The Role of Forkhead Box Q1 Transcription Factor in Ovarian Epithelial Carcinomas

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
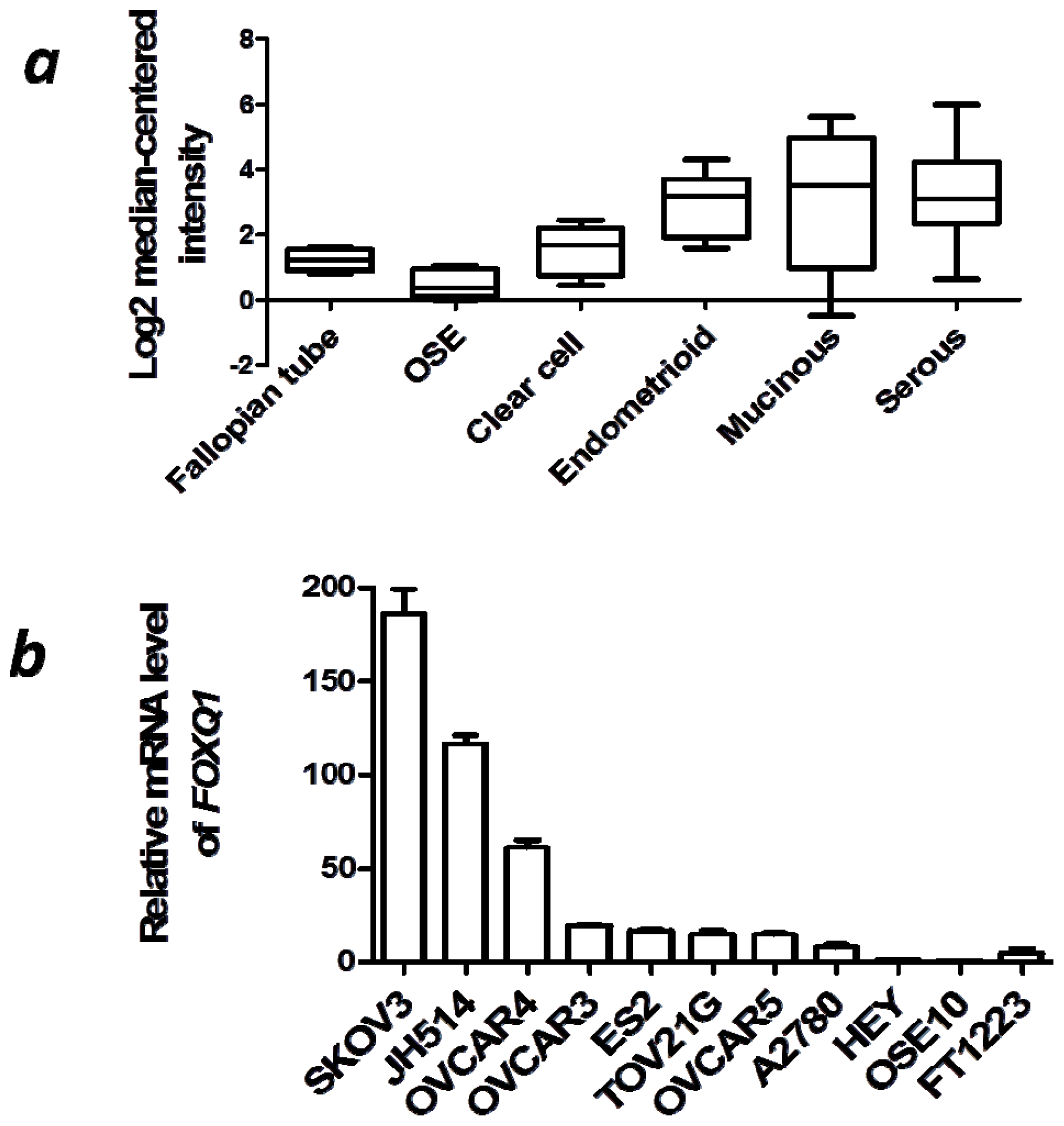
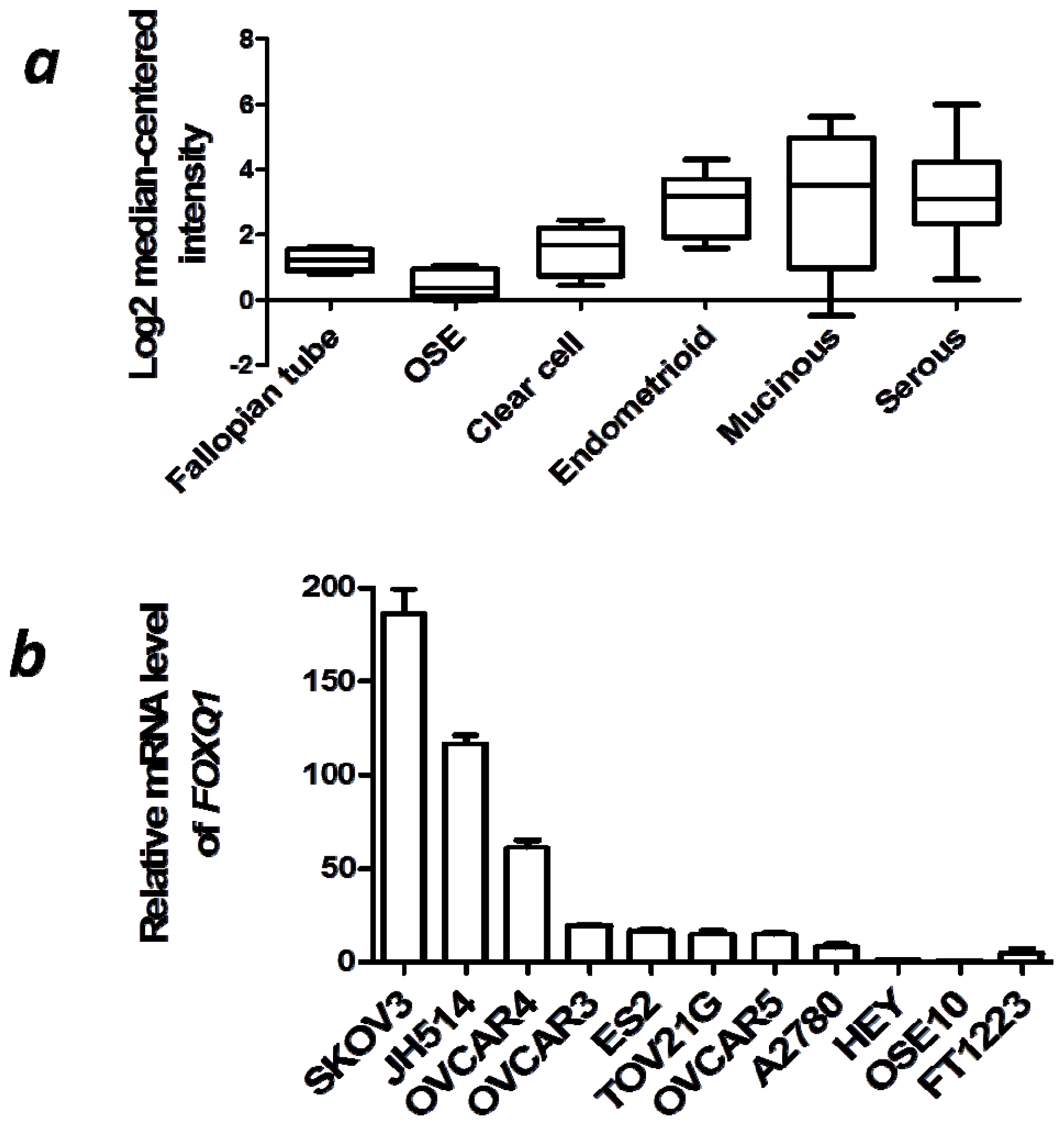
2.1. FOXQ1 is Upregulated in Ovarian Cancer Tissues and Cell Lines
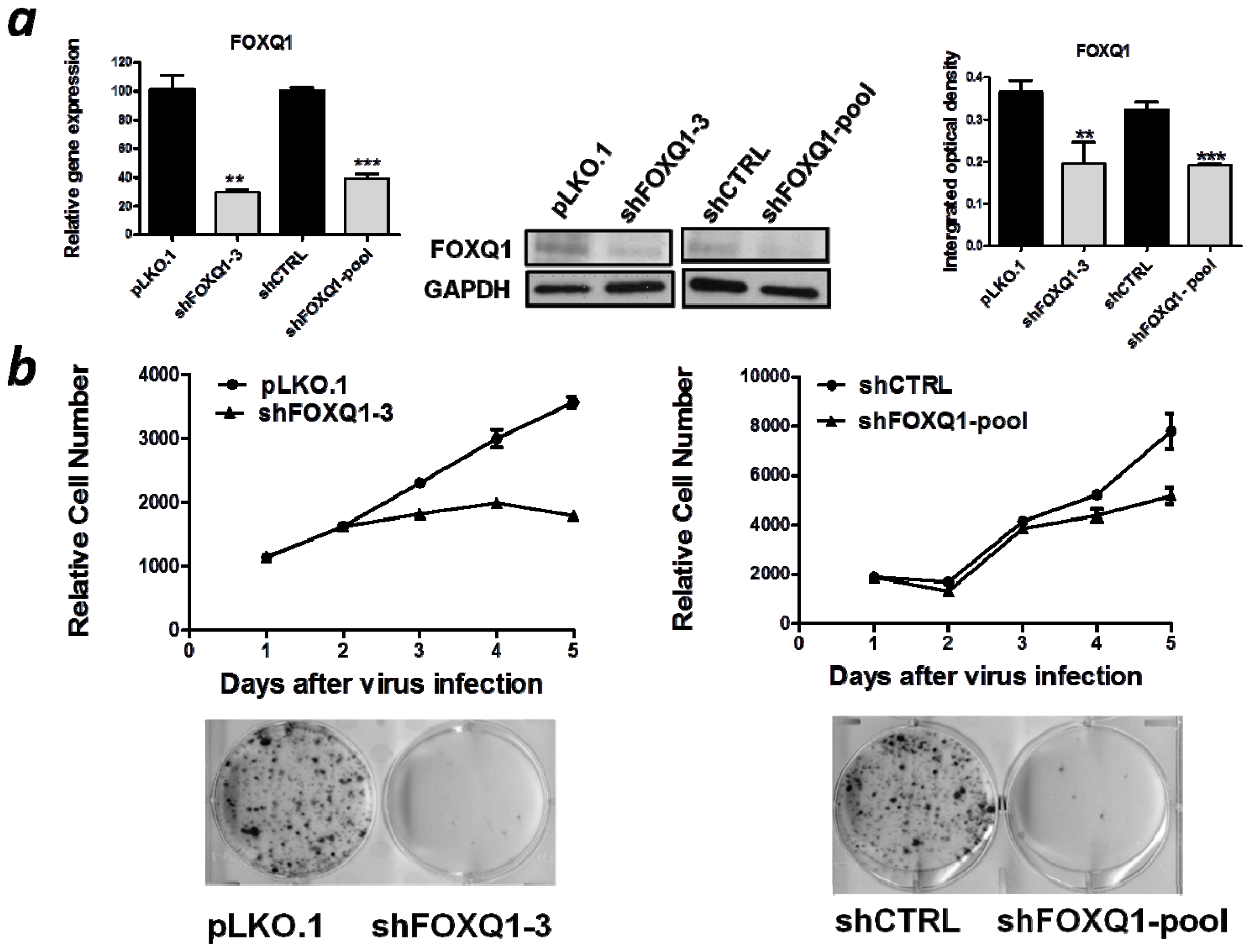
2.2. FOXQ1 Knockdown Reduces Cell Growth and Clonogenicity
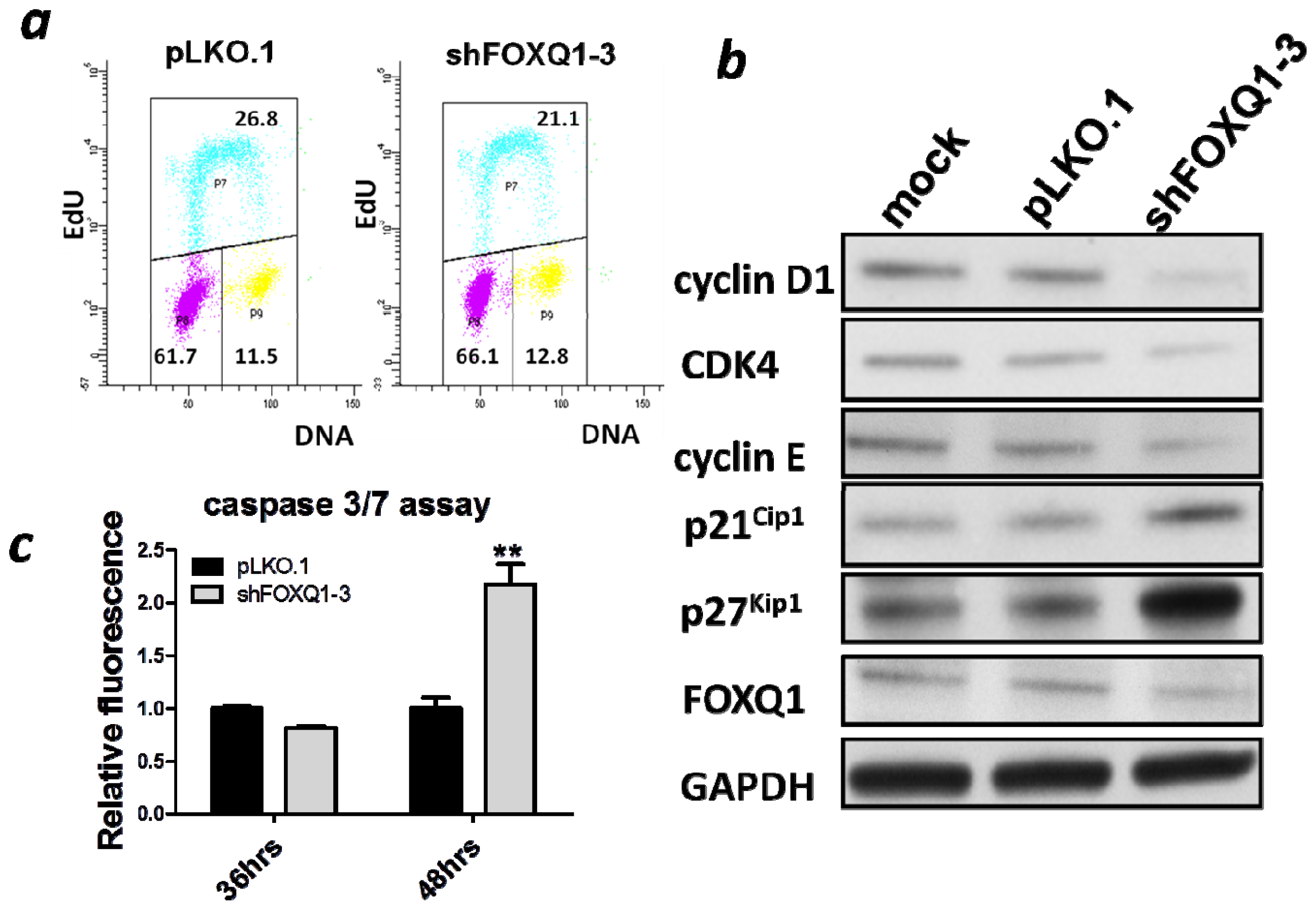
2.3. FOXQ1 Depletion Leads to G1/S Cell Cycle Arrest and Alters Cell Cycle Regulators
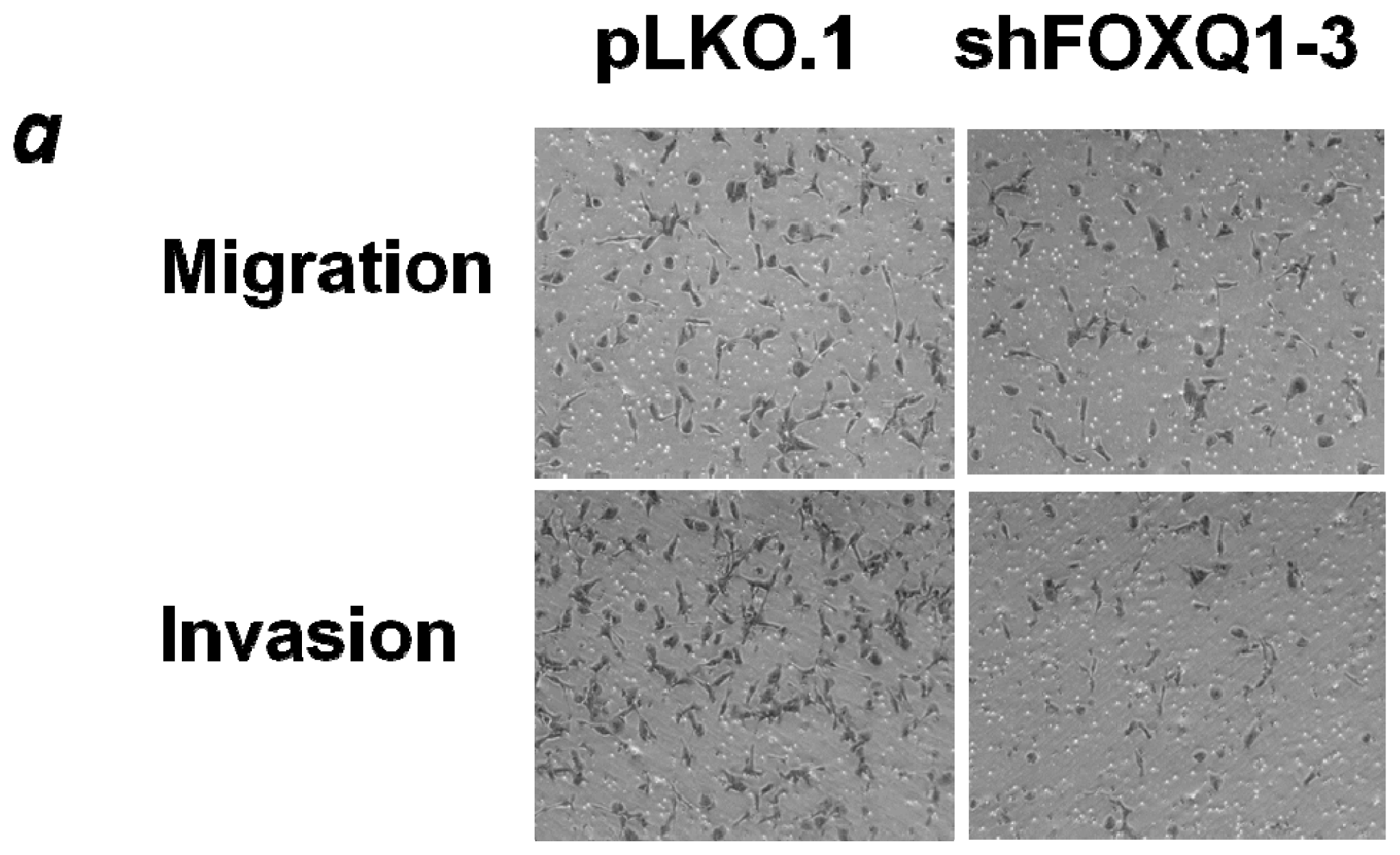
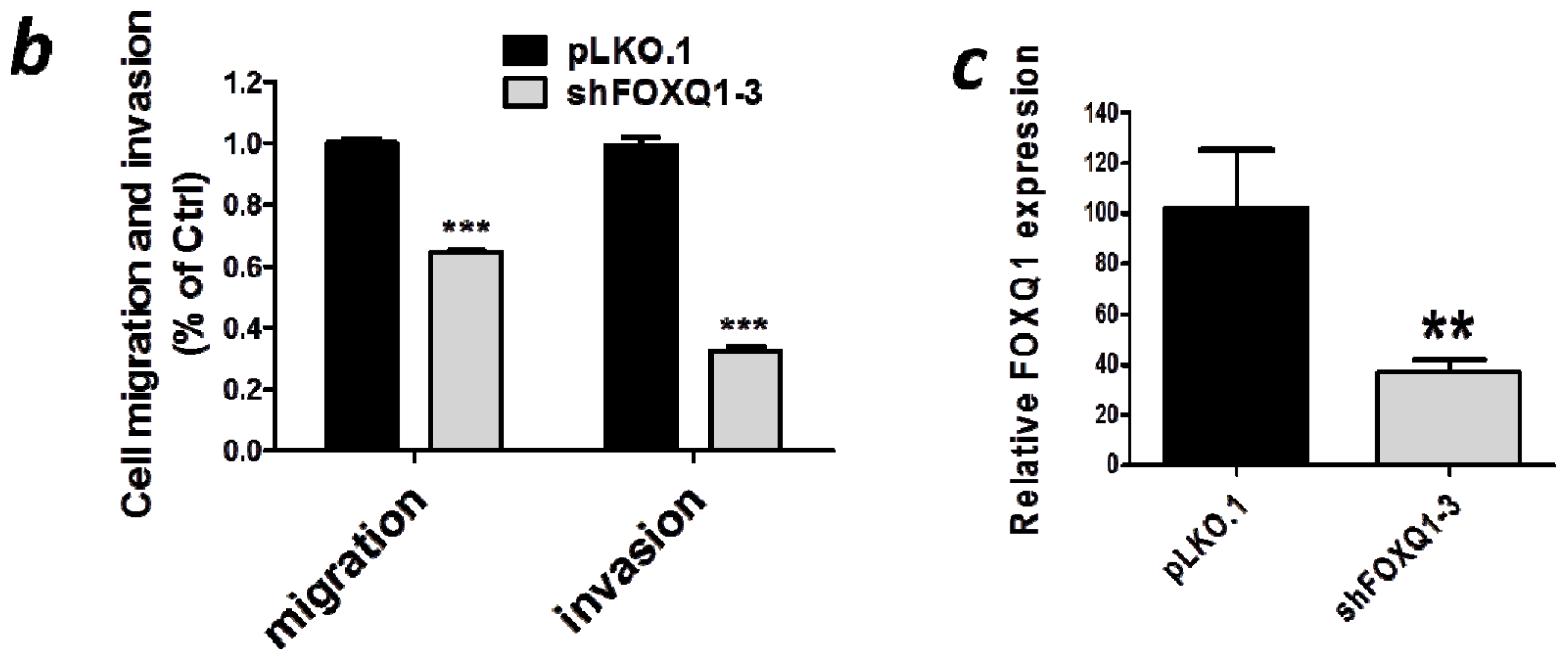
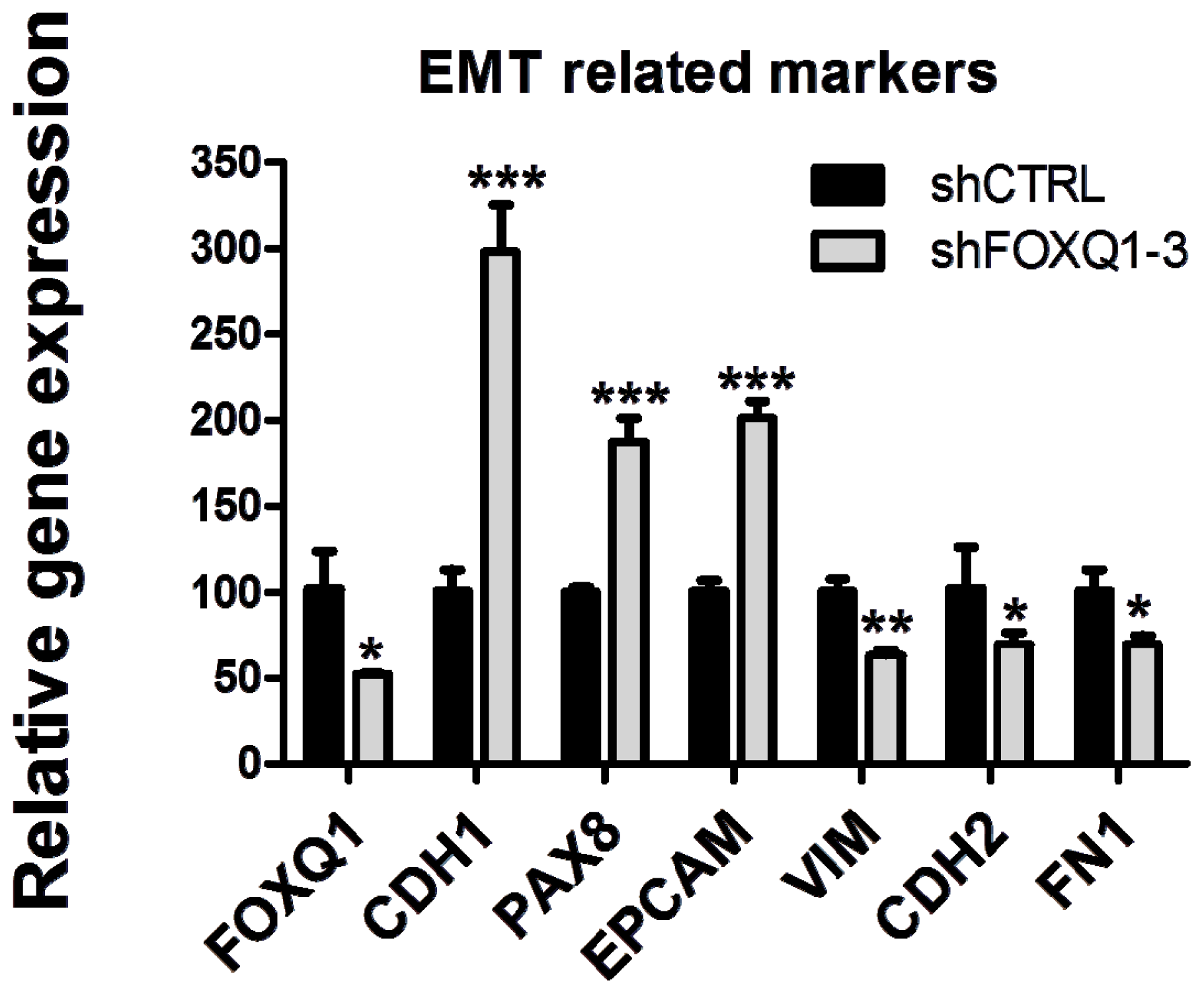
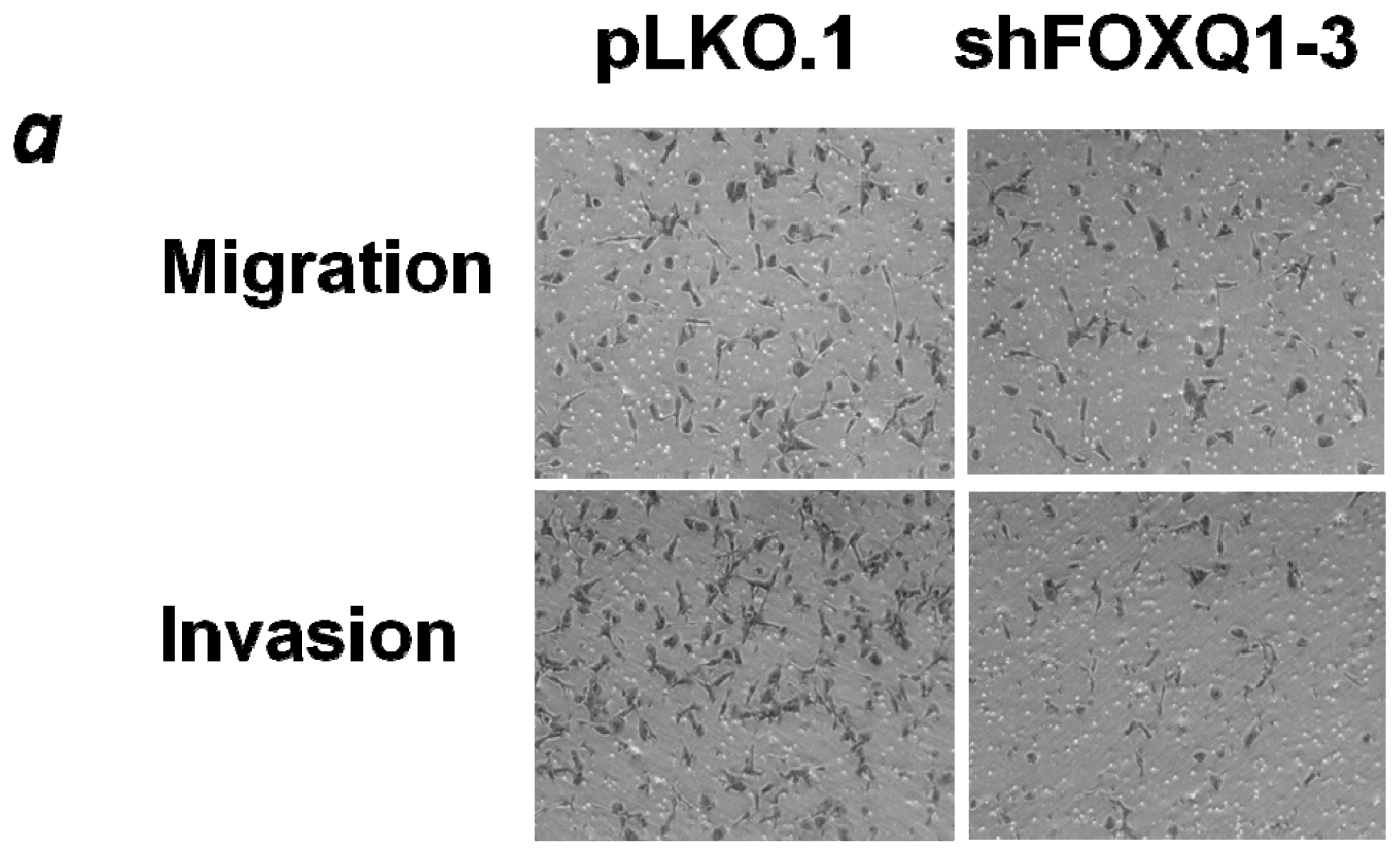
