Non-Coding RNAs in Retinal Development

Abstract
:
1. Introduction
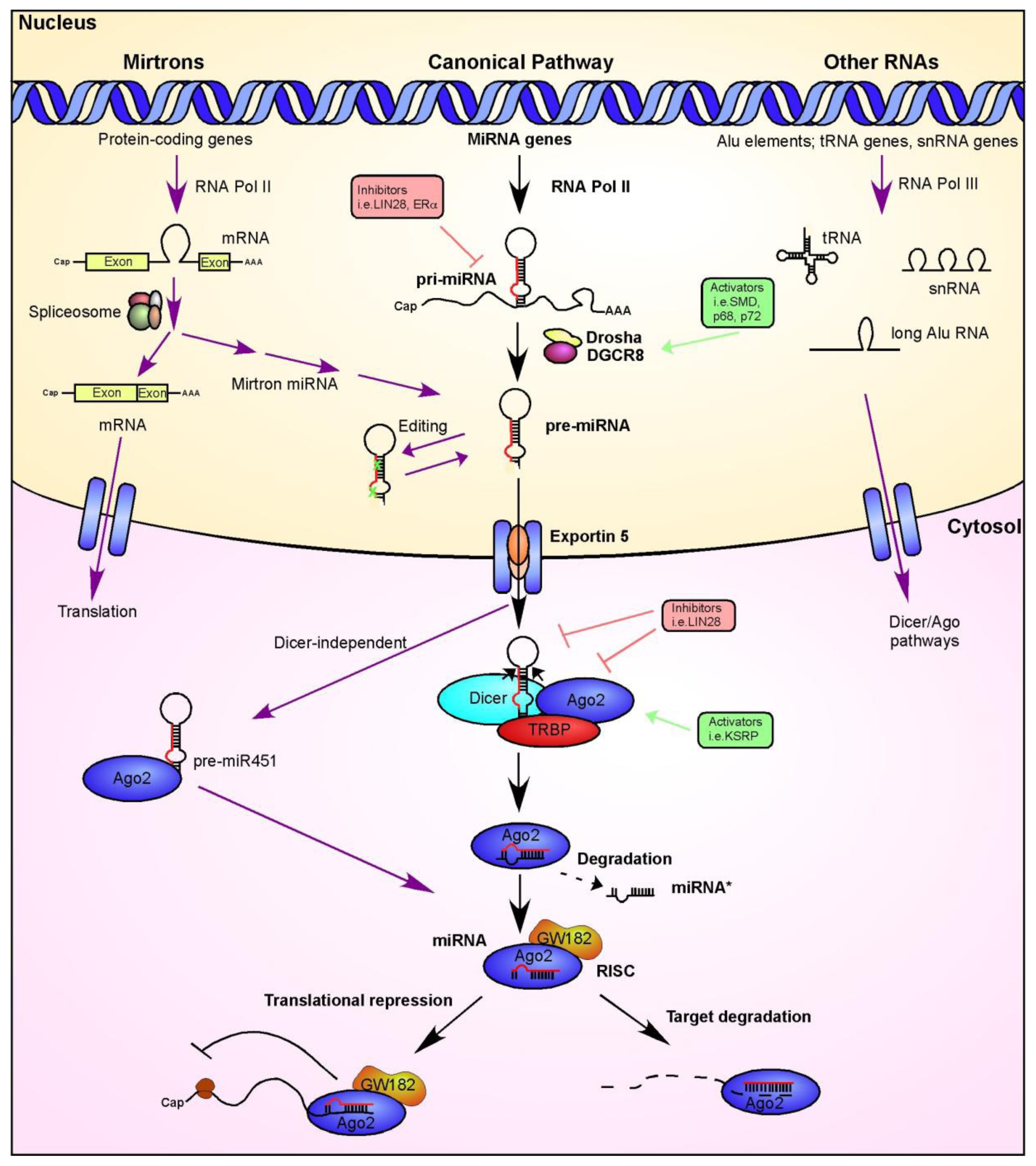
2. Identification of miRNAs in the Retina
3. Function of MicroRNAs in the Visual System
3.1. Dicer Deletions
3.2. Functions of Individual miRNAs in the Visual System
4. MicroRNAs and Retinal Diseases
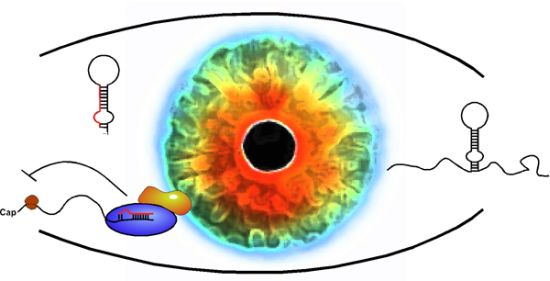
5. Long Non-Coding RNAs in the Visual System
6. Conclusion
Acknowledgements
- Conflict of InterestThe authors declare no conflict of interest.
