Polyphenol Content and Modulatory Activities of Some Tropical Dietary Plant Extracts on the Oxidant Activities of Neutrophils and Myeloperoxidase


Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. Biochemical Investigations
2.1.1.1. Cell viability in Presence of the Plant Extracts Tested
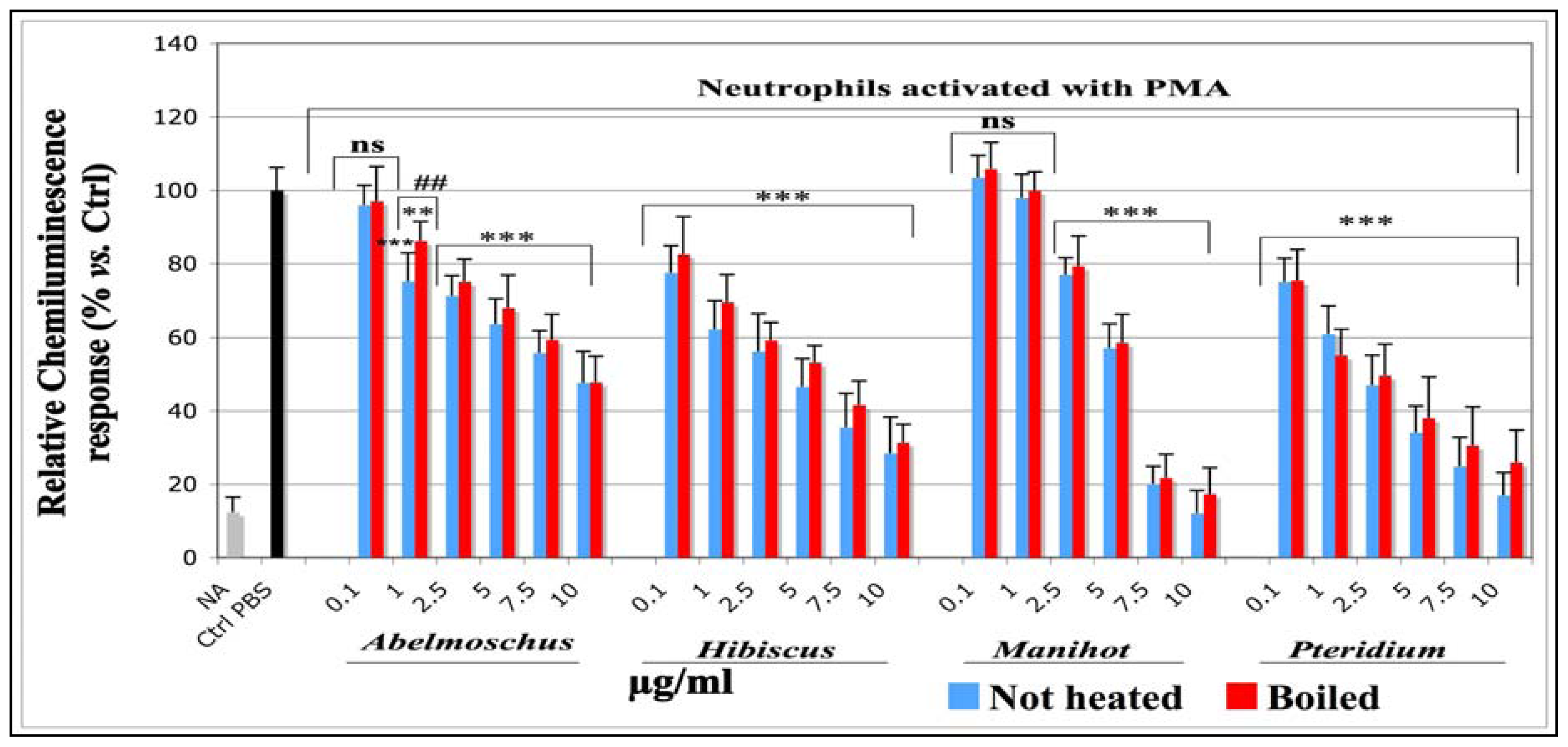
2.1.1.2. Effects on the Total ROS Amount Produced by Neutrophils Activated with PMA (CL assay)
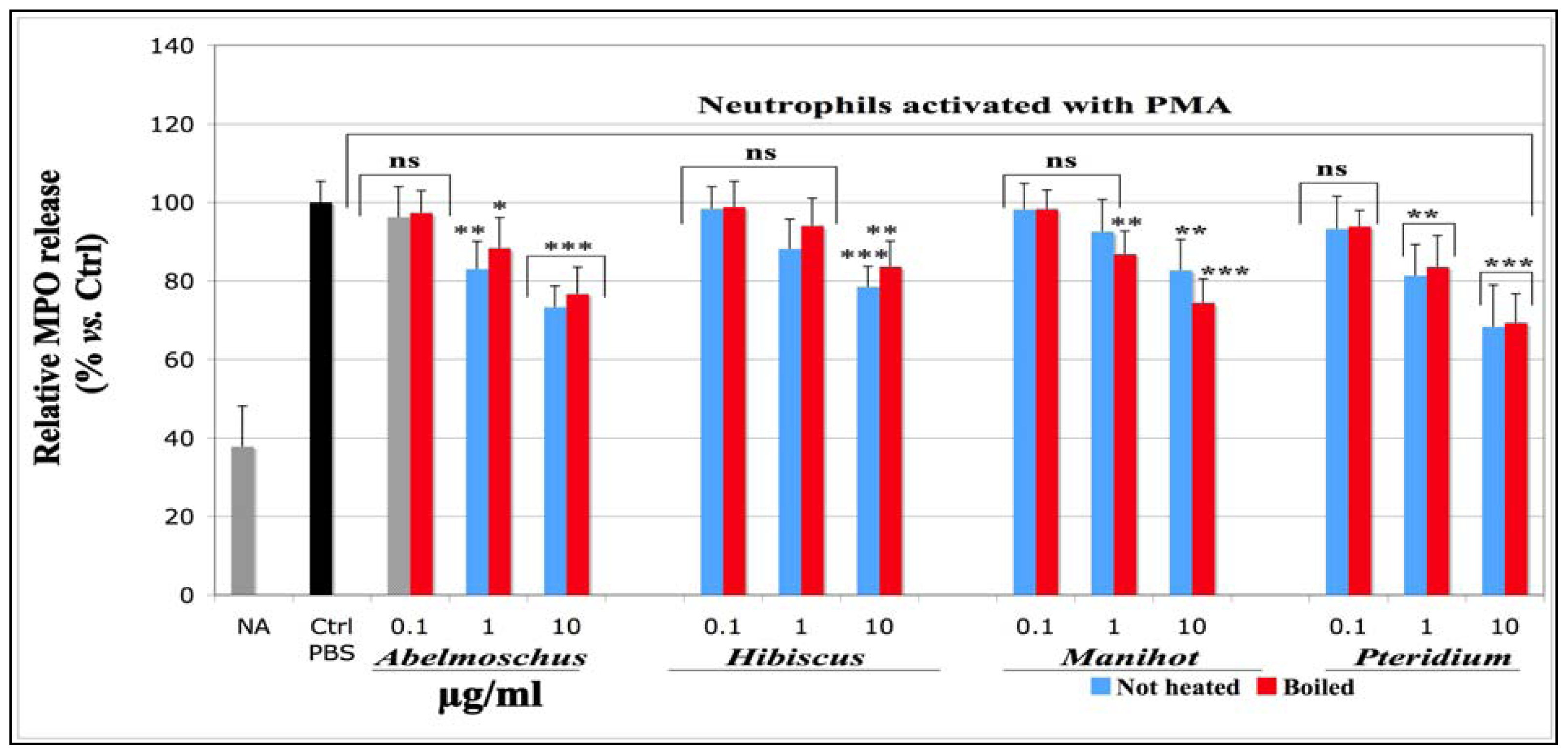
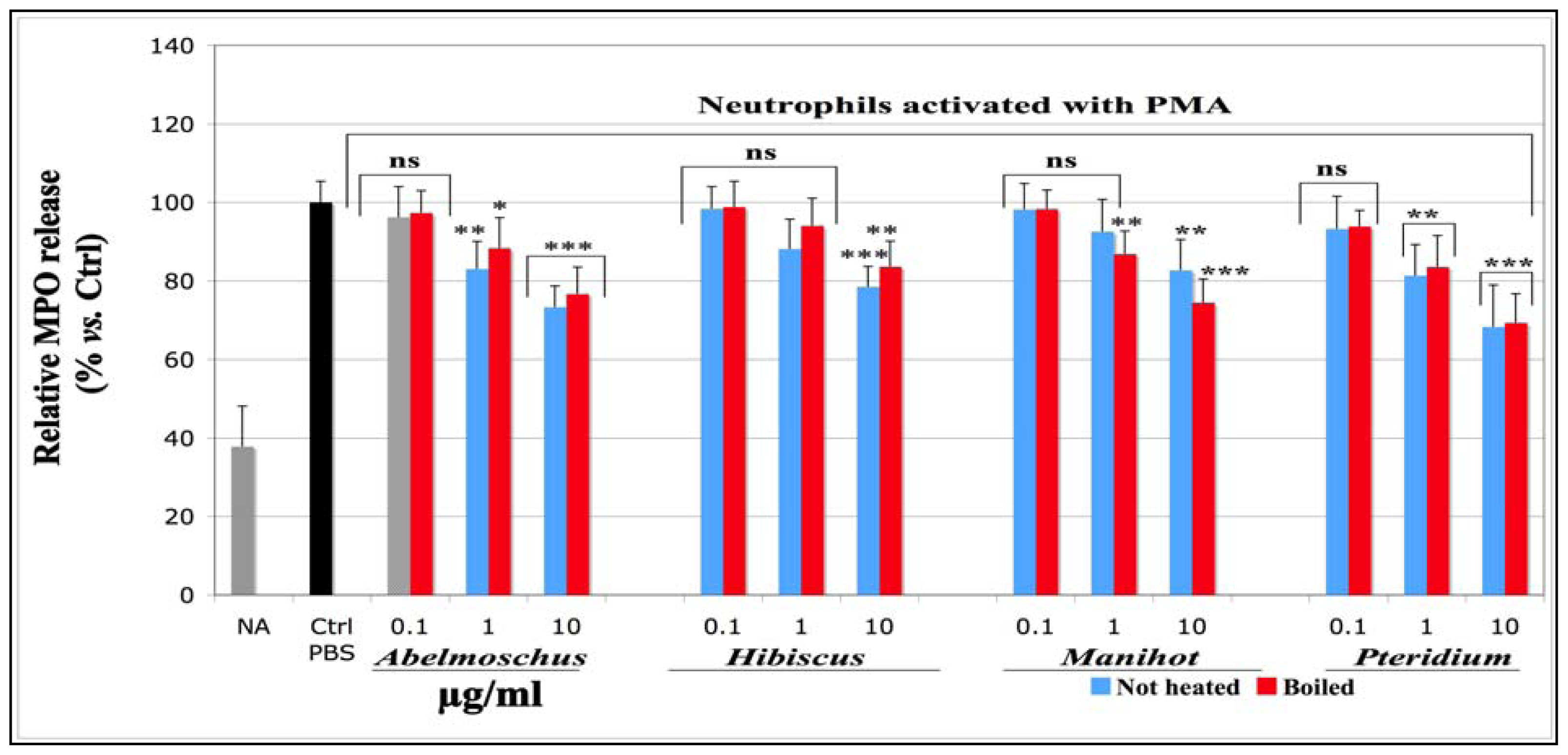
2.1.1.3. Effects on Total MPO Amount Released by Neutrophils Stimulated with PMA (MPO-ELISA)
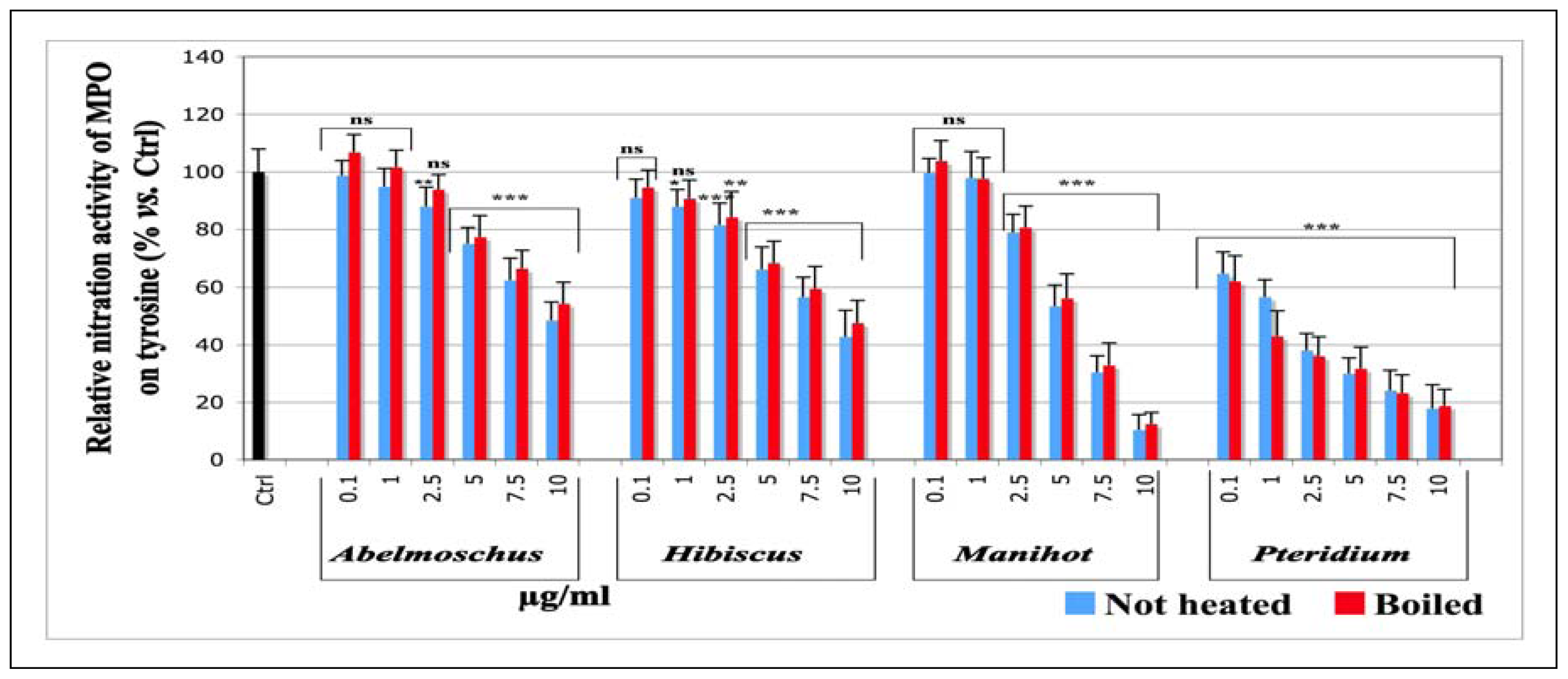
2.1.1.4. Effects on the Nitration Activity of MPO Measured by Tyrosine nitration
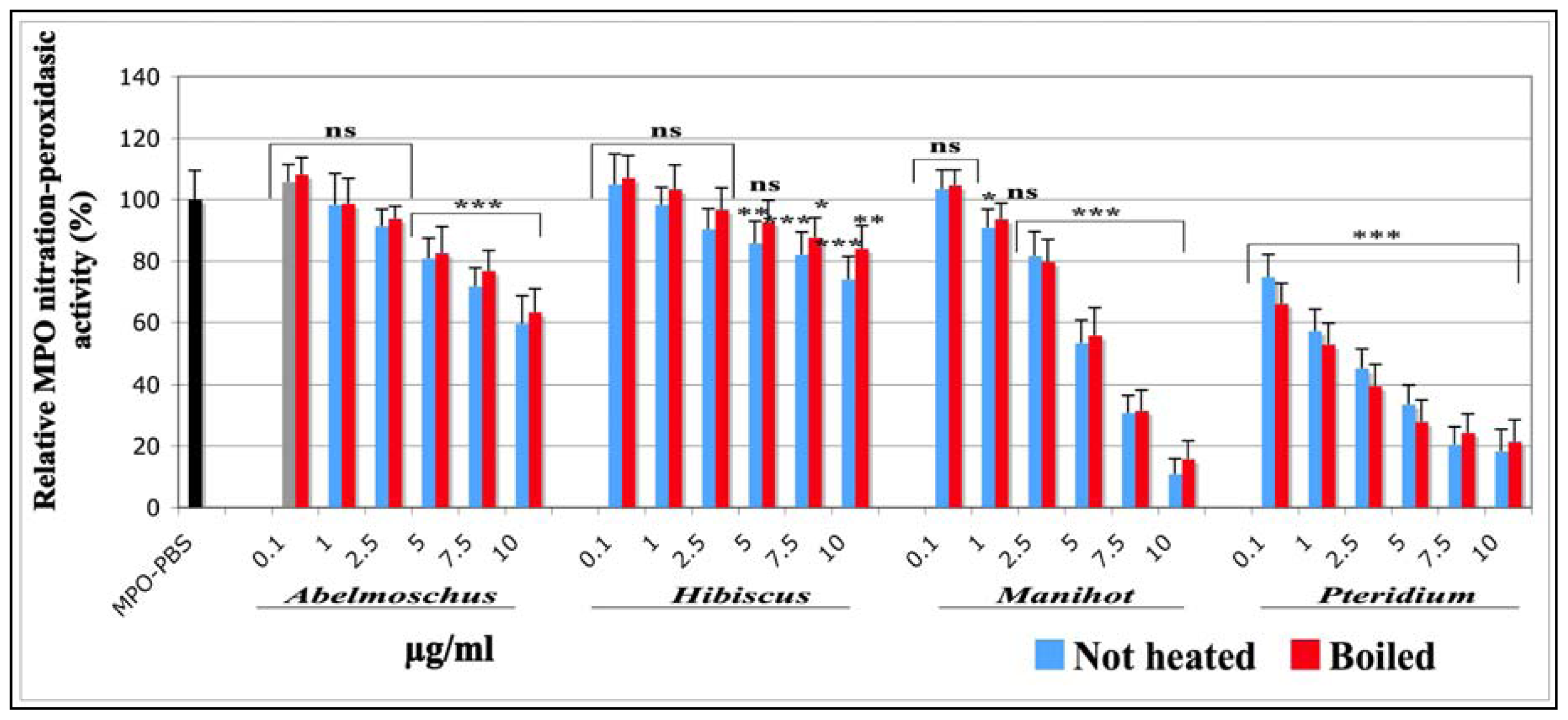
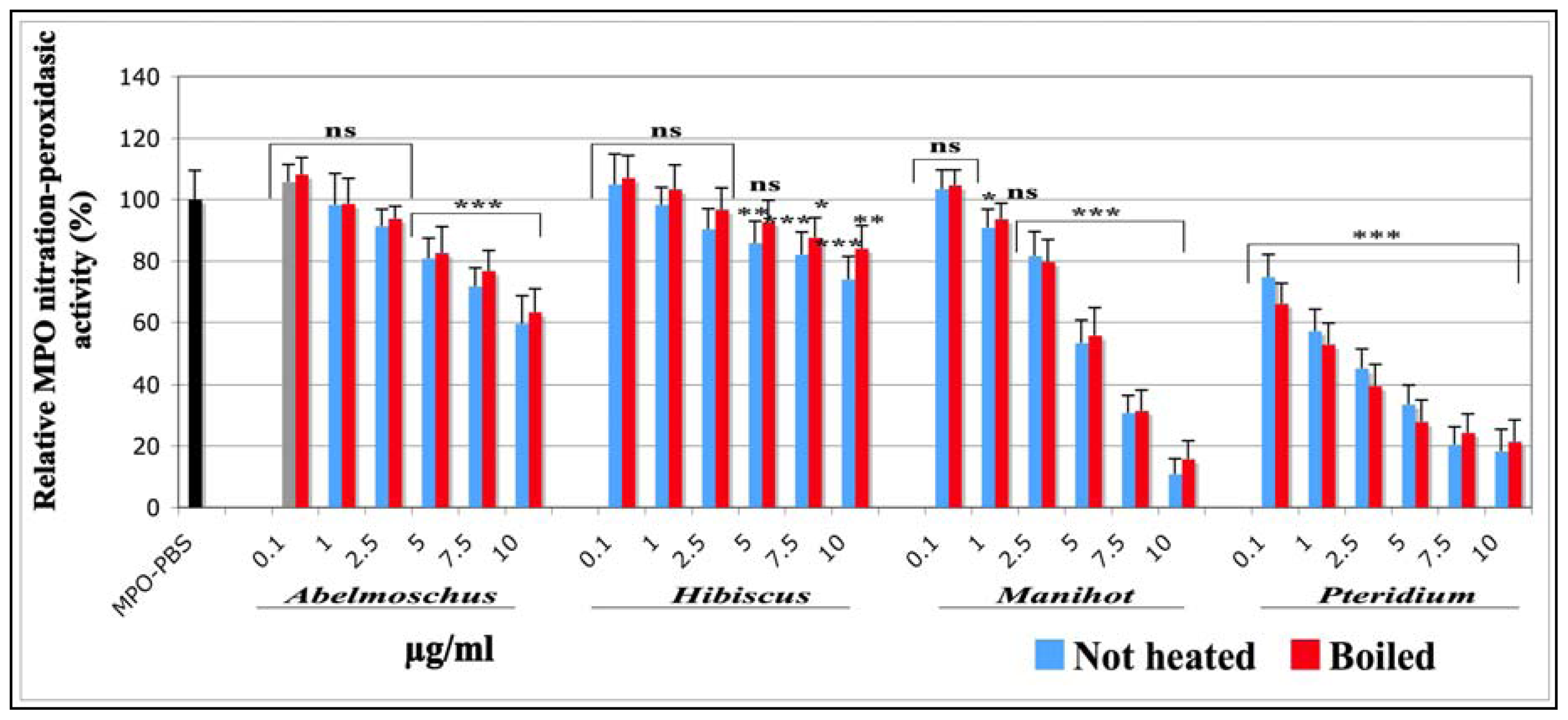
2.1.1.5. Effects on the Nitration-Peroxidasic Activity of MPO Measured by SIEFED Technique
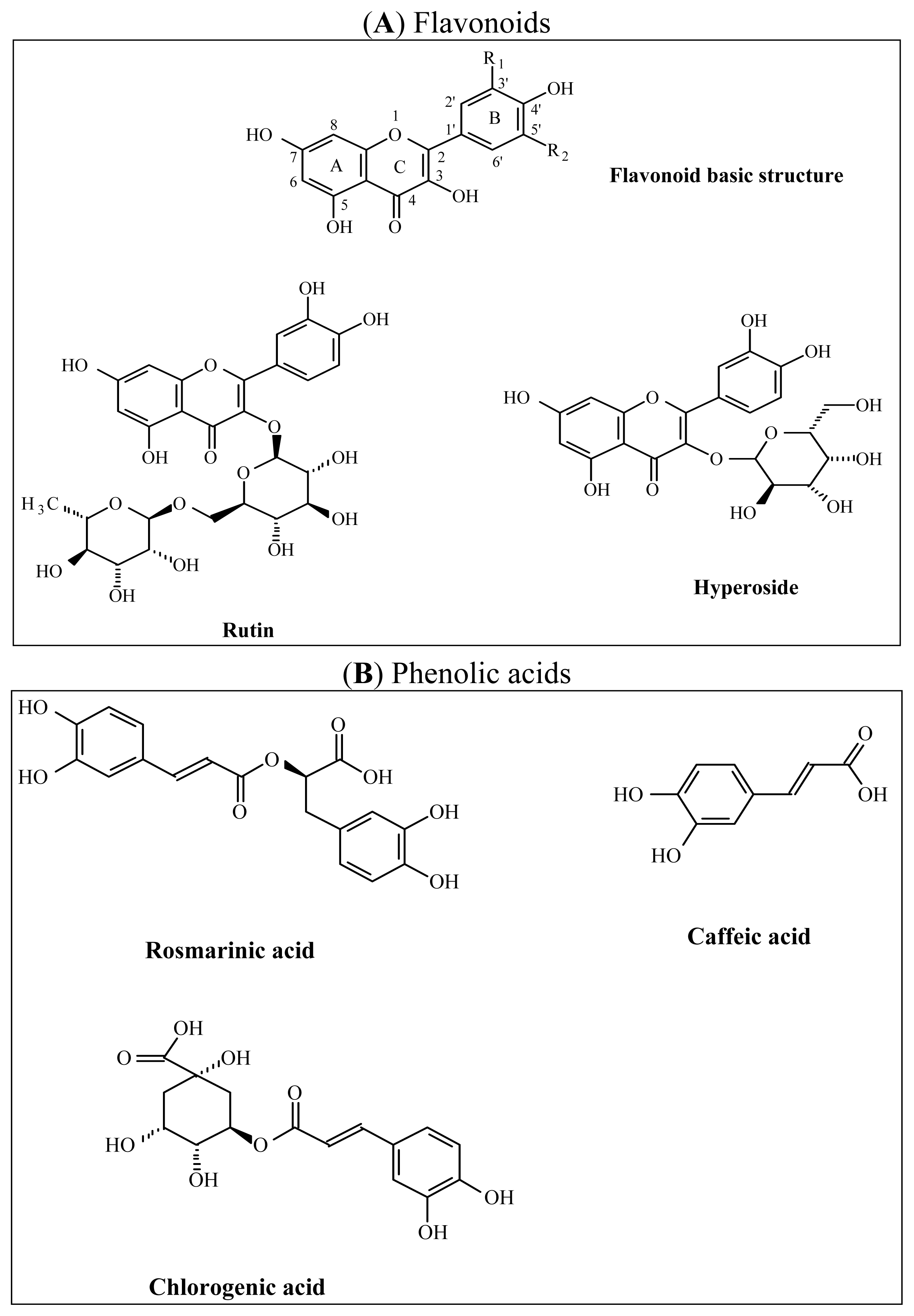
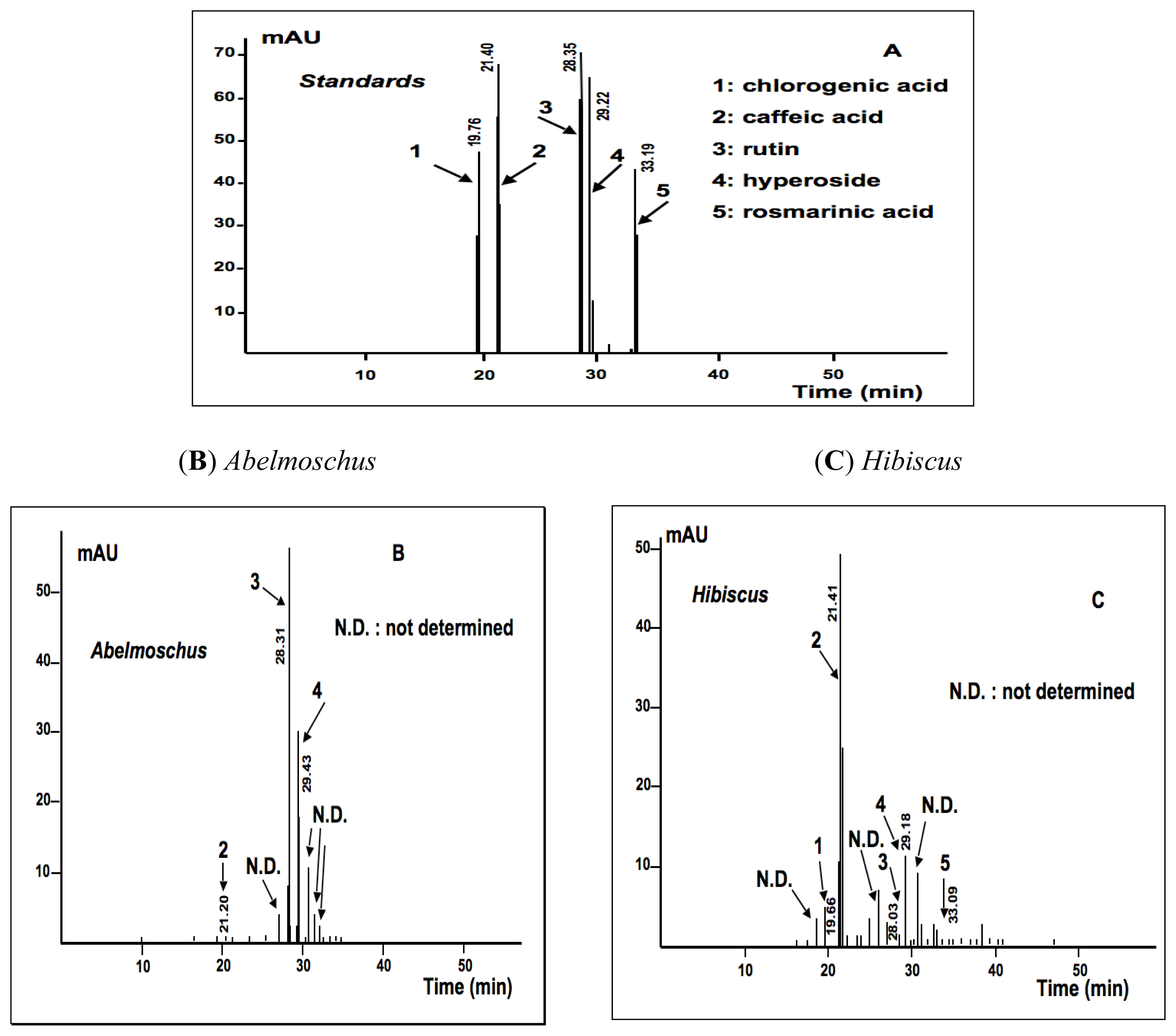
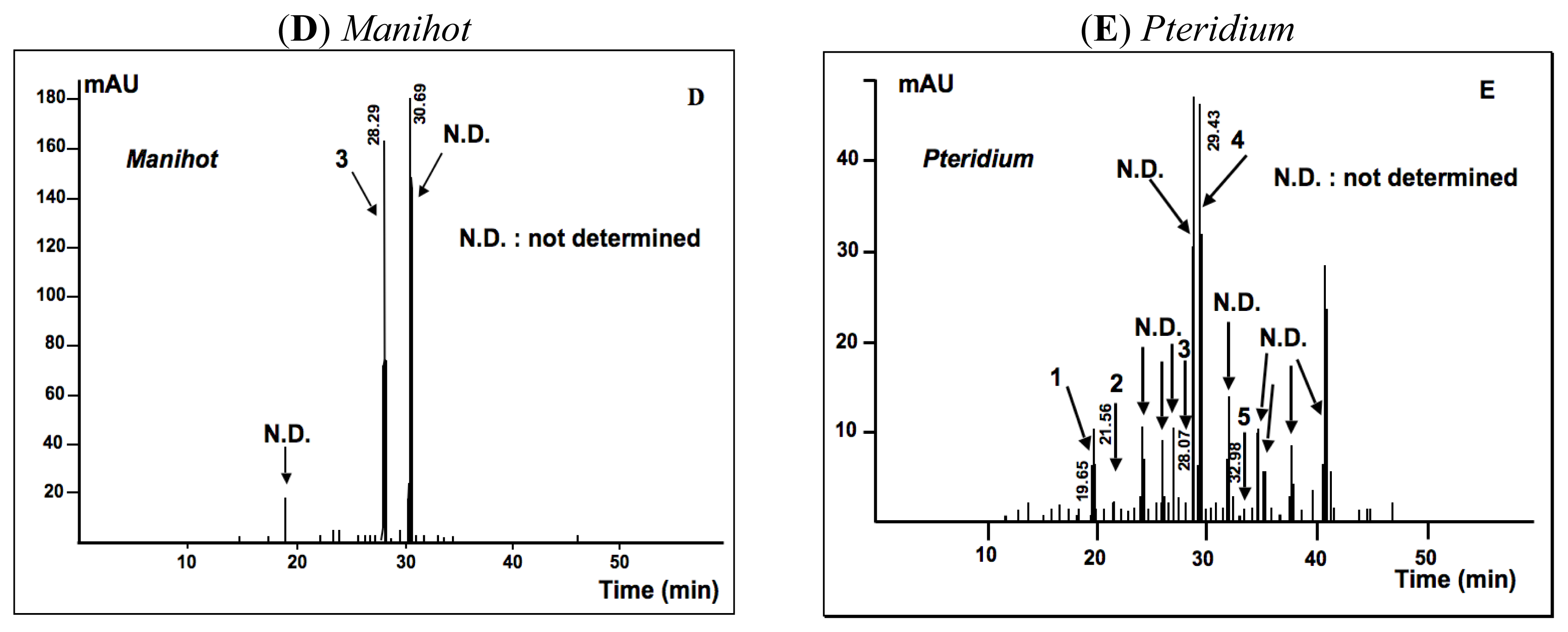
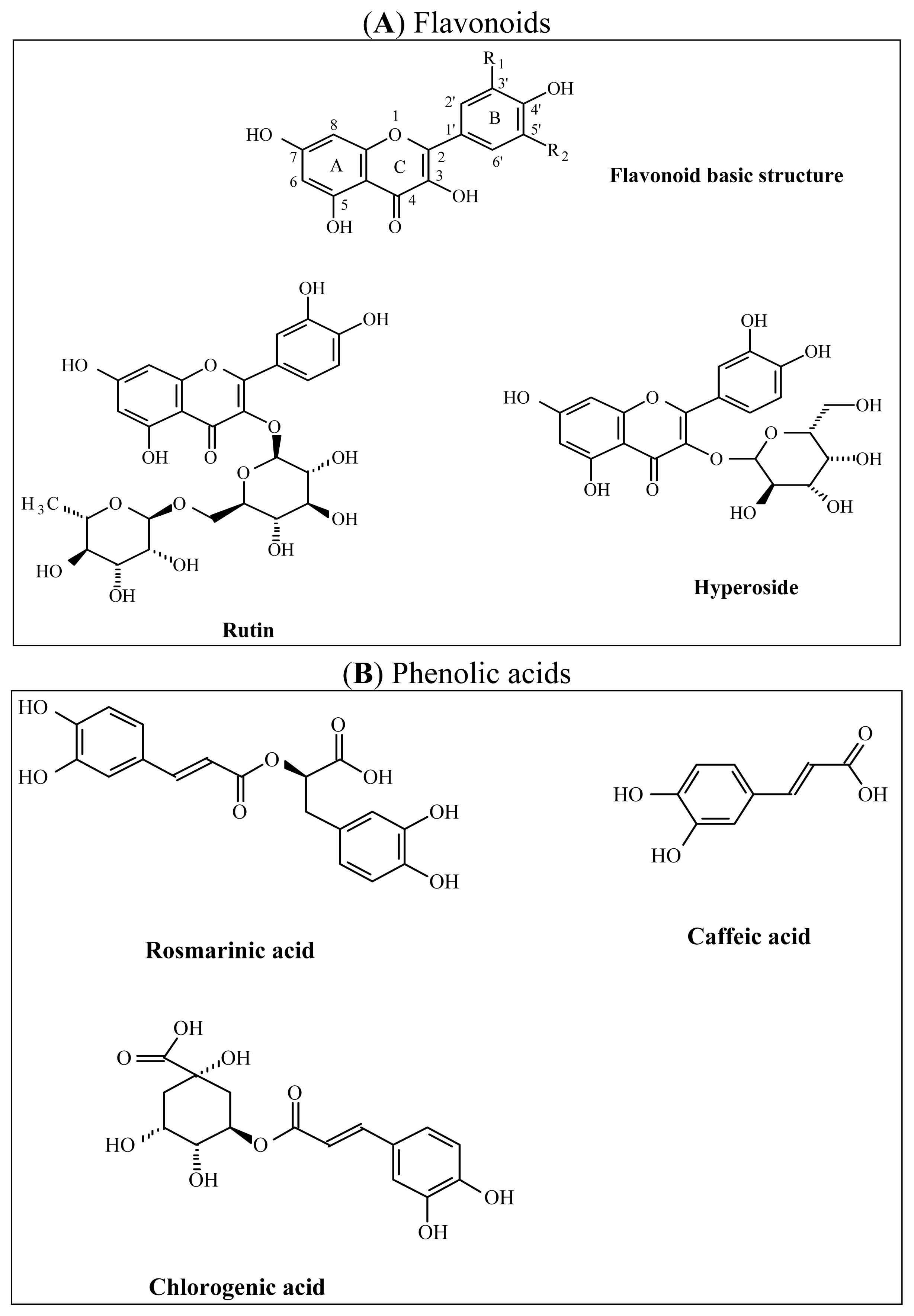
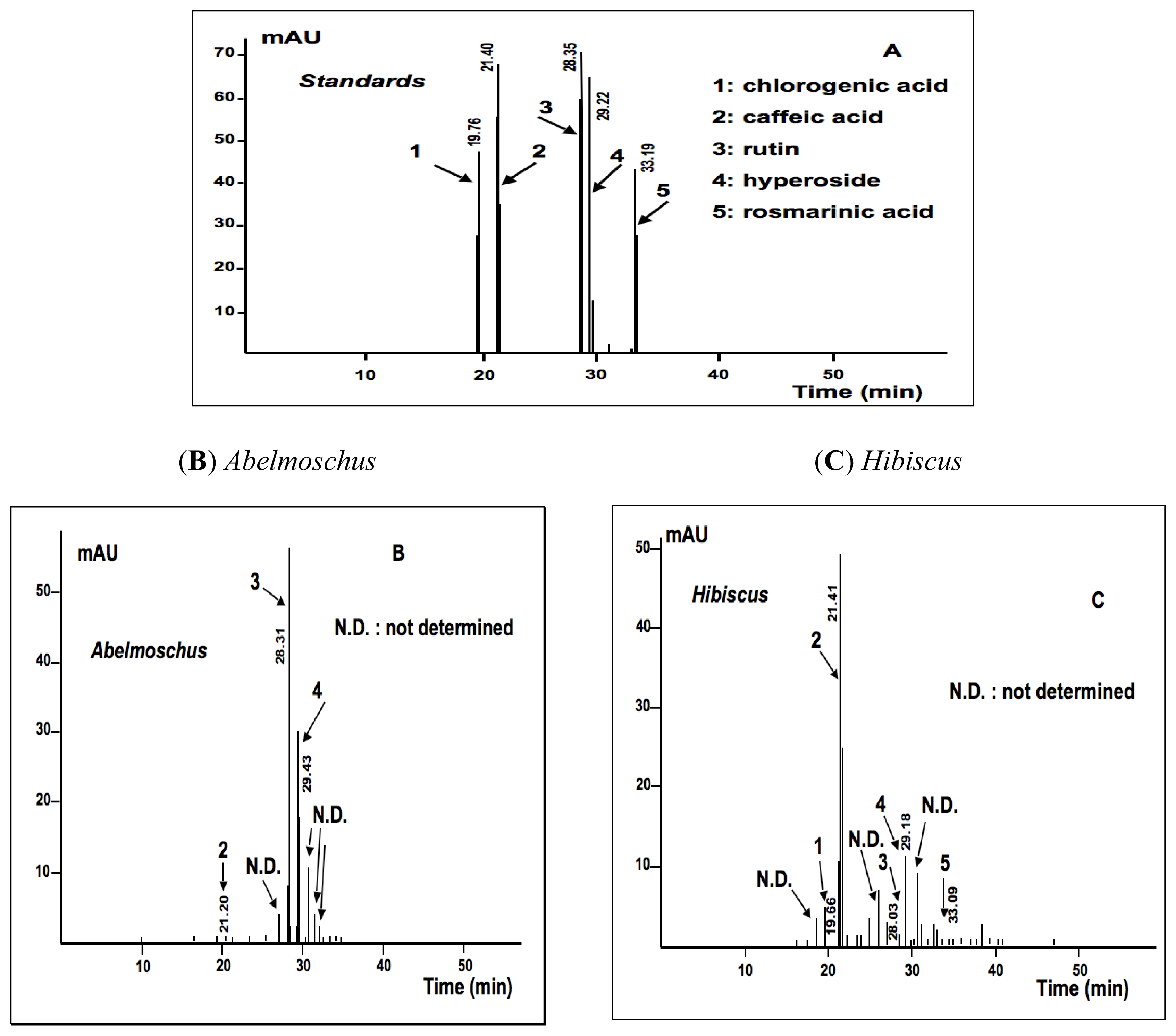
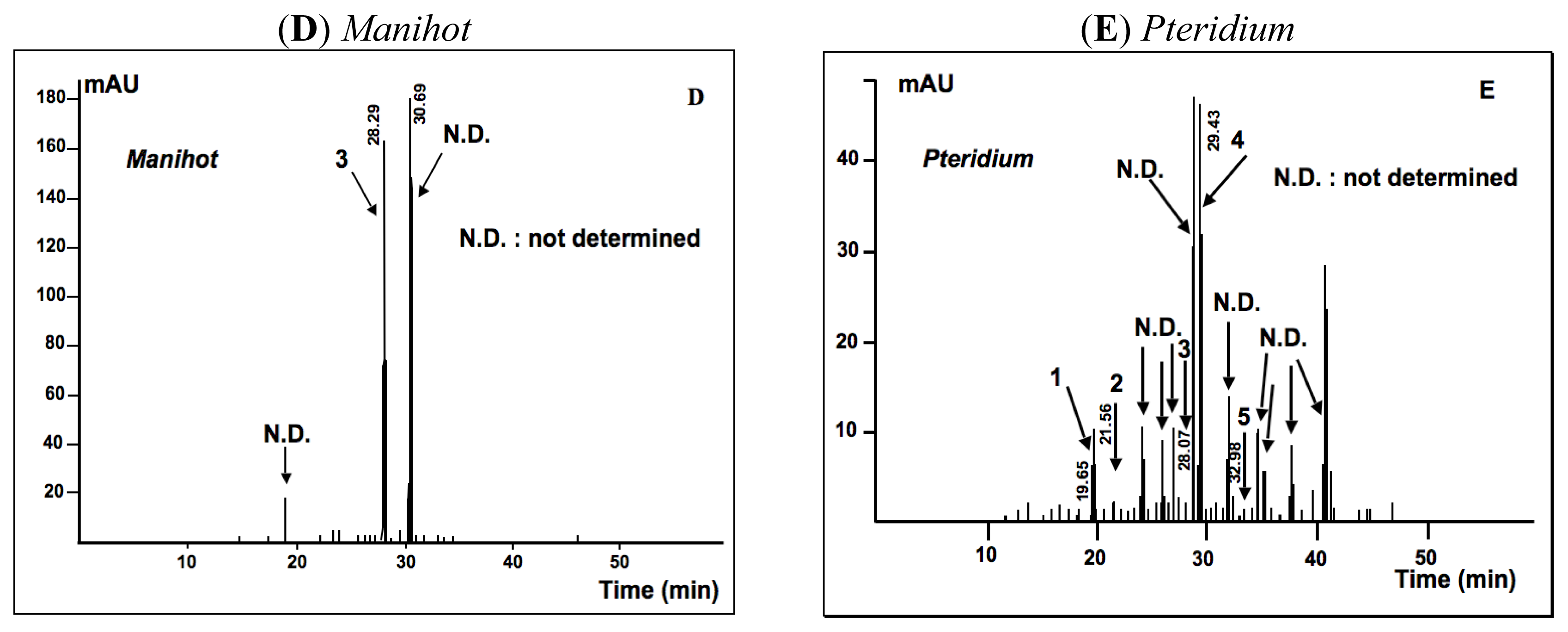
2.1.2. Phytochemical Analysis
2.2. Discussion
3. Experimental Section
3.1. Chemicals and Reagents
3.2. Vegetal Material
3.3. Preparation of the Plant Extracts for Biochemical Assays on Cell and Enzyme Models
3.4. Isolation of Equine Neutrophils
3.5. Preparation of the Neutrophil Activator and the Plant Extract Solution
3.6. Biochemical Investigations
3.6.1. Cell Viability Assay
3.6.2. Measurement of the Total ROS Produced by Neutrophils Activated with PMA- (CL Assay)
3.6.3. Measurement of Total MPO Released in the Extra-Cellular Milieu by Neutrophils Activated with PMA (MPO-ELISA Assay)
3.6.4. Measurement of the Nitration Activity of Myeloperoxidase in a Tyrosine Nitration Assay
3.6.5. Measurement of the Nitration-Peroxidasic Activity of Myeloperoxidase by SIEFED Technique
3.7. Phytochemical Analysis
3.7.1. Extract Preparation
3.7.2. Determination of Total Polyphenol in the Extracts
3.7.3. Determination of Tannins in the Extracts
3.7.4. HPLC-UV/DAD Determination of Total Flavonoid and Phenol Acid
3.8. Statistical Analysis
4. Conclusion
Acknowledgments
References
