Impact of Temporary Nitrogen Deprivation on Tomato Leaf Phenolics

Abstract
:
1. Introduction
2. Results
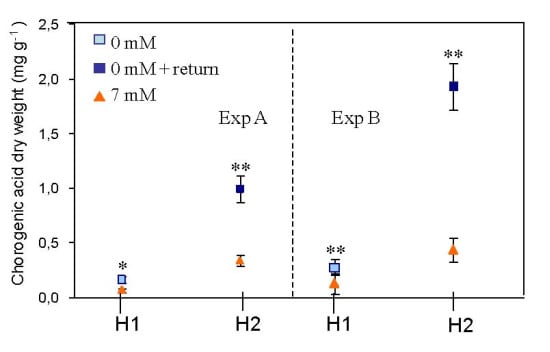
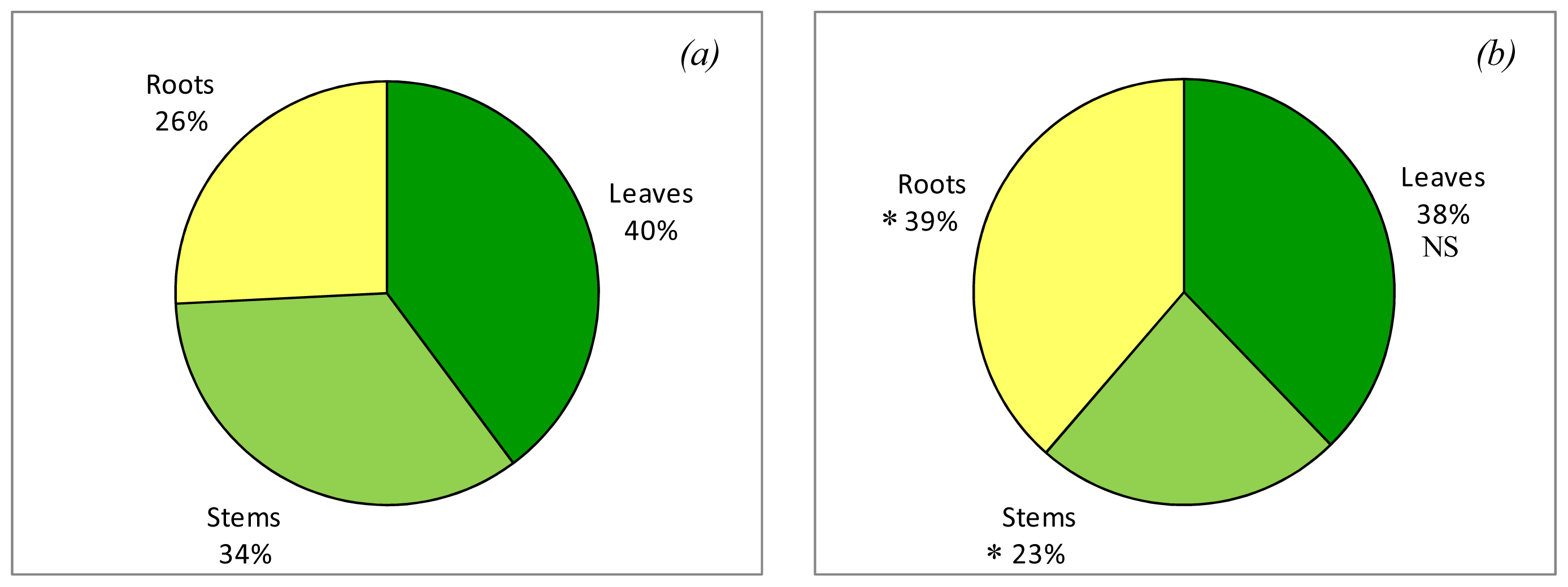
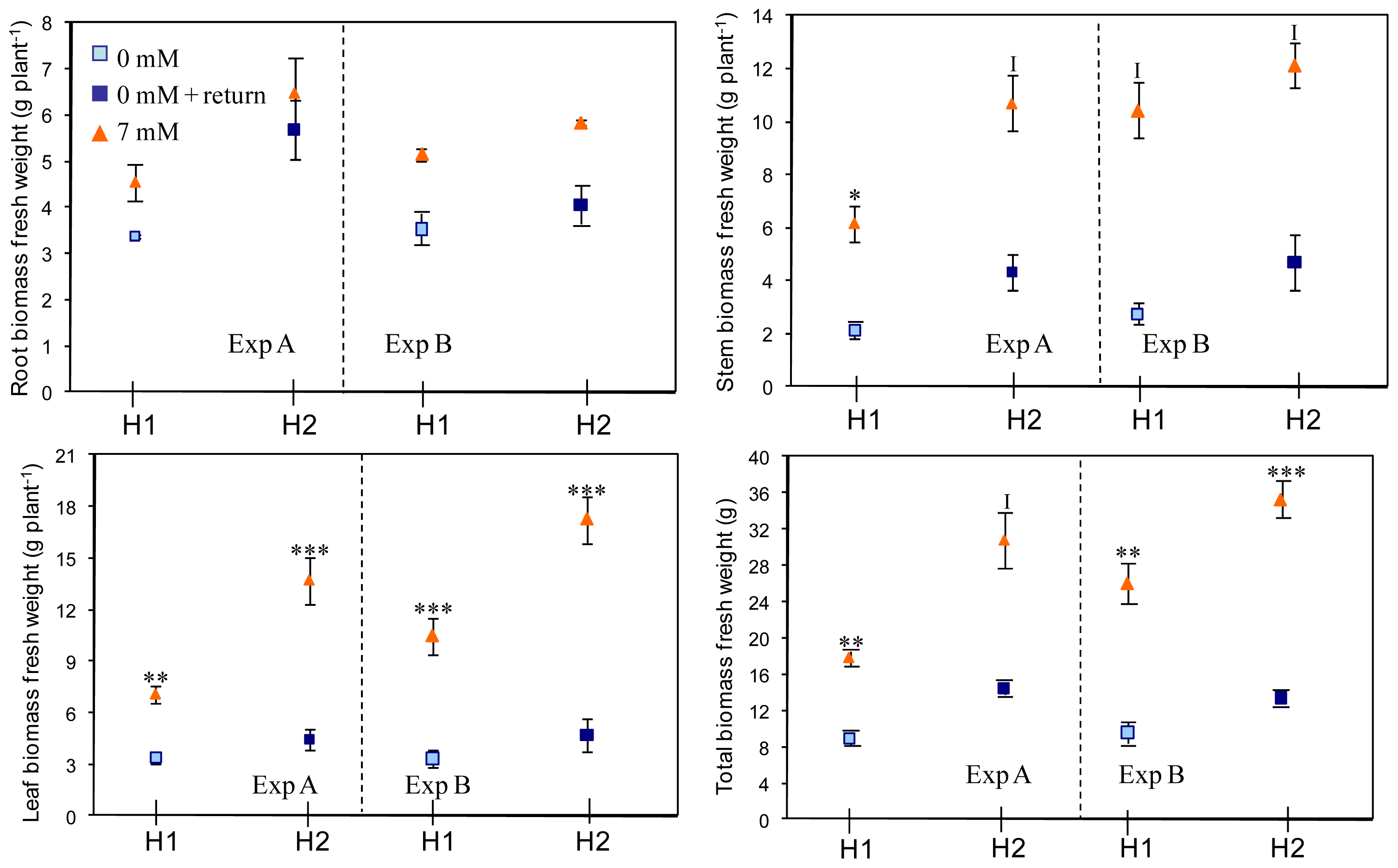