References
- Chow, R.L.; Lang, R.A. Early eye development in vertebrates. Annu. Rev. Cell Dev. Biol 2001, 17, 255–296. [Google Scholar]
- Harada, T.; Harada, C.; Parada, L.F. Molecular regulation of visual system development: More than meets the eye. Genes Dev 2007, 21, 367–378. [Google Scholar]
- Marquardt, T.; Gruss, P. Generating neuronal diversity in the retina: One for nearly all. Trends Neurosci 2002, 25, 32–38. [Google Scholar]
- McLaughlin, T.; Hindges, R.; O’Leary, D.D. Regulation of axial patterning of the retina and its topographic mapping in the brain. Curr. Opin. Neurobiol 2003, 13, 57–69. [Google Scholar]
- Roska, B.; Werblin, F. Vertical interactions across ten parallel, stacked representations in the mammalian retina. Nature 2001, 410, 583–587. [Google Scholar]
- Amaral, P.P.; Mattick, J.S. Noncoding RNA in development. Mamm. Genome 2008, 19, 454–492. [Google Scholar]
- Cao, X.; Yeo, G.; Muotri, A.R.; Kuwabara, T.; Gage, F.H. Noncoding RNAs in the mammalian central nervous system. Annu. Rev. Neurosci 2006, 29, 77–103. [Google Scholar]
- Kosik, K.S. The neuronal microRNA system. Nat. Rev. Neurosci 2006, 7, 911–920. [Google Scholar]
- Carninci, P.; Kasukawa, T.; Katayama, S.; Gough, J.; Frith, M.C.; Maeda, N.; Oyama, R.; Ravasi, T.; Lenhard, B.; Wells, C.; et al. The transcriptional landscape of the mammalian genome. Science 2005, 309, 1559–1563. [Google Scholar]
- Bartel, D.P. Micrornas: Genomics, biogenesis, mechanism, and function. Cell 2004, 116, 281–297. [Google Scholar]
- Krol, J.; Loedige, I.; Filipowicz, W. The widespread regulation of microRNA biogenesis, function and decay. Nat. Rev. Genet 2010, 11, 597–610. [Google Scholar]
- Baek, D.; Villen, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of microRNAs on protein output. Nature 2008, 455, 64–71. [Google Scholar]
- Selbach, M.; Schwanhausser, B.; Thierfelder, N.; Fang, Z.; Khanin, R.; Rajewsky, N. Widespread changes in protein synthesis induced by microRNAs. Nature 2008, 455, 58–63. [Google Scholar]
- Giraldez, A.J.; Mishima, Y.; Rihel, J.; Grocock, R.J.; van Dongen, S.; Inoue, K.; Enright, A.J.; Schier, A.F. Zebrafish miR-430 promotes deadenylation and clearance of maternal mRNAs. Science 2006, 312, 75–79. [Google Scholar]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J.; Bartel, D.P.; Linsley, P.S.; Johnson, J.M. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar]
- Giraldez, A.J.; Cinalli, R.M.; Glasner, M.E.; Enright, A.J.; Thomson, J.M.; Baskerville, S.; Hammond, S.M.; Bartel, D.P.; Schier, A.F. MicroRNAs regulate brain morphogenesis in zebrafish. Science 2005, 308, 833–838. [Google Scholar]
- Harfe, B.D.; McManus, M.T.; Mansfield, J.H.; Hornstein, E.; Tabin, C.J. The RNaseIII enzyme dicer is required for morphogenesis but not patterning of the vertebrate limb. Proc. Natl. Acad. Sci. USA 2005, 102, 10898–10903. [Google Scholar]
- Furuno, M.; Pang, K.C.; Ninomiya, N.; Fukuda, S.; Frith, M.C.; Bult, C.; Kai, C.; Kawai, J.; Carninci, P.; Hayashizaki, Y.; et al. Clusters of internally primed transcripts reveal novel long noncoding RNAs. PLoS Genet 2006, 2. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet 2009, 10, 155–159. [Google Scholar]
- Ponjavic, J.; Oliver, P.L.; Lunter, G.; Ponting, C.P. Genomic and transcriptional co-localization of protein-coding and long non-coding RNA pairs in the developing brain. PLoS Genet 2009, 5. [Google Scholar] [CrossRef]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P.; et al. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar]