- Faurschou, M.; Borregaard, N. Neutrophil granules and secretory vesicles in inflammation. Microbes Infect 2003, 5, 1317–1327. [Google Scholar]
- Deby-Dupont, G.; Deby, C.; Lamy, M. Neutrophil myeloperoxidase revisited: it’s role in health and disease. Intensivmed. Notfallmed 1999, 36, 500–513. [Google Scholar]
- Serteyn, D.; Grulke, S.; Franck, T.; Mouithys-Mickalad, A.; Deby-Dupont, G. La myéloperoxydase des neutrophiles, une enzyme de défense aux capacités oxydantes. Ann. Méd. Vét 2003, 147, 79–93. [Google Scholar]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev 2008, 88, 1243–1276. [Google Scholar]
- Burner, U.; Furtmüller, P.G.; Kettle, A.J.; Koppenol, W.H.; Obinger, C. Mechanism of reaction of myeloperoxidase with nitrite. J. Biolog. Chem 2000, 275, 20597–20601. [Google Scholar]
- Kohnen, S.; Franck, T.; van Antwerpen, P.; Boudjeltia, K.Z.; Mouithys-Mickalad, A.; Deby, C.; Moguilevsky, N.; Deby-Dupont, G.; Lamy, M.; Serteyn, D. Resveratrol inhibits the activity of equine neutrophil myeloperoxidase by a direct interaction with the enzyme. J. Agric. Food Chem 2007, 55, 8080–8087. [Google Scholar]
- Griendling, K.K.; Sorescu, D.; Lassègue, B.; Ushio-Fukai, M. Modulation of protein kinase activity and gene expression by reactive oxygen species and their role in vascular physiology and pathophysiology. Arterioscler. Thromb. Vasc. Biol 2000, 20, 2175–2183. [Google Scholar]
- Valko, M.; Leibfritz, D.; Moncol, J.; Croninc, M. T. D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol 2007, 39, 44–84. [Google Scholar]
- Dröge, W. Free radicals in the physiological control of cell function. Physiol. Rev 2002, 82, 47–95. [Google Scholar]
- Federico, A.; Morgillo, F.; Tuccillo, C.; Ciardiello, F.; Loguercio, C. Chronic inflammation and oxidative stress in human carcinogenesis. Int. J. Cancer 2007, 121, 2381–2386. [Google Scholar]
- Klebanoff, S.J. Myeloperoxidase: friend and foe. J. Leukoc. Biol 2005, 77, 598–625. [Google Scholar]
- Malle, E.; Furtmüller, P.G.; Sattler, W.; Obinger, C. Myeloperoxidase: A target for new drug development? Br. J. Pharmacol 2007, 152, 838–854. [Google Scholar]
- Kim, J.A.; Neupane, G.P.; Lee, E.S.; Jeong, B.S.; Park, B.C.; Thapa, B.C. NADPH oxidase inhibitors: a patent review. Expert Opin. Ther. Pat 2011, 21, 1147–1158. [Google Scholar]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar]