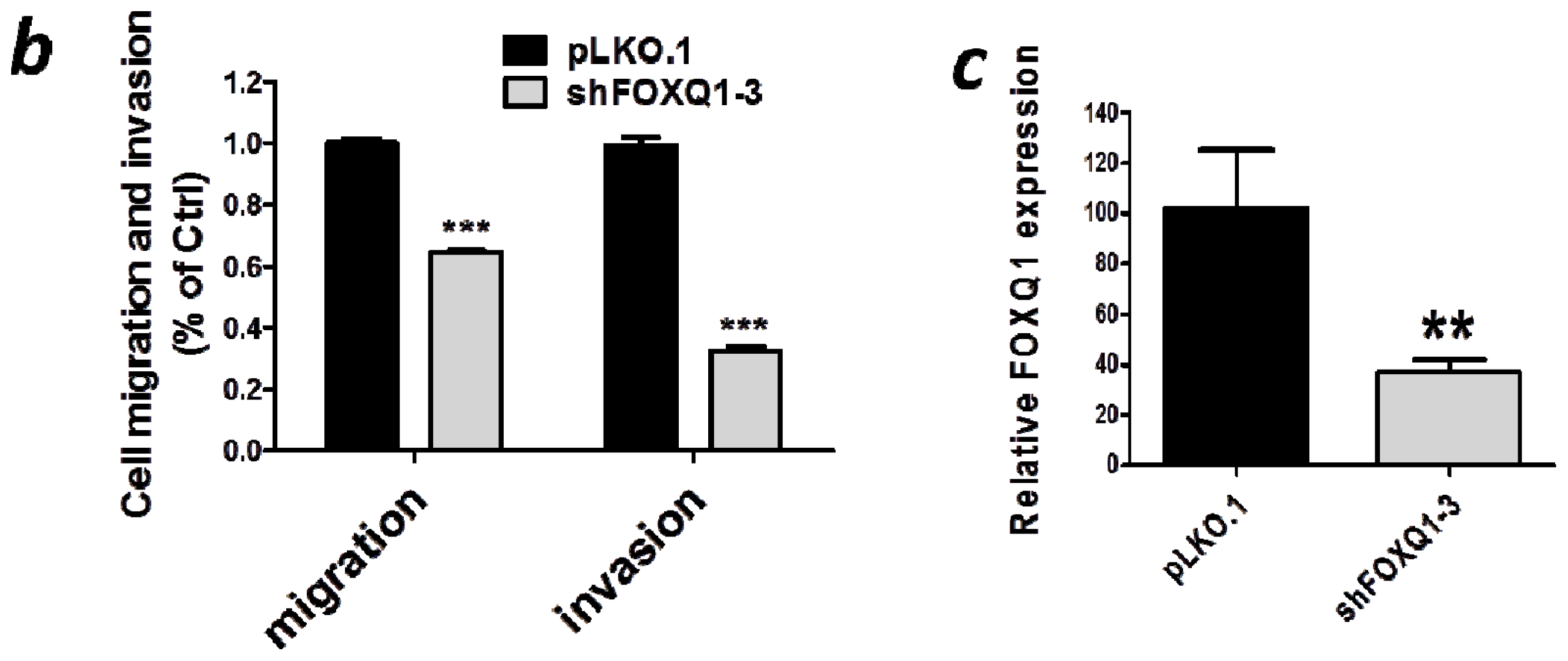
2.4. FOXQ1 Knockdown Decreases Ovarian Cancer Cell Movement and Affects the Expression of Epithelial-Mesenchymal Transition (EMT) Markers
3. Experimental Section
3.1. Cell Line and Culture Conditions
3.2. Quantitative Real-Time PCR
3.3. Western Blots
3.4. FOXQ1 Gene Knockdown
3.5. Cellular Proliferation Assay
3.6. Colony Formation Assay
3.7. Flow Cytometry Assay
3.8. Caspase Assay
3.9. Migration and Invasion Assay
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Kurman, R.J.; Shih Ie, M. The origin and pathogenesis of epithelial ovarian cancer: A proposed unifying theory. Am. J. Surg. Pathol 2010, 34, 433–443. [Google Scholar]