- Guttman, M.; Donaghey, J.; Carey, B.W.; Garber, M.; Grenier, J.K.; Munson, G.; Young, G.; Lucas, A.B.; Ach, R.; Bruhn, L.; et al. LincRNAs act in the circuitry controlling pluripotency and differentiation. Nature 2011, 477, 295–300. [Google Scholar]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermuller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar]
- Pedersen, J.S.; Bejerano, G.; Siepel, A.; Rosenbloom, K.; Lindblad-Toh, K.; Lander, E.S.; Kent, J.; Miller, W.; Haussler, D. Identification and classification of conserved RNA secondary structures in the human genome. PLoS Comput. Biol 2006, 2. [Google Scholar] [CrossRef]
- Washietl, S.; Hofacker, I.L.; Lukasser, M.; Huttenhofer, A.; Stadler, P.F. Mapping of conserved RNA secondary structures predicts thousands of functional noncoding RNAs in the human genome. Nat. Biotechnol 2005, 23, 1383–1390. [Google Scholar]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell 2011, 43, 904–914. [Google Scholar]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; van Oudenaarden, A.; et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar]
- Whitehead, J.; Pandey, G.K.; Kanduri, C. Regulation of the mammalian epigenome by long noncoding RNAs. Biochim. Biophys. Acta 2009, 1790, 936–947. [Google Scholar]
- Amaral, P.P.; Clark, M.B.; Gascoigne, D.K.; Dinger, M.E.; Mattick, J.S. LncRNAdb: A reference database for long noncoding RNAs. Nucleic Acids Res 2011, 39, D146–D151. [Google Scholar]
- Rapicavoli, N.A.; Blackshaw, S. New meaning in the message: Noncoding RNAs and their role in retinal development. Dev. Dyn 2009, 238, 2103–2114. [Google Scholar]
- Berezikov, E.; Cuppen, E.; Plasterk, R.H. Approaches to microRNA discovery. Nat. Genet 2006, 38, S2–S7. [Google Scholar]
- Creighton, C.J.; Reid, J.G.; Gunaratne, P.H. Expression profiling of microRNAs by deep sequencing. Brief. Bioinform 2009, 10, 490–497. [Google Scholar]
- Xie, X.; Lu, J.; Kulbokas, E.J.; Golub, T.R.; Mootha, V.; Lindblad-Toh, K.; Lander, E.S.; Kellis, M. Systematic discovery of regulatory motifs in human promoters and 3′ UTRs by comparison of several mammals. Nature 2005, 434, 338–345. [Google Scholar]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A.; Pfeffer, S.; Rice, A.; Kamphorst, A.O.; Landthaler, M.; et al. A mammalian microRNA expression atlas based on small RNA library sequencing. Cell 2007, 129, 1401–1414. [Google Scholar]
- Kloosterman, W.P.; Steiner, F.A.; Berezikov, E.; de Bruijn, E.; van de Belt, J.; Verheul, M.; Cuppen, E.; Plasterk, R.H. Cloning and expression of new microRNAs from zebrafish. Nucleic Acids Res 2006, 34, 2558–2569. [Google Scholar]
- Berezikov, E.; Guryev, V.; van de Belt, J.; Wienholds, E.; Plasterk, R.H.; Cuppen, E. Phylogenetic shadowing and computational identification of human microRNA genes. Cell 2005, 120, 21–24. [Google Scholar]
- Griffiths-Jones, S.; Saini, H.K.; van Dongen, S.; Enright, A.J. Mirbase: Tools for microRNA genomics. Nucleic Acids Res 2008, 36, D154–D158. [Google Scholar]
- Kozomara, A.; Griffiths-Jones, S. Mirbase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res 2011, 39, D152–D157. [Google Scholar]
- Lagos-Quintana, M.; Rauhut, R.; Meyer, J.; Borkhardt, A.; Tuschl, T. New microRNAs from mouse and human. RNA 2003, 9, 175–179. [Google Scholar]
- Wienholds, E.; Kloosterman, W.P.; Miska, E.; Alvarez-Saavedra, E.; Berezikov, E.; de Bruijn, E.; Horvitz, H.R.; Kauppinen, S.; Plasterk, R.H. MicroRNA expression in zebrafish embryonic development. Science 2005, 309, 310–311. [Google Scholar]
- Arora, A.; McKay, G.J.; Simpson, D.A. Prediction and verification of miRNA expression in human and rat retinas. Invest. Ophthalmol. Vis. Sci 2007, 48, 3962–3967. [Google Scholar]