- Heim, K.E.; Tagliaferro, A.R.; Bobilya, D.J. Flavonoid antioxidants: Chemistry, metabolism and structure–activity relationships. J. Nutr. Biochem 2002, 13, 572–584. [Google Scholar]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-antioxidant activity relationships of flavonoids and phenolic acids. Free Radical Biol. Med 1996, 20, 933–956. [Google Scholar]
- Seifried, H.E.; Anderson, D.E.; Fisher, E.I.; Milner, J.A. A review of the interaction among dietary antioxidants and reactive oxygen species. J. Nutr. Biochem 2007, 18, 567–579. [Google Scholar]
- Anand, P.; Kunnumakara, A.B.; Sundaram, C.; Harikumar, K.B.; Tharakan, S.T.; Lai, O.S.; Sung, B.; Aggarwal, B.B. Cancer is a preventable disease that requires major lifestyle changes. Pharm. Res 2008, 25, 2097–2116. [Google Scholar]
- Prior, R.L. Fruits and vegetables in the prevention of cellular oxidative damage. Am. J. Clin. Nutr 2003, 78, 570S–578S. [Google Scholar]
- Vauzour, D.; Rodriguez-Mateos, A.; Corona, G.; Oruna-Concha, M.J.; Spencer, J.P.E. Polyphenols and human health: prevention of disease and mechanisms of action. Nutrients 2010, 2, 1106–1131. [Google Scholar]
- Weisburger, J.H. Mechanisms of action of antioxidants as exemplified in vegetables, tomatoes and tea. Food Chem. Toxicol 1999, 3, 943–948. [Google Scholar]
- Almazan, A.M.; Theberge, R.L. Influence of cassava mosaic virus on cassava leaf-vegetable quality. Trop. Agric 1989, 66, 305–308. [Google Scholar]
- Kubo, I.; Masuoka, N.; Nihei, K.I.; Burgheim, B. Maniçoba, a quercetin-rich Amazona dish. J. Food Compos. Anal 2006, 19, 579–588. [Google Scholar]
- Miladiyah, I.; Dayi, F.; Desrini, S. Analgesic activity of ethanolic extract of Manihot esculenta Crantz leaves in mice. Universa Med 2011, 30, 3–10. [Google Scholar]
- Wright, C.I.; Van-Buren, L.; Kroner, C.I.; Koning, M.M.G. Herbal medicines as diuretics: a review of the scientific evidence. J. Ethnopharmacol 2007, 114, 1–31. [Google Scholar]
- Farombi, E.O. African indigenous plants with chemotherapeutic potentials and biotechnological approach to the production of bioactive prophylactic agents. Afr. J. Biotechnol 2003, 2, 662–671. [Google Scholar]
- Wong, S.P.; Leong, L.P.; Koh, J.H.W. Antioxidant activities of aqueous extracts of selected plants. Food Chem 2006, 99, 775–783. [Google Scholar]
