Hwang, Y.K.; Park, Y.S. Molecular cloning and disruption of a novel gene encoding UDP-glucose: Tetrahydrobiopterin alpha-glucosyltransferase in the cyanobacterium Synechococcus sp. PCC 7942. FEBS Lett 2001, 502, 73–78. [Google Scholar]
- Hwang, Y.K.; Kang, J.Y.; Woo, H.J.; Choi, Y.K.; Park, Y.S. Functional investigation of a gene encoding pteridine glycosyltransferase for cyanopterin synthesis in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 2002, 1570, 141–144. [Google Scholar]
- Nichol, C.A.; Smith, G.K.; Duch, D.S. Biosynthesis and metabolism of tetrahydrobiopterin and molybdopterin. Annu. Rev. Biochem 1985, 54, 729–764. [Google Scholar]
- Thöny, B.; Auerbach, G.; Blau, N. Tetrahydrobiopterin biosynthesis, regeneration and functions. Biochem. J. 2000, 347 Pt 1, 1–16. [Google Scholar]
- Kong, J.S.; Kang, J.Y.; Kim, H.L.; Kwon, O.S.; Lee, K.H.; Park, Y.S. 6-Pyruvoyltetrahydropterin synthase orthologs of either a single or dual domain structure are responsible for tetrahydrobiopterin synthesis in bacteria. FEBS Lett 2006, 580, 4900–4904. [Google Scholar]
- Hatfield, D.L.; van Baalen, C.; Forrest, H.S. Pteridines in blue green algae. Plant Physiol 1961, 36, 240–243. [Google Scholar]
- Forrest, H.S.; van Baalen, C. Microbiology of unconjugated pteridines. Annu. Rev. Microbiol 1970, 24, 91–108. [Google Scholar]
- Lee, H.W.; Oh, C.H.; Geyer, A.; Pfleiderer, W.; Park, Y.S. Characterization of a novel unconjugated pteridine glycoside, cyanopterin, in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 1999, 1410, 61–70. [Google Scholar]
- Chung, H.J.; Kim, Y.A.; Kim, Y.J.; Choi, Y.K.; Hwang, Y.K.; Park, Y.S. Purification and characterization of UDP-glucose: Tetrahydrobiopterin glucosyltransferase from Synechococcus sp. PCC 7942. Biochim. Biophys. Acta 2000, 1524, 183–188. [Google Scholar]
- Lee, Y.G.; Kim, A.H.; Park, M.B.; Kim, H.L.; Lee, K.H.; Park, Y.S. Molecular cloning of cyanobacterial pteridine glycosyltransferases that catalyze the transfer of either glucose or xylose to tetrahydrobiopterin. Appl. Environ. Microbiol 2010, 76, 7658–7661. [Google Scholar]
- Cho, S.H.; Na, J.U.; Youn, H.; Hwang, C.S.; Lee, C.H.; Kang, S.O. Tepidopterin, 1-O-(l-threo-biopterin-2′-yl)-β-N-acetylglucosamine from Chlorobium tepidum. Biochim. Biophys. Acta 1998, 1379, 53–60. [Google Scholar]
- Cha, K.W.; Pfleiderer, W.; Yim, J.J. Pteridines. Part CVI. Isolation and characterization of limipterin (1-O-(l-erythro-biopterin-2′-yl)-β-N-acetylglucosamine) and its 5,6,7,8-tetrahydro derivative from green sulfur bacterium Chlorobium limicola f. thiosulfatophilum NCIB 8327. Helv. Chim. Acta 1995, 78, 600–614. [Google Scholar]
- Lin, X.L.; White, R.H. Structure of solfapterin (erythro-neopterin-3′-d-2-deoxy-2- aminoglucopyranoside) isolated from the thermophilic archaebacterium Sulfolobus solfataricus. J. Bacteriol 1988, 170, 1396–1398. [Google Scholar]
- Maclean, F.I.; Fujita, Y.; Forrest, H.S.; Myers, J. Photosynthetic phosphorylation: Stimulation by pteridines and a comparison with phosphodoxin. Science 1965, 149, 636–638. [Google Scholar]
- Matsunaga, T.; Burgess, J.G.; Yamada, N.; Komatsu, K.; Yoshida, S.; Wachi, Y. An ultraviolet (UV-A) absorbing biopterin glucoside from the marine planktonic cyanobacterium Oscillatoria sp. Appl. Microbiol. Biotechnol 1993, 39, 250–253. [Google Scholar]
- Wachi, Y.; Burgess, J.G.; Iwamoto, K.; Yamada, N.; Nakamura, N.; Matsunaga, T. Effect of ultraviolet-A (UV-A) light on growth, photosynthetic activity and production of biopterin glucoside by the marine UV-A resistant cyanobacterium Oscillatoria sp. Biochim. Biophys. Acta 1995, 1244, 165–168. [Google Scholar]