- Makarev, E.; Spence, J.R.; del Rio-Tsonis, K.; Tsonis, P.A. Identification of microRNAs and other small RNAs from the adult newt eye. Mol. Vis 2006, 12, 1386–1391. [Google Scholar]
- Xu, S.; Witmer, P.D.; Lumayag, S.; Kovacs, B.; Valle, D. MicroRNA (miRNA) transcriptome of mouse retina and identification of a sensory organ-specific mirna cluster. J. Biol. Chem 2007, 282, 25053–25066. [Google Scholar]
- Kapsimali, M.; Kloosterman, W.P.; de Bruijn, E.; Rosa, F.; Plasterk, R.H.; Wilson, S.W. MicroRNAs show a wide diversity of expression profiles in the developing and mature central nervous system. Genome Biol 2007, 8, R173. [Google Scholar]
- Karali, M.; Peluso, I.; Marigo, V.; Banfi, S. Identification and characterization of microRNAs expressed in the mouse eye. Invest. Ophthalmol. Vis. Sci 2007, 48, 509–515. [Google Scholar]
- Ryan, D.G.; Oliveira-Fernandes, M.; Lavker, R.M. MicroRNAs of the mammalian eye display distinct and overlapping tissue specificity. Mol. Vis 2006, 12, 1175–1184. [Google Scholar]
- Xu, S. MicroRNA expression in the eyes and their significance in relation to functions. Prog. Retin. Eye Res 2009, 28, 87–116. [Google Scholar]
- Hackler, L., Jr; Wan, J.; Swaroop, A.; Qian, J.; Zack, D.J. MicroRNA profile of the developing mouse retina. Invest. Ophthalmol. Vis. Sci 2010, 51, 1823–1831. [Google Scholar]
- Weston, M.D.; Pierce, M.L.; Rocha-Sanchez, S.; Beisel, K.W.; Soukup, G.A. MicroRNA gene expression in the mouse inner ear. Brain Res 2006, 1111, 95–104. [Google Scholar]
- Kloosterman, W.P.; Wienholds, E.; de Bruijn, E.; Kauppinen, S.; Plasterk, R.H. In situ detection of miRNAs in animal embryos using lna-modified oligonucleotide probes. Nat. Methods 2006, 3, 27–29. [Google Scholar]
- Pierce, M.L.; Weston, M.D.; Fritzsch, B.; Gabel, H.W.; Ruvkun, G.; Soukup, G.A. MicroRNA-183 family conservation and ciliated neurosensory organ expression. Evol. Dev 2008, 10, 106–113. [Google Scholar]
- Loscher, C.J.; Hokamp, K.; Kenna, P.F.; Ivens, A.C.; Humphries, P.; Palfi, A.; Farrar, G.J. Altered retinal microRNA expression profile in a mouse model of retinitis pigmentosa. Genome Biol 2007, 8. [Google Scholar] [CrossRef]
- Karali, M.; Peluso, I.; Gennarino, V.A.; Bilio, M.; Verde, R.; Lago, G.; Dolle, P.; Banfi, S. MiRNeye: A microRNA expression atlas of the mouse eye. BMC Genomics 2010, 11. [Google Scholar] [CrossRef]
- Schratt, G. MicroRNAs at the synapse. Nat. Rev. Neurosci 2009, 10, 842–849. [Google Scholar]
- Stefani, G.; Slack, F.J. Small non-coding RNAs in animal development. Nat. Rev. Mol. Cell Biol 2008, 9, 219–230. [Google Scholar]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.Org resource: Targets and expression. Nucleic Acids Res 2008, 36, D149–D153. [Google Scholar]
- Friedman, R.C.; Farh, K.K.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res 2009, 19, 92–105. [Google Scholar]
- Krek, A.; Grun, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet 2005, 37, 495–500. [Google Scholar]
- Bernstein, E.; Kim, S.Y.; Carmell, M.A.; Murchison, E.P.; Alcorn, H.; Li, M.Z.; Mills, A.A.; Elledge, S.J.; Anderson, K.V.; Hannon, G.J. Dicer is essential for mouse development. Nat. Genet 2003, 35, 215–217. [Google Scholar]
- Damiani, D.; Alexander, J.J.; O’Rourke, J.R.; McManus, M.; Jadhav, A.P.; Cepko, C.L.; Hauswirth, W.W.; Harfe, B.D.; Strettoi, E. Dicer inactivation leads to progressive functional and structural degeneration of the mouse retina. J. Neurosci 2008, 28, 4878–4887. [Google Scholar]