- Suresh, R.; Saravanakumar, M.; Suganyadevi, P. Anthocyanins from Indian cassava (Manihot esculenta Crantz) and its antioxidant properties. Int. J. Pharm. Sci. Res 2011, 2, 1819–1828. [Google Scholar]
- Katsube, T.; Tabata, H.; Ohta, Y.; Yamasaki, Y.; Anuurad, E.; Shiwaku, K.; Yamana, Y. Screening of antioxidant activity in edible plant products: Comparison of low-density lipoprotein oxidation assay, DPPH radical scavenging assay, and Folin-Ciocalteu assay. J. Agric. Food Chem 2004, 52, 2391–2396. [Google Scholar]
- Khomsug, P.; Thongjaroenbuangam, W.; Pakdeenarong, N.; Suttajit, M.; Chantiratikul, P. Antioxidative activities and phenolic content of extracts from Okra (Abelmoschus esculentus L.). Res. J. Biologic. Sci 2010, 5, 310–313. [Google Scholar]
- Adelakun, O.E.; Oyelade, O.J.; Ade-Omowaye, B.I.O.; Adeyemi, I.A.; Van de Venter, M. Chemical composition and the antioxidative properties of Nigerian Okra seed (Abelmoschus esculentus Moench) Flour. Food Chem. Toxicol 2009, 47, 1123–1126. [Google Scholar]
- Yang, R.-Y.; Tsou, S.C.S.; Lee, T.-C.; Wu, W.-J.; Hanson, P.M.; Kuo, G.; Engle, L.M.; Lai, P.-Y. Distribution of 127 edible plant species for antioxidant activities by two assays. J. Sci. Food Agric 2006, 86, 2395–2403. [Google Scholar]
- Imperato, F. Kaempferol-7-O-Rhamnoside-4′-O-glucoside from Pteridium Aquilinum. Phytochemistry 1998, 47, 911–913. [Google Scholar]
- Mendez, J. Dihydrocynnamic acids in Pteridium Aquilinum. Food Chem 2005, 93, 251–252. [Google Scholar]
- Tsumbu, C.N.; Deby-Dupont, G.; Tits, M.; Angenot, L.; Franck, T.; Serteyn, D.; Mouithys-Mickalad, A. Antioxidant and antiradical activities of Manihot esculenta Crantz (Euphorbiaceae) leaves and other selected tropical green vegetables investigated on lipoperoxidation and PMA activated monocytes. Nutrients 2011, 3, 818–838. [Google Scholar]
- Tenant, J.R. Evaluation of the Trypan blue technique for determination of cell viability. Transplantation 1964, 2, 685–694. [Google Scholar]
- Franck, T.; Grulke, S.; Deby-Dupont, G.; Deby, C.; Duvivier, H.; Peters, F.; Serteyn, D. Development of an enzyme-linked immunosorbent assay for specific neutrophil myeloperoxidase measurement in blood. J. Vet. Diagn. Invest 2005, 17, 412–419. [Google Scholar]