- Kurman, R.J.; Shih Ie, M. Molecular pathogenesis and extraovarian origin of epithelial ovarian cancer--shifting the paradigm. Hum. Pathol 2011, 42, 918–931. [Google Scholar]
- Nakayama, K.; Nakayama, N.; Davidson, B.; Sheu, J.J.; Jinawath, N.; Santillan, A.; Salani, R.; Bristow, R.E.; Morin, P.J.; Kurman, R.J.; et al. A BTB/POZ protein, NAC-1, is related to tumor recurrence and is essential for tumor growth and survival. Proc. Natl. Acad. Sci. USA 2006, 103, 18739–18744. [Google Scholar]
- Myatt, S.S.; Lam, E.W. The emerging roles of forkhead box (Fox) proteins in cancer. Nat. Rev. Cancer 2007, 7, 847–859. [Google Scholar]
- Bieller, A.; Pasche, B.; Frank, S.; Glaser, B.; Kunz, J.; Witt, K.; Zoll, B. Isolation and characterization of the human forkhead gene FOXQ1. DNA Cell Biol 2001, 20, 555–561. [Google Scholar]
- Martinez-Ceballos, E.; Chambon, P.; Gudas, L.J. Differences in gene expression between wild type and Hoxa1 knockout embryonic stem cells after retinoic acid treatment or leukemia inhibitory factor (LIF) removal. J. Biol. Chem 2005, 280, 16484–16498. [Google Scholar]
- Potter, C.S.; Peterson, R.L.; Barth, J.L.; Pruett, N.D.; Jacobs, D.F.; Kern, M.J.; Argraves, W.S.; Sundberg, J.P.; Awgulewitsch, A. Evidence that the satin hair mutant gene FOXQ1 is among multiple and functionally diverse regulatory targets for Hoxc13 during hair follicle differentiation. J. Biol. Chem 2006, 281, 29245–29255. [Google Scholar]