- Rowan, S.; Cepko, C.L. Genetic analysis of the homeodomain transcription factor chx10 in the retina using a novel multifunctional bac transgenic mouse reporter. Dev. Biol 2004, 271, 388–402. [Google Scholar]
- Lin, S.C.; Skapek, S.X.; Papermaster, D.S.; Hankin, M.; Lee, E.Y. The proliferative and apoptotic activities of e2f1 in the mouse retina. Oncogene 2001, 20, 7073–7084. [Google Scholar]
- de Pietri Tonelli, D.; Pulvers, J.N.; Haffner, C.; Murchison, E.P.; Hannon, G.J.; Huttner, W.B. MiRNAs are essential for survival and differentiation of newborn neurons but not for expansion of neural progenitors during early neurogenesis in the mouse embryonic neocortex. Development 2008, 135, 3911–3921. [Google Scholar]
- Pinter, R.; Hindges, R. Perturbations of microRNA function in mouse dicer mutants produce retinal defects and lead to aberrant axon pathfinding at the optic chiasm. PLoS One 2010, 5. [Google Scholar] [CrossRef]
- Soukup, G.A.; Fritzsch, B.; Pierce, M.L.; Weston, M.D.; Jahan, I.; McManus, M.T.; Harfe, B.D. Residual microRNA expression dictates the extent of inner ear development in conditional dicer knockout mice. Dev. Biol 2009, 328, 328–341. [Google Scholar]
- Decembrini, S.; Andreazzoli, M.; Barsacchi, G.; Cremisi, F. Dicer inactivation causes heterochronic retinogenesis in xenopus laevis. Int. J. Dev. Biol 2008, 52, 1099–1103. [Google Scholar]
- Georgi, S.A.; Reh, T.A. Dicer is required for the transition from early to late progenitor state in the developing mouse retina. J. Neurosci 2010, 30, 4048–4061. [Google Scholar]
- Marquardt, T.; Ashery-Padan, R.; Andrejewski, N.; Scardigli, R.; Guillemot, F.; Gruss, P. Pax6 is required for the multipotent state of retinal progenitor cells. Cell 2001, 105, 43–55. [Google Scholar]
- Georgi, S.A.; Reh, T.A. Dicer is required for the maintenance of notch signaling and gliogenic competence during mouse retinal development. Dev. Neurobiol 2011, 71, 1153–1169. [Google Scholar]
- Davis, N.; Mor, E.; Ashery-Padan, R. Roles for dicer1 in the patterning and differentiation of the optic cup neuroepithelium. Development 2011, 138, 127–138. [Google Scholar]
- Erskine, L.; Herrera, E. The retinal ganglion cell axon’s journey: Insights into molecular mechanisms of axon guidance. Dev. Biol 2007, 308, 1–14. [Google Scholar]
- Feldheim, D.A.; O’Leary, D.D. Visual map development: Bidirectional signaling, bifunctional guidance molecules, and competition. Cold Spring Harb. Perspect. Biol 2010, 2. [Google Scholar] [CrossRef]
- Swindell, E.C.; Bailey, T.J.; Loosli, F.; Liu, C.; Amaya-Manzanares, F.; Mahon, K.A.; Wittbrodt, J.; Jamrich, M. Rx-cre, a tool for inactivation of gene expression in the developing retina. Genesis 2006, 44, 361–363. [Google Scholar]
- Mathers, P.H.; Grinberg, A.; Mahon, K.A.; Jamrich, M. The Rx homeobox gene is essential for vertebrate eye development. Nature 1997, 387, 603–607. [Google Scholar]
- Harris, K.; Zhang, Z.; McManus, M.T.; Harfe, B.D.; Sun, X. Dicer function is essential for lung epithelium morphogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 2208–2213. [Google Scholar]
- Ho, J.; Ng, K.H.; Rosen, S.; Dostal, A.; Gregory, R.I.; Kreidberg, J.A. Podocyte-specific loss of functional microRNAs leads to rapid glomerular and tubular injury. J. Am. Soc. Nephrol 2008, 19, 2069–2075. [Google Scholar]
- Kersigo, J.; D’Angelo, A.; Gray, B.D.; Soukup, G.A.; Fritzsch, B. The role of sensory organs and the forebrain for the development of the craniofacial shape as revealed by foxg1-cre-mediated microRNA loss. Genesis 2011, 49, 326–341. [Google Scholar]
- Hebert, J.M.; McConnell, S.K. Targeting of cre to the foxg1 (bf-1) locus mediates loxp recombination in the telencephalon and other developing head structures. Dev. Biol 2000, 222, 296–306. [Google Scholar]