- Franck, T.; Kohnen, S.; Deby-Dupont, G.; Grulke, S.; Deby, C.; Serteyn, D. A specific method for measurement of equine active myeloperoxidase in biological samples and in in vitro tests. J. Vet. Diagn. Invest 2006, 18, 326–334. [Google Scholar]
- Cox, A.J.; Jeng, A.Y.; Sharkey, N.A.; Blumberg, P.M.; Tauber, A.I. Activation of the human neutrophil nicotinamide adenine dinucleotide phosphate (NADPH)-Oxidase by protein kinase C. J. Clinic. Invest 1985, 76, 1932–1938. [Google Scholar]
- Calixto, J.B.; Otuki, M.F.; Santos, A.R.S. Anti-inflammatory compounds of plant origin. Part I. Action on arachidonic acid pathway, nitric oxide and nuclear factor kappa B (NF-kappa B). Planta Med 2003, 69, 973–983. [Google Scholar]
- Calixto, J.B.; Campos, M.M.; Otuki, M.F.; Santos, A.R.S. Anti-inflammatory compounds of plant origin. Part II. Modulation of pro-inflammatory cytokines, chemokines and adhesion molecules. Planta Med 2004, 70, 93–103. [Google Scholar]
- Caldefie-Chézet, F.; Walrand, S.; Moinard, C.; Tridon, A.; Chassagne, J.; Vasson, M.P. Is the neutrophil reactive oxygen species production measured by luminol and lucigenin chemiluminescence intra or extracellular? Comparison with DCFH-DA flow cytometry and cytochrome C reduction. Clin. Chim. Acta 2002, 319, 9–17. [Google Scholar]
- Yunbo, L.; Zhu, H.; Kuppusamy, P.; Roubaud, V.; Zweier, J.L.; Trush, M.A. Validation of lucigenin (Bis-N-methylacridinium) as a chemilumigenic probe for detecting superoxide anion radical production by enzymatic and cellular systems. J. Biol. Chem 1998, 273, 2015–2023. [Google Scholar]
- Ferriola, P.C.; Cody, V.; Middelton, E., Jr. Protein kinase C inhibition by plant flavonoids. Biochem. Pharmacol 1989, 38, 1617–1624. [Google Scholar]
- da Cunha, F.M.; Dunna, D.; Assreuy, J.; Buzzi, F.C.; Campos, M.M.; Calixto, J.B. Caffeic acid derivates: In vitro and in vivo anti-inflammatory properties. Free Radic. Res 2004, 38, 1241–1253. [Google Scholar]
- Kim, S.J.; Um, J.Y.; Lee, J.Y. Anti-inflammatory activity of hyperoside through the suppression of nuclear factor - κ B activation in mouse peritoneal macrophages. Am. J. Chin. Med 2011, 39, 171–181. [Google Scholar]
- de la Rebière de Pouyade, G.; Serteyn, D.; Deby-Dupont, G.; Franck, T. Method for co-purification of equine neutrophil elastase and myeloperoxidase from a limited blood volume. Res. Vet. Sci 2009, 87, 358–363. [Google Scholar]