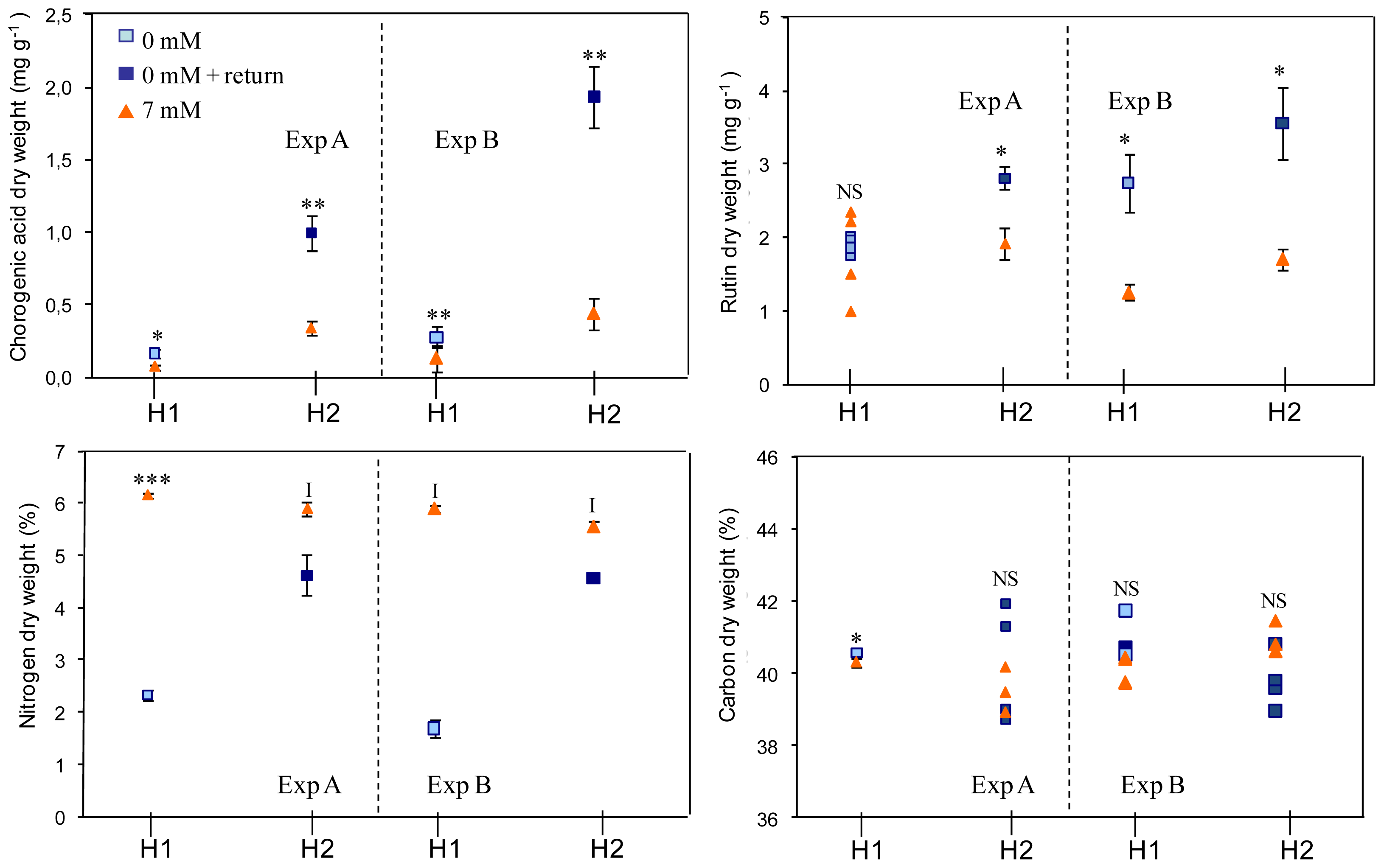
2.1. Effects on Plant Biomass
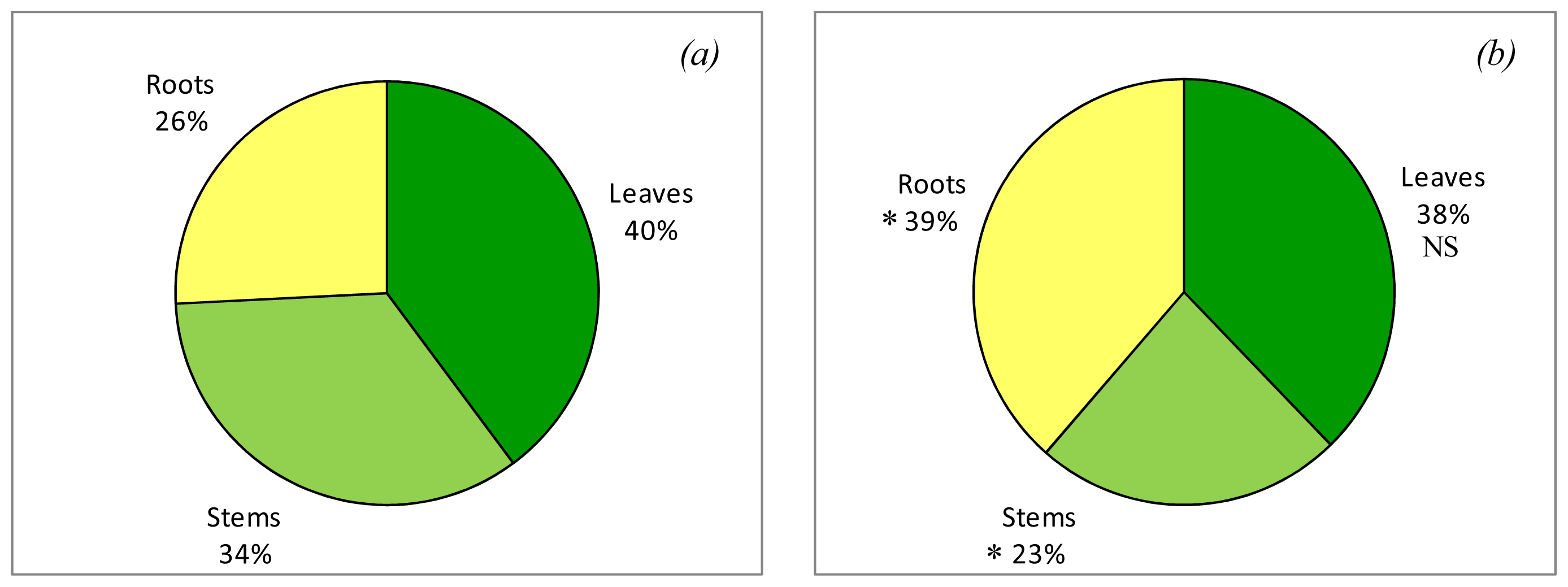