- Noguchi, Y.; Ishii, A.; Matsushima, A.; Haishi, D.; Yasumuro, K.; Moriguchi, T.; Wada, T.; Kodera, Y.; Hiroto, M.; Nishimura, H.; Sekine, M.; Inada, Y. Isolation of biopterin-alpha-glucoside from Spirulina (Arthrospira) platensis and Its physiologic function. Mar. Biotechnol 1999, 1, 207–210. [Google Scholar]
- Saito, T.; Ishikura, H.; Hada, Y.; Fukui, K.; Kodera, Y.; Matsushim, A.; Inada, Y. Photostabilization of phycocyanin and anthocyanin in the presence of biopterin-α-glucoside from Spirulina platensis under ultraviolet ray. Dyes Pigments 2003, 56, 203–207. [Google Scholar]
- Ikawa, M.; Sasner, J.J.; Haney, J.F.; Foxall, T.L. Pterines of cyanobacterium Aphanizomenon flos-aquae. Phytochemistry 1995, 38, 1229–1232. [Google Scholar]
- Galland, P. Action spectra of photogeotropic equilibrium in Phycomyces wild-type and three behavioral mutants. Photochem. Photobiol 1983, 37, 221–228. [Google Scholar]
- Ninnemann, H. Participation of the molybdenum cofactor of nitrate reductase from Neurospora crassa in light-promoted conidation. J. Plant Physiol 1991, 137, 677–682. [Google Scholar]
- Johnson, J.L.; Hamm-Alvarez, S.; Payne, G.; Sancar, G.B.; Rajagopalan, K.V.; Sancar, A. Identification of the second chromophore of Escherichia coli and yeast DNA photolyases as 5,10-methenyltetrahydrofolate. Proc. Natl. Acad. Sci. USA 1988, 85, 2046–2050. [Google Scholar]
- Galland, P.; Lipson, E.D. Modified action spectra of photogeotropic equilibrium in Phycomyces blakesleeanus mutants with defects in genes madA, madB, madC, and madH. Photochem. Photobiol 1985, 41, 331–335. [Google Scholar]
- Galland, P.; Lipson, E.D. Action spectra for phototropic balance in Phycomyces blakesleeanus: Dependence on reference wavelength and intensity range. Photochem. Photobiol 1985, 41, 323–329. [Google Scholar]
- Galland, P.; Lipson, E.D. Blue-light reception in Phycomyces phototropism: Evidence for two photosystems operating in low- and high-intensity ranges. Proc. Natl. Acad. Sci. USA 1987, 84, 104–108. [Google Scholar]
- Hohl, N.; Galland, P.; Senger, H. Altered pterin patterns in photobehavioral mutants of Phycomyces blakesleeanus. Photochem. Photobiol 1992, 55, 239–245. [Google Scholar]
- Hohl, N.; Galland, P.; Senger, H. Altered flavin patterns in photobehavioral mutants of Phycomyces blakesleeanus. Photochem. Photobiol 1992, 55, 247–255. [Google Scholar]
- Sancar, A. Structure and function of DNA photolyase. Biochemistry 1994, 33, 2–9. [Google Scholar]
- Diehn, B. Action spectra of the phototactic responses in Euglena. Biochim. Biophys. Acta 1969, 177, 136–143. [Google Scholar]
- Matsunaga, S.; Hori, T.; Takahashi, T.; Kubota, M.; Watanabe, M.; Okamoto, K.; Masuda, K.; Sugai, M. Discovery of signaling effect of UV-B/C light in the extended UV-A/blue-type action spectra for step-down and step-up photophobic responses in the unicellular flagellate alga Euglena gracilis. Protoplasma 1998, 201, 45–52. [Google Scholar]
- Häder, D.-P.; Lebert, M. The photoreceptor for phototaxis in the photosynthetic flagellate Euglena gracilis. Photochem. Photobiol 1998, 68, 260–265. [Google Scholar]
- Brodhun, B.; Häder, D.-P. Photoreceptor proteins and pigments in the paraflagellar body of the flaellate Euglena gracilis. Photochem. Photobiol 1990, 52, 865–871. [Google Scholar]
- Schmidt, W.; Galland, P.; Senger, H.; Furuya, M. Microspectrophotometry of Euglena gracilis. Planta 1990, 182, 375–381. [Google Scholar]
- Galland, P.; Keiner, P.; Dörmemann, D.; Senger, H.; Brodhun, B.; Häder, D.-P. Pterin- and flavin-like fluorescence associated with isolated flagella of Euglena gracilis. Photochem. Photobiol 1990, 51, 675–680. [Google Scholar]