- Gach, O.; Magne, J.; Franck, T.; Derochette, S.; Deby, G.; Serteyn, D.; Defraigne, J.O.; Lancelotti, P.; Legrand, V.; Pierard, L.A. Clinical significance of active myeloperoxidase in carotid atherosclerotic plaques. Int. J. Cardiol 2011. [Google Scholar] [CrossRef]
- Jog, N.R.; Rane, M.J.; Lominadze, G.; Luerman, G.C.; Ward, R.A.; McLeisch, K.R. The actin cytoskeleton regulates exocytosis of all neutrophils subsets. Am. J. Physiol. Cell Physiol 2007, 292, 1690–1700. [Google Scholar]
- Borregard, N.; Cowland, J.B. Granules of the human neutrophilic polymorphonuclear leukocytes. Blood 1997, 89, 3503–3521. [Google Scholar]
- Sheloum, L.; Bourich, H.; Tigrine, C.; Boudoukha, A. Anti-inflammatory effects of rutin on rat paw oedema and on neutrophils chemotaxis and degranulation. Exp. Toxicol. Pathol 2003, 54, 313–318. [Google Scholar]
- Soubhye, J.; Prévost, M.; Van Antwerpen, P.; Zouaoui, B.K.; Rousseau, A.; Furtmüller, P.G.; Obinger, C.; Vanhaeverbeek, M.; Ducobu, J.; Nève, J.; Gelbcke, M.; Dufrasne, F.O. Structure-based design, synthesis, and pharmacological evaluation of 3-(aminoalkyl)-5- fluoroindoles as myeloperoxidase inhibitors. J. Med. Chem 2010, 53, 8747–8759. [Google Scholar]
- Kirchner, T.; Flemmig, J.; Furtmüller, P.G.; Obinger, C.; Arnhold, J. (−)-Epicatechin enhances the chlorinating activity of human myeloperoxidase. Arch. Biochem. Biophys 2010, 495, 21–27. [Google Scholar]
- Franck, T.; Kohnen, S.; Grulke, S.; Neven, P.; Goutman, Y.; Peters, F.; Pirotte, B.; Deby-Dupont, G.; Serteyn, D. Inhibitory effect of curcuminoids and tetracurcuminoids on equine activated neutrophils and myeloperoxidase activity. Physiol. Res 2008, 57, 577–587. [Google Scholar]
- Boly, R.; Dessy, S.; Kohnen, S.; Kini, F.; Lompo, M.; Mouithuys-Mickalad, A.; Guissou, I.P.; Dubois, J.; Deby-Dupont, G.; Serteyn, D.; et al. Modulatory activities of Agelanthus dodoneifolius (Loranthaceae) extracts on stimulated equine neutrophils and myeloperoxidase activity. Int. J. Mol. Med 2011, 28, 261–270. [Google Scholar]
- Shiba, Y.; Kinoshita, T.; Chuman, H.; Taketani, Y.; Takeda, E.; Kato, Y.; Naito, M.; Kawabata, K.; Ishisaka, A.; Terao, J.; Kawai, Y. Flavonoids as substrates and inhibitors of myeloperoxidase: Molecular actions of aglycone and metabolites. Chem. Res. Toxicol 2008, 21, 1600–1609. [Google Scholar]