- Iida, A.; Shinoe, T.; Baba, Y.; Mano, H.; Watanabe, S. Dicer plays essential roles for retinal development by regulation of survival and differentiation. Invest. Ophthalmol. Vis. Sci 2011, 52, 3008–3017. [Google Scholar]
- Davis, N.; Yoffe, C.; Raviv, S.; Antes, R.; Berger, J.; Holzmann, S.; Stoykova, A.; Overbeek, P.A.; Tamm, E.R.; Ashery-Padan, R. Pax6 dosage requirements in iris and ciliary body differentiation. Dev. Biol 2009, 333, 132–142. [Google Scholar]
- Sage, C.; Huang, M.; Vollrath, M.A.; Brown, M.C.; Hinds, P.W.; Corey, D.P.; Vetter, D.E.; Chen, Z.Y. Essential role of retinoblastoma protein in mammalian hair cell development and hearing. Proc. Natl. Acad. Sci. USA 2006, 103, 7345–7350. [Google Scholar]
- Sato, S.; Inoue, T.; Terada, K.; Matsuo, I.; Aizawa, S.; Tano, Y.; Fujikado, T.; Furukawa, T. Dkk3-cre BAC transgenic mouse line: A tool for highly efficient gene deletion in retinal progenitor cells. Genesis 2007, 45, 502–507. [Google Scholar]
- Kaneko, H.; Dridi, S.; Tarallo, V.; Gelfand, B.D.; Fowler, B.J.; Cho, W.G.; Kleinman, M.E.; Ponicsan, S.L.; Hauswirth, W.W.; Chiodo, V.A.; et al. Dicer1 deficit induces Alu RNA toxicity in age-related macular degeneration. Nature 2011, 471, 325–330. [Google Scholar]
- Chong, M.M.; Rasmussen, J.P.; Rudensky, A.Y.; Littman, D.R. The rnaseiii enzyme drosha is critical in t cells for preventing lethal inflammatory disease. J. Exp. Med 2008, 205, 2005–2017. [Google Scholar]
- Yi, R.; Pasolli, H.A.; Landthaler, M.; Hafner, M.; Ojo, T.; Sheridan, R.; Sander, C.; O’Carroll, D.; Stoffel, M.; Tuschl, T.; et al. Dgcr8-dependent microRNA biogenesis is essential for skin development. Proc. Natl. Acad. Sci. USA 2009, 106, 498–502. [Google Scholar]
- O’Carroll, D.; Mecklenbrauker, I.; Das, P.P.; Santana, A.; Koenig, U.; Enright, A.J.; Miska, E.A.; Tarakhovsky, A. A slicer-independent role for argonaute 2 in hematopoiesis and the microRNA pathway. Genes Dev 2007, 21, 1999–2004. [Google Scholar]
- Chow, R.L.; Volgyi, B.; Szilard, R.K.; Ng, D.; McKerlie, C.; Bloomfield, S.A.; Birch, D.G.; McInnes, R.R. Control of late off-center cone bipolar cell differentiation and visual signaling by the homeobox gene vsx1. Proc. Natl. Acad. Sci. USA 2004, 101, 1754–1759. [Google Scholar]
- Decembrini, S.; Andreazzoli, M.; Vignali, R.; Barsacchi, G.; Cremisi, F. Timing the generation of distinct retinal cells by homeobox proteins. PLoS Biol 2006, 4. [Google Scholar] [CrossRef]
- Viczian, A.S.; Vignali, R.; Zuber, M.E.; Barsacchi, G.; Harris, W.A. Xotx5b and xotx2 regulate photoreceptor and bipolar fates in the xenopus retina. Development 2003, 130, 1281–1294. [Google Scholar]
- Decembrini, S.; Bressan, D.; Vignali, R.; Pitto, L.; Mariotti, S.; Rainaldi, G.; Wang, X.; Evangelista, M.; Barsacchi, G.; Cremisi, F. MicroRNAs couple cell fate and developmental timing in retina. Proc. Natl. Acad. Sci. USA 2009, 106, 21179–21184. [Google Scholar]
- Qiu, R.; Liu, K.; Liu, Y.; Mo, W.; Flynt, A.S.; Patton, J.G.; Kar, A.; Wu, J.Y.; He, R. The role of miR-124a in early development of the xenopus eye. Mech. Dev 2009, 126, 804–816. [Google Scholar]
- Walker, J.C.; Harland, R.M. MicroRNA-24a is required to repress apoptosis in the developing neural retina. Genes Dev 2009, 23, 1046–1051. [Google Scholar]
- Jin, Z.B.; Hirokawa, G.; Gui, L.; Takahashi, R.; Osakada, F.; Hiura, Y.; Takahashi, M.; Yasuhara, O.; Iwai, N. Targeted deletion of miR-182, an abundant retinal microRNA. Mol. Vis 2009, 15, 523–533. [Google Scholar]