2.2. Effects on Plant Composition
3. Discussion
3.1. Effect of Nitrogen Deprivation on Plant Development
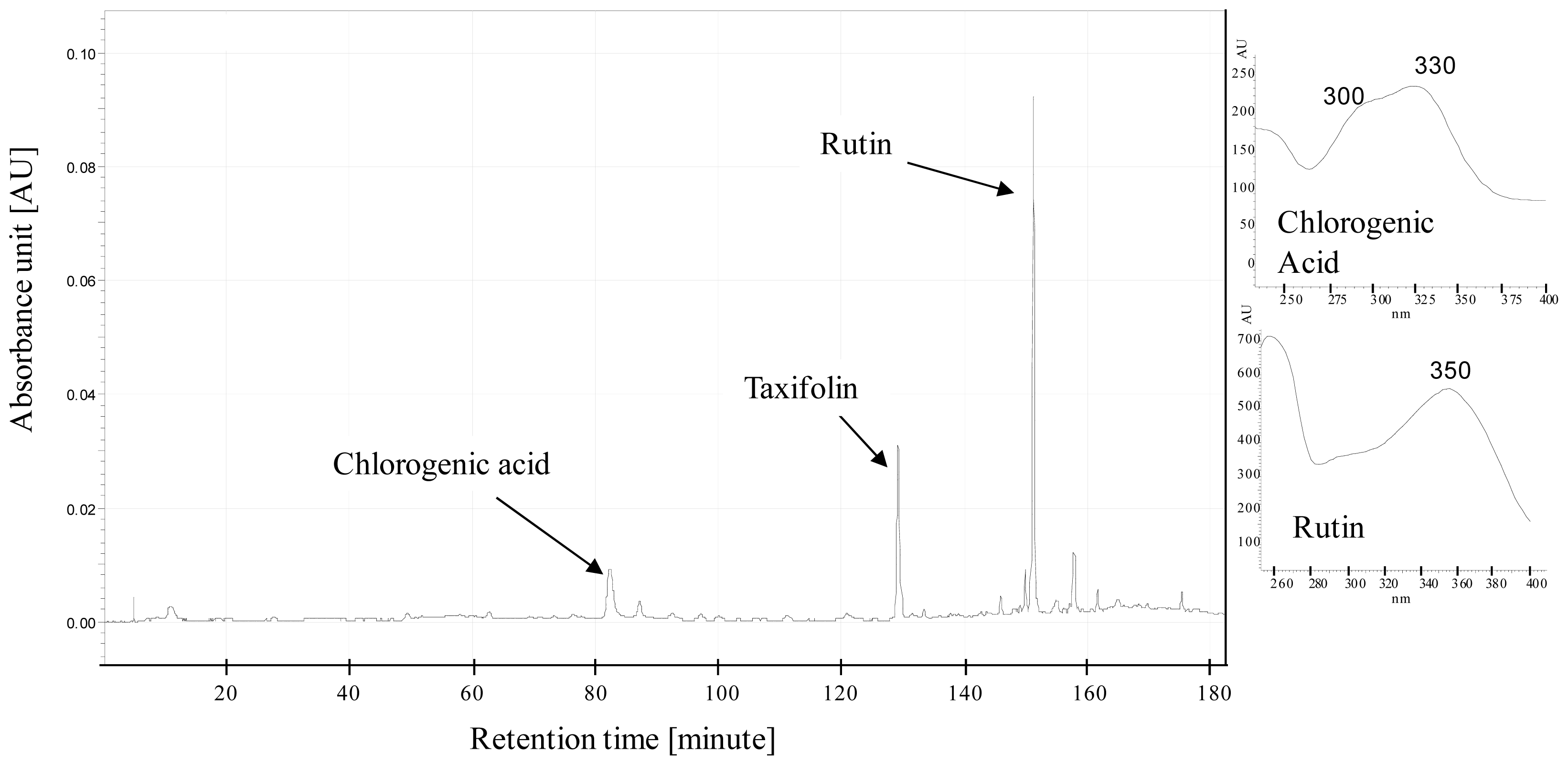
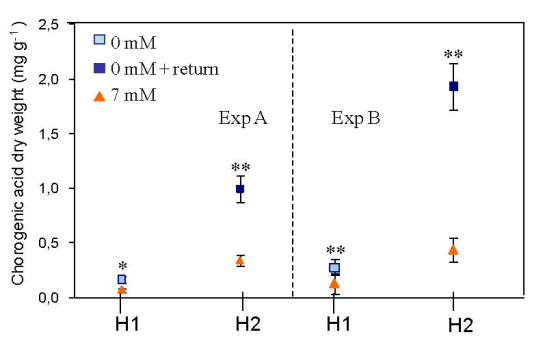