- Meotti, F.C.; Senthilmohan, R.; Harwood, D.T.; Missau, F.C.; Pizzolatti, M.G.; Kettle, A.J. Myricitrin as substrate and inhibitor of myeloperoxidase: Implications for the pharmacological effects of flavonoids. Free Radic. Biol. Med 2008, 44, 109–120. [Google Scholar]
- Westby, A. Cassava Utilization, Storage and Small-Scale Processing. In Cassava: Biology, Production and Utilization; Hahn, S.K., Tresh, J.M., Belloti, A.C., Eds.; CABI Publishing: New York, NY, USA, 2001; pp. 281–300. [Google Scholar]
- Montagnac, J.A.; Davis, C.R.; Tanumihardjo, S.A. Processing techniques to reduce toxicity and antinutrients of cassava for use as a staple food. Comp. Rev. Food Sci. Food Saf 2009, 8, 17–27. [Google Scholar]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem 2002, 50, 3010–3014. [Google Scholar]
- Jeong, S.M.; Kim, S.Y.; Kim, D.R.; Jo, S.C.; Nam, K.C.; Ahn, D.U.; Lee, S.C. Effect of heat treatment on the antioxidant activity of extracts of citrus peels. J. Agric. Food Chem 2004, 52, 3389–3393. [Google Scholar]
- Temitope, A.O.; Olufemi, A.G.; Alaba, F.T. Effect of heat treatment on antioxidant activity of some spices. Cont. J. Food Sci. Technol 2010, 4, 53–59. [Google Scholar]
- Pycock, J.F.; Allen, W.E.; Morris, T.H. Rapid, single-step isolation of equine neutrophils on a discontinuous Percoll density gradient. Res. Vet. Sci 1987, 42, 411–412. [Google Scholar]
- Benbarek, H.; Deby-Dupont, G.; Deby, C.; Caudron, I.; Mathy-Hartet, M.; Lamy, M.; Serteyn, D. Experimental model for the study by chemiluminescence of the activation of isolated equine leucocytes. Res. Vet. Sci 1996, 61, 59–64. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by average of Folin-Ciocalteu reagent. Methods Enzymol 1999, 299, 152–178. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (μg/mL) | ||||||
|---|---|---|---|---|---|---|
| Chemiluminescence | Tyrosine nitration | SIEFED technique | ||||
| not heated | boiled | not heated | boiled | not heated | boiled | |
| Abel | 10.40 ± 1.1 (0.901) | 10.77 ± 1.0 (0.942) | 10.06 ± 1.0 (0.962) | 10.92 ± 1.6 (0.913) | 13.57 ± 1.1 (0.834) | 14.71 ± 1.1 (0.824) |