- Krol, J.; Busskamp, V.; Markiewicz, I.; Stadler, M.B.; Ribi, S.; Richter, J.; Duebel, J.; Bicker, S.; Fehling, H.J.; Schubeler, D.; et al. Characterizing light-regulated retinal microRNAs reveals rapid turnover as a common property of neuronal microRNAs. Cell 2010, 141, 618–631. [Google Scholar]
- Bhattacharyya, S.N.; Habermacher, R.; Martine, U.; Closs, E.I.; Filipowicz, W. Relief of microRNA-mediated translational repression in human cells subjected to stress. Cell 2006, 125, 1111–1124. [Google Scholar]
- Tognini, P.; Putignano, E.; Coatti, A.; Pizzorusso, T. Experience-dependent expression of miR-132 regulates ocular dominance plasticity. Nat. Neurosci 2011, 14, 1237–1239. [Google Scholar]
- Mellios, N.; Sugihara, H.; Castro, J.; Banerjee, A.; Le, C.; Kumar, A.; Crawford, B.; Strathmann, J.; Tropea, D.; Levine, S.S.; et al. MiR-132, an experience-dependent microRNA, is essential for visual cortex plasticity. Nat. Neurosci 2011, 14, 1240–1242. [Google Scholar]
- Kloosterman, W.P.; Plasterk, R.H. The diverse functions of microRNAs in animal development and disease. Dev. Cell 2006, 11, 441–450. [Google Scholar]
- Sayed, D.; Abdellatif, M. MicroRNAs in development and disease. Physiol. Rev 2011, 91, 827–887. [Google Scholar]
- Eacker, S.M.; Dawson, T.M.; Dawson, V.L. Understanding microRNAs in neurodegeneration. Nat. Rev. Neurosci 2009, 10, 837–841. [Google Scholar]
- Loscher, C.J.; Hokamp, K.; Wilson, J.H.; Li, T.; Humphries, P.; Farrar, G.J.; Palfi, A. A common microRNA signature in mouse models of retinal degeneration. Exp. Eye Res 2008, 87, 529–534. [Google Scholar]
- Shen, J.; Yang, X.; Xie, B.; Chen, Y.; Swaim, M.; Hackett, S.F.; Campochiaro, P.A. MicroRNAs regulate ocular neovascularization. Mol. Ther 2008, 16, 1208–1216. [Google Scholar]
- Mattick, J.S.; Taft, R.J.; Faulkner, G.J. A global view of genomic information--moving beyond the gene and the master regulator. Trends Genet 2010, 26, 21–28. [Google Scholar]
- Mercer, T.R.; Dinger, M.E.; Sunkin, S.M.; Mehler, M.F.; Mattick, J.S. Specific expression of long noncoding RNAs in the mouse brain. Proc. Natl. Acad. Sci. USA 2008, 105, 716–721. [Google Scholar]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet 2006, 15, R17–R29. [Google Scholar]
- Nagano, T.; Fraser, P. No-nonsense functions for long noncoding RNAs. Cell 2011, 145, 178–181. [Google Scholar]
- Alfano, G.; Vitiello, C.; Caccioppoli, C.; Caramico, T.; Carola, A.; Szego, M.J.; McInnes, R.R.; Auricchio, A.; Banfi, S. Natural antisense transcripts associated with genes involved in eye development. Hum. Mol. Genet 2005, 14, 913–923. [Google Scholar]
- Blackshaw, S.; Harpavat, S.; Trimarchi, J.; Cai, L.; Huang, H.; Kuo, W.P.; Weber, G.; Lee, K.; Fraioli, R.E.; Cho, S.H.; et al. Genomic analysis of mouse retinal development. PLoS Biol 2004, 2. [Google Scholar] [CrossRef]
- Geng, X.; Lavado, A.; Lagutin, O.V.; Liu, W.; Oliver, G. Expression of six3 opposite strand (six3os) during mouse embryonic development. Gene Expr. Patterns 2007, 7, 252–257. [Google Scholar]
- Meola, N.; Pizzo, M.; Alfano, G.; Surace, E.M.; Banfi, S. The long noncoding RNA vax2os1 controls the cell cycle progression of photoreceptor progenitors in the mouse retina. RNA 2012, 18, 111–123. [Google Scholar]
- Feng, J.; Bi, C.; Clark, B.S.; Mady, R.; Shah, P.; Kohtz, J.D. The evf-2 noncoding RNA is transcribed from the dlx-5/6 ultraconserved region and functions as a dlx-2 transcriptional coactivator. Genes Dev 2006, 20, 1470–1484. [Google Scholar]
- Rapicavoli, N.A.; Poth, E.M.; Zhu, H.; Blackshaw, S. The long noncoding RNA six3os acts in trans to regulate retinal development by modulating six3 activity. Neural Dev 2011, 6. [Google Scholar] [CrossRef]