3.2. Effect of Nitrogen Deprivation on Leaf Composition
4. Experimental Section
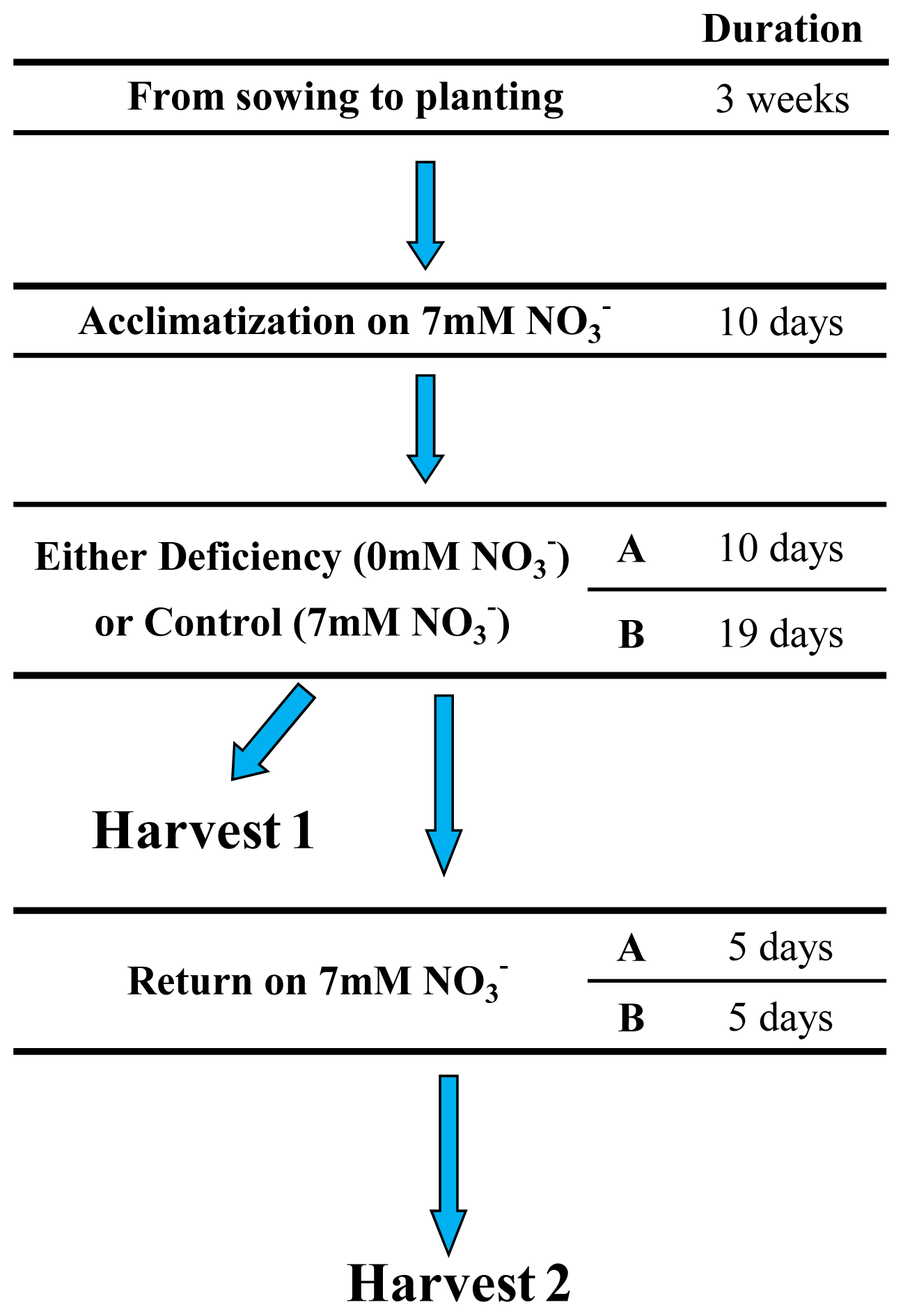