- Qiao, Y.Y.; Jiang, X.; Lee, S.T.; Karuturi, R.K.M.; Hooi, S.C.; Yu, Q.A. FOXQ1 regulates epithelial-mesenchymal transition in human cancers. Cancer Res 2011, 71, 3076–3086. [Google Scholar]
- Zhang, H.J.; Meng, F.Y.; Liu, G.; Zhang, B.; Zhu, J.; Wu, F.; Ethier, S.P.; Miller, F.; Wu, G.J. Forkhead transcription factor FOXQ1 promotes epithelial-mesenchymal transition and breast cancer metastasis. Cancer Res 2011, 71, 1292–1301. [Google Scholar]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar]
- Marquez, R.T.; Baggerly, K.A.; Patterson, A.P.; Liu, J.; Broaddus, R.; Frumovitz, M.; Atkinson, E.N.; Smith, D.I.; Hartmann, L.; Fishman, D.; et al. Patterns of gene expression in different histotypes of epithelial ovarian cancer correlate with those in normal fallopian tube, endometrium, and colon. Clin. Cancer Res 2005, 11, 6116–6126. [Google Scholar]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar]
- Feuerborn, A.; Srivastava, P.K.; Kuffer, S.; Grandy, W.A.; Sijmonsma, T.P.; Gretz, N.; Brors, B.; Grone, H.J. The forkhead factor FOXQ1 influences epithelial differentiation. J. Cell. Physiol 2011, 226, 710–719. [Google Scholar]
- Kaneda, H.; Arao, T.; Tanaka, K.; Tamura, D.; Aomatsu, K.; Kudo, K.; Sakai, K.; De Velasco, M.A.; Matsumoto, K.; Fujita, Y.; et