- Bernstein, E.; Duncan, E.M.; Masui, O.; Gil, J.; Heard, E.; Allis, C.D. Mouse polycomb proteins bind differentially to methylated histone h3 and RNA and are enriched in facultative heterochromatin. Mol. Cell. Biol 2006, 26, 2560–2569. [Google Scholar]
- Sanchez-Elsner, T.; Gou, D.; Kremmer, E.; Sauer, F. Noncoding RNAs of trithorax response elements recruit drosophila ash1 to ultrabithorax. Science 2006, 311, 1118–1123. [Google Scholar]
- Martens, J.A.; Wu, P.Y.; Winston, F. Regulation of an intergenic transcript controls adjacent gene transcription in saccharomyces cerevisiae. Genes Dev 2005, 19, 2695–2704. [Google Scholar]
- Ho, Y.; Elefant, F.; Liebhaber, S.A.; Cooke, N.E. Locus control region transcription plays an active role in long-range gene activation. Mol. Cell 2006, 23, 365–375. [Google Scholar]
- Young, T.L.; Matsuda, T.; Cepko, C.L. The noncoding RNA taurine upregulated gene 1 is required for differentiation of the murine retina. Curr. Biol 2005, 15, 501–512. [Google Scholar]
- Rapicavoli, N.A.; Poth, E.M.; Blackshaw, S. The long noncoding RNA rncr2 directs mouse retinal cell specification. BMC Dev. Biol 2010, 10. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Cre-line | Onset of Creexpression | Location of Cre-expression (visual system) | Survival (homozygotes) | Reported phenotypes | Reference |
|---|---|---|---|---|---|
| Chx10-cre [61] | E10.5 | Retinal progenitors, mosaic | Normal | Photoreceptor rosettes, apoptosis, abnormal ERG responses | Damiani et al., 2008, [60] |
| Rx-cre [73] | E7.5 | Eye-forming tissues, anterior neural plate | Die at P0 | Micropthalmia, apoptosis, axon guidance defects | Pinter & Hindges 2010, [64] |
| Pax6-α-cre [68] | E10.5 | Nasal and temporal distal regions of the retina | Normal | Micropthalmia, apoptosis, increase of early retinal cells, decrease of late progenitors | Georgi & Reh, 2010, [67] |
| Detached iris pigmented epithelium, hypoplastic ciliary body | Davis et al., 2011, [70] | ||||
| Tyrp2-cre [80] | E9 | From E11 in pigmented retinal cells, at late embryogenesis in presumptive pigmented epithelia of the ciliary body and the iris and in the muscles/stroma of iris; (detectable from E9 onwards in eye, forebrain and DRGs); | Poor at adult stages | Microphthalmia, most iris tissues missing, hypoplastic ciliary body | Davis et al., 2011, [70] |
| Pou4f3-cre [81] | P1 | Non-pigmented ciliary body and iris pigmented epithelium | Die soon after birth (most) | Detached iris pigmented epithelia, hyperplastic ciliary body | Davis et al., 2011, [70] |
| Foxg1-cre [78] | E8.5 | Optic vesicle, lens, telencephalon, olfactory epithelium, ear, foregut, isthmus | Die just before birth | Microphtalamia mostly associated with high apoptosis and depigmentation in nasal retina, lens missing. | Kersigo et al., 2011, [77] |
| Dkk3-cre [82] | E10.5 | Retinal progenitors | Die 4–6 weeks postnatal | Micropthalmia, apoptosis, defects in retinal cell differentiation | Iida et al., 2011, [79] |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Maiorano, N.A.; Hindges, R. Non-Coding RNAs in Retinal Development. Int. J. Mol. Sci. 2012, 13, 558-578. https://doi.org/10.3390/ijms13010558
Maiorano NA, Hindges R. Non-Coding RNAs in Retinal Development. International Journal of Molecular Sciences. 2012; 13(1):558-578. https://doi.org/10.3390/ijms13010558
Chicago/Turabian StyleMaiorano, Nicola A., and Robert Hindges. 2012. "Non-Coding RNAs in Retinal Development" International Journal of Molecular Sciences 13, no. 1: 558-578. https://doi.org/10.3390/ijms13010558