| Hib | 1.70 ± 0.8 (0.890) | 2.23 ± 1.1 (0.897) | 8.77 ± 1.1 (0.851) | 9.81 ± 1.1 (0.889) | 28.72 ± 1.2 (0.806) | 30.55 ± 1.6 (0.579) |
| Man | 4.82 ± 1.0 (0.965) | 5.04 ± 1.0 (0.970) | 4.92 ± 1.0 (0.963) | 5.23 ± 1.0 (0.966) | 4.96 ± 1.0 (0.967) | 5.04 ± 1.0 (0.996) |
| Pter | 1.43 ± 0.8 (0.907) | 1.89 ± 0.8 (0.688) | 1.10 ± 0.7 (0.827 | 1.14 ± 0.4 (0.940) | 1.50 ± 0.8 (0.917) | 1.28 ± 0.7 (0.921) |
| Abelmoschus | Hibiscus | Manihot | Pteridium | |
|---|---|---|---|---|
| Total polyphenol a | 1,480 | 1,730 | N.D. | 6,740 |
| Tannin b | 600 | 500 | N.D. | 5,550 |
| Non-tannin | 880 | 1,230 | N.D. | 1,190 |
| Total Flavonoid c | 425 | 775 | 1,381 | 448 |
| Total phenolic acid d | N.D. | 975 | N.D. | 278 |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Tsumbu, C.N.; Deby-Dupont, G.; Tits, M.; Angenot, L.; Frederich, M.; Kohnen, S.; Mouithys-Mickalad, A.; Serteyn, D.; Franck, T. Polyphenol Content and Modulatory Activities of Some Tropical Dietary Plant Extracts on the Oxidant Activities of Neutrophils and Myeloperoxidase. Int. J. Mol. Sci. 2012, 13, 628-650. https://doi.org/10.3390/ijms13010628
Tsumbu CN, Deby-Dupont G, Tits M, Angenot L, Frederich M, Kohnen S, Mouithys-Mickalad A, Serteyn D, Franck T. Polyphenol Content and Modulatory Activities of Some Tropical Dietary Plant Extracts on the Oxidant Activities of Neutrophils and Myeloperoxidase. International Journal of Molecular Sciences. 2012; 13(1):628-650. https://doi.org/10.3390/ijms13010628
Chicago/Turabian StyleTsumbu, Cesar N., Ginette Deby-Dupont, Monique Tits, Luc Angenot, Michel Frederich, Stephane Kohnen, Ange Mouithys-Mickalad, Didier Serteyn, and Thierry Franck. 2012. "Polyphenol Content and Modulatory Activities of Some Tropical Dietary Plant Extracts on the Oxidant Activities of Neutrophils and Myeloperoxidase" International Journal of Molecular Sciences 13, no. 1: 628-650. https://doi.org/10.3390/ijms13010628
APA StyleTsumbu, C. N., Deby-Dupont, G., Tits, M., Angenot, L., Frederich, M., Kohnen, S., Mouithys-Mickalad, A., Serteyn, D., & Franck, T. (2012). Polyphenol Content and Modulatory Activities of Some Tropical Dietary Plant Extracts on the Oxidant Activities of Neutrophils and Myeloperoxidase. International Journal of Molecular Sciences, 13(1), 628-650. https://doi.org/10.3390/ijms13010628