al. FOXQ1 is overexpressed in colorectal cancer and enhances tumorigenicity and tumor growth. Cancer Res 2010, 70, 2053–2063. [Google Scholar]
- Feng, J.; Zhang, X.; Zhu, H.; Wang, X.; Ni, S.; Huang, J. FOXQ1 overexpression influences poor prognosis in non-small cell lung cancer, associates with the phenomenon of EMT. PLoS One 2012, 7, e39937. [Google Scholar]



© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gao, M.; Shih, I.-M.; Wang, T.-L. The Role of Forkhead Box Q1 Transcription Factor in Ovarian Epithelial Carcinomas. Int. J. Mol. Sci. 2012, 13, 13881-13893. https://doi.org/10.3390/ijms131113881
Gao M, Shih I-M, Wang T-L. The Role of Forkhead Box Q1 Transcription Factor in Ovarian Epithelial Carcinomas. International Journal of Molecular Sciences. 2012; 13(11):13881-13893. https://doi.org/10.3390/ijms131113881
Chicago/Turabian StyleGao, Min, Ie-Ming Shih, and Tian-Li Wang. 2012. "The Role of Forkhead Box Q1 Transcription Factor in Ovarian Epithelial Carcinomas" International Journal of Molecular Sciences 13, no. 11: 13881-13893. https://doi.org/10.3390/ijms131113881
APA StyleGao, M., Shih, I.-M., & Wang, T.-L. (2012). The Role of Forkhead Box Q1 Transcription Factor in Ovarian Epithelial Carcinomas. International Journal of Molecular Sciences, 13(11), 13881-13893. https://doi.org/10.3390/ijms131113881