- Selbach, M.; Häder, D.-P.; Kuhlmann, H.-W. Phototaxis in Chlamydodon mnemosyne: Determination of the illuminance-response curve and the action spectrum. J. Photochem. Photobiol. B 1999, 49, 35–40. [Google Scholar]
- Selbach, M.; Kuhlmann, H.W. Structure, fluorescent properties and proposed function in phototaxis of the stigma apparatus in the ciliate Chlamydodon mnemosyne. J. Exp. Biol 1999, 202, 919–927. [Google Scholar]
- Portwich, A.; Garcia-Pichel, F. A novel prokaryotic UVB photoreceptor in the cyanobacterium Chlorogloeopsis PCC 6912. Photochem. Photobiol 2000, 71, 493–498. [Google Scholar]
- Maier, J.; Schmid, R. Inhibition of pteridine biosynthesis eliminates blue-light dependent stimulation of red-light saturated photosynthesis in Laminaria saccharina (L.) Lamouroux. J. Photochem. Photobiol. B 1997, 38, 274–278. [Google Scholar]
- Huang, Y.; Baxter, R.; Smith, B.S.; Partch, C.L.; Colbert, C.L.; Deisenhofer, J. Crystal structure of cryptochrome 3 from Arabidopsis thaliana and its implications for photolyase activity. Proc. Natl. Acad. Sci. USA 2006, 103, 17701–17706. [Google Scholar]
- Klar, T.; Pokorny, R.; Moldt, J.; Batschauer, A.; Essen, L.O. Cryptochrome 3 from Arabidopsis thaliana: Structural and functional analysis of its complex with a folate light antenna. J. Mol. Biol 2007, 366, 954–964. [Google Scholar]
- Ahmad, M.; Cashmore, A.R. Seeing blue: The discovery of cryptochrome. Plant Mol. Biol 1996, 30, 851–861. [Google Scholar]
- Partch, C.L.; Sancar, A. Photochemistry and photobiology of cryptochrome blue-light photopigments: The search for a photocycle. Photochem. Photobiol 2005, 81, 1291–1304. [Google Scholar]
- Lin, C.; Shalitin, D. Cryptochrome structure and signal transduction. Annu. Rev. Plant Biol 2003, 54, 469–496. [Google Scholar]
- Yu, X.; Liu, H.; Klejnot, J.; Lin, C. The cryptochrome blue light receptors. Arabidopsis Book 2010, 8, e0135. [Google Scholar]
- Hitomi, K.; Okamoto, K.; Daiyasu, H.; Miyashita, H.; Iwai, S.; Toh, H.; Ishiura, M.; Todo, T. Bacterial cryptochrome and photolyase: Characterization of two photolyase-like genes of Synechocystis sp. PCC6803. Nucleic Acids Res 2000, 28, 2353–2362. [Google Scholar]
- Ng, W.O.; Pakrasi, H.B. DNA photolyase homologs are the major UV resistance factors in the cyanobacterium Synechocystis sp. PCC 6803. Mol. Gen. Genet 2001, 264, 924–930. [Google Scholar]
- Brudler, R.; Hitomi, K.; Daiyasu, H.; Toh, H.; Kucho, K.; Ishiura, M.; Kanehisa, M.; Roberts, V.A.; Todo, T.; Tainer, J.A.; Getzoff, E.D. Identification of a new cryptochrome class. Structure, function, and evolution. Mol. Cell 2003, 11, 59–67. [Google Scholar]
- Brautigam, C.A.; Smith, B.S.; Ma, Z.; Palnitkar, M.; Tomchick, D.R.; Machius, M.; Deisenhofer, J. Structure of the photolyase-like domain of cryptochrome 1 from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 12142–12147. [Google Scholar]
- Kleine, T.; Lockhart, P.; Batschauer, A. An Arabidopsis protein closely related to Synechocystis cryptochrome is targeted to organelles. Plant J 2003, 35, 93–103. [Google Scholar]
- Saxena, C.; Wang, H.; Kavakli, I.H.; Sancar, A.; Zhong, D. Ultrafast dynamics of resonance energy transfer in cryptochrome. J. Am. Chem. Soc 2005, 127, 7984–7985. [Google Scholar]
- Moharikar, S.; D’Souza, J.S.; Kulkarni, A.B.; Rao, B.J. Apoptotic-like cell death pathway is induced in unicellular chlorophyte Chlamydomonas reinhardtii (Chlorophyceae) cells following UV irradiation: Detection and functional analyses. J. Phycol 2006, 42, 423–433. [Google Scholar]
- Jiménez, C.; Capasso, J.M.; Edelstein, C.L.; Rivard, C.J.; Lucia, S.; Breusegem, S.; Berl, T.; Segovia, M. Different ways to die: Cell death modes of the unicellular chlorophyte Dunaliella viridis exposed to various environmental stresses are mediated by the caspase-like activity DEVDase. J. Exp. Bot 2009, 60, 815–828. [Google Scholar]
- Ning, S.B.; Guo, H.L.; Wang, L.; Song, Y.C. Salt stress induces programmed cell death in prokaryotic organism Anabaena. J. Appl. Microbiol 2002, 93, 15–28. [Google Scholar]
- Ross, C.; Santiago-Vazquez, L.; Paul, V. Toxin release in response to oxidative stress and programmed cell death in the cyanobacterium Microcystis aeruginosa. Aquat. Toxicol 2006, 78, 66–73. [Google Scholar]
- Ding, Y.; Gan, N.; Li, J.; Sedmak, B.; Song, L. Hydrogen peroxide induces apoptotic-like cell death in Microcystis aeruginosa (Chroococcales, Cyanobacteria) in a dose-dependent manner. Phycologia 2012, 51, 567–575. [Google Scholar]
- Berman-Frank, I.; Bidle, K.D.; Haramaty, L.; Falkowski, P.G. The demise of the marine cyanobacterium, Trichodesmium spp. via an autocatalyzed cell death pathway. Limnol. Oceanogr 2004, 49, 997–1005. [Google Scholar]
- Bidle, K.D.; Haramaty, L.; Barcelos, E.R.J.; Falkowski, P. Viral activation and recruitment of metacaspases in the unicellular coccolithophore, Emiliania huxleyi. Proc. Natl. Acad. Sci. USA 2007, 104, 6049–6054. [Google Scholar]
- Le Chat, L.; Sinden, R.E.; Dessens, J.T. The role of metacaspase 1 in Plasmodium berghei development and apoptosis. Mol. Biochem. Parasitol 2007, 153, 41–47. [Google Scholar]
- Robson, G.D. Programmed cell death in the aspergilli and other filamentous fungi. Med. Mycol 2006, 44, S109–S114. [Google Scholar]
- Trzyna, W.C.; Legras, X.D.; Cordingley, J.S. A type-1 metacaspase from Acanthamoeba castellanii. Microbiol. Res 2008, 163, 414–423. [Google Scholar]
- He, R.; Drury, G.E.; Rotari, V.I.; Gordon, A.; Willer, M.; Farzaneh, T.; Woltering, E.J.; Gallois, P. Metacaspase-8 modulates programmed cell death induced by ultraviolet light and H2O2 in Arabidopsis. J. Biol. Chem 2008, 283, 774–783. [Google Scholar]
- Berman-Frank, I.; Rosenberg, G.; Levitan, O.; Haramaty, L.; Mari, X. Coupling between autocatalytic cell death and transparent exopolymeric particle production in the marine cyanobacterium Trichodesmium. Environ. Microbiol 2007, 9, 1415–1422. [Google Scholar]
- Bidle, K.D.; Falkowski, P.G. Cell death in planktonic, photosynthetic microorganisms. Nat. Rev. Microbiol 2004, 2, 643–655. [Google Scholar]
- Jiang, Q.; Qin, S.; Wu, Q.Y. Genome-wide comparative analysis of metacaspases in unicellular and filamentous cyanobacteria. BMC Genomics 2010, 11, 198. [Google Scholar]
- Lane, N. Marine microbiology: Origins of death. Nature 2008, 453, 583–585. [Google Scholar]



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moon, Y.-J.; Kim, S.I.; Chung, Y.-H. Sensing and Responding to UV-A in Cyanobacteria. Int. J. Mol. Sci. 2012, 13, 16303-16332. https://doi.org/10.3390/ijms131216303
Moon Y-J, Kim SI, Chung Y-H. Sensing and Responding to UV-A in Cyanobacteria. International Journal of Molecular Sciences. 2012; 13(12):16303-16332. https://doi.org/10.3390/ijms131216303
Chicago/Turabian StyleMoon, Yoon-Jung, Seung Il Kim, and Young-Ho Chung. 2012. "Sensing and Responding to UV-A in Cyanobacteria" International Journal of Molecular Sciences 13, no. 12: 16303-16332. https://doi.org/10.3390/ijms131216303
APA StyleMoon, Y.-J., Kim, S. I., & Chung, Y.-H. (2012). Sensing and Responding to UV-A in Cyanobacteria. International Journal of Molecular Sciences, 13(12), 16303-16332. https://doi.org/10.3390/ijms131216303