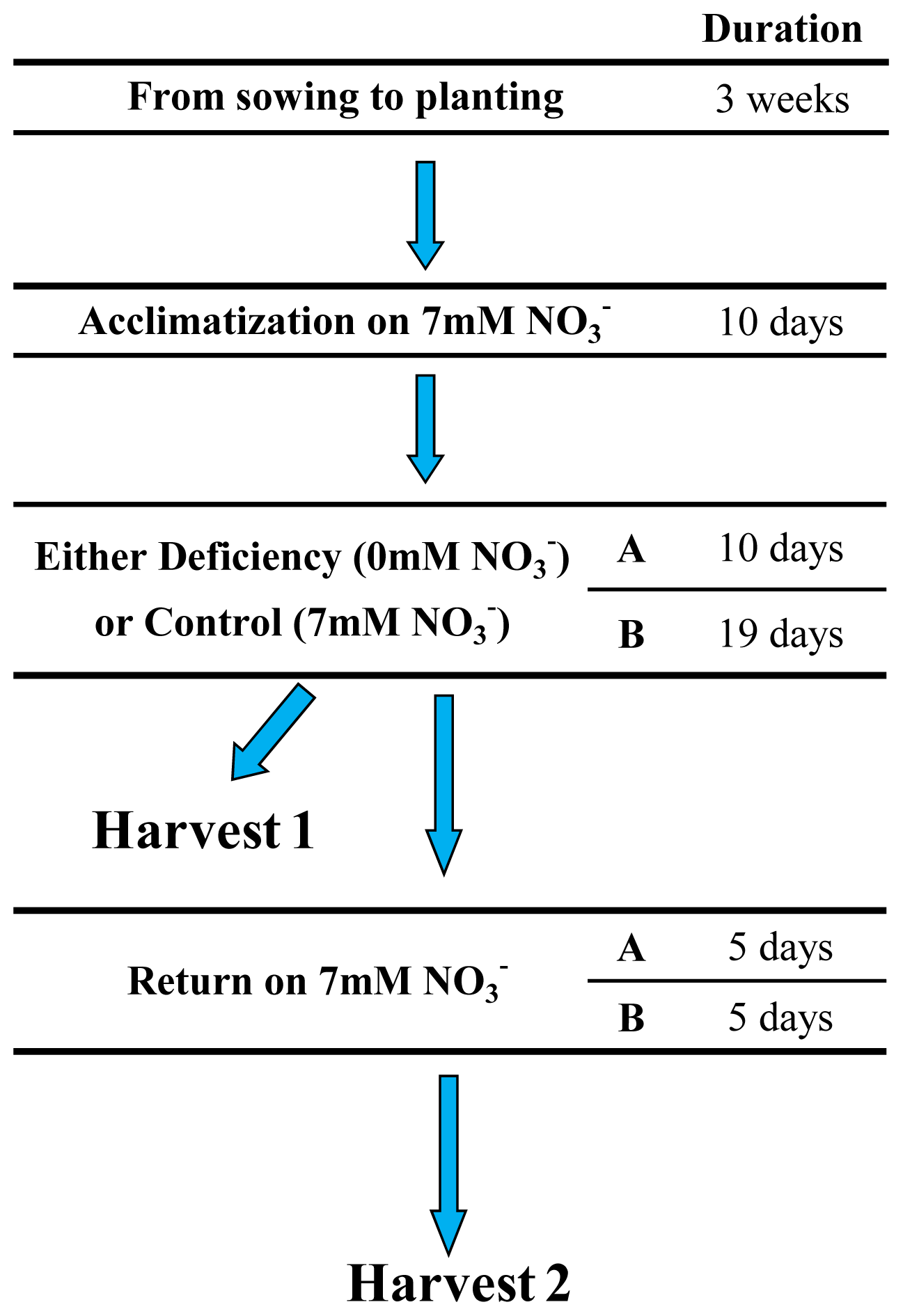
4.1. Plant Materials, Growth Conditions and Nitrogen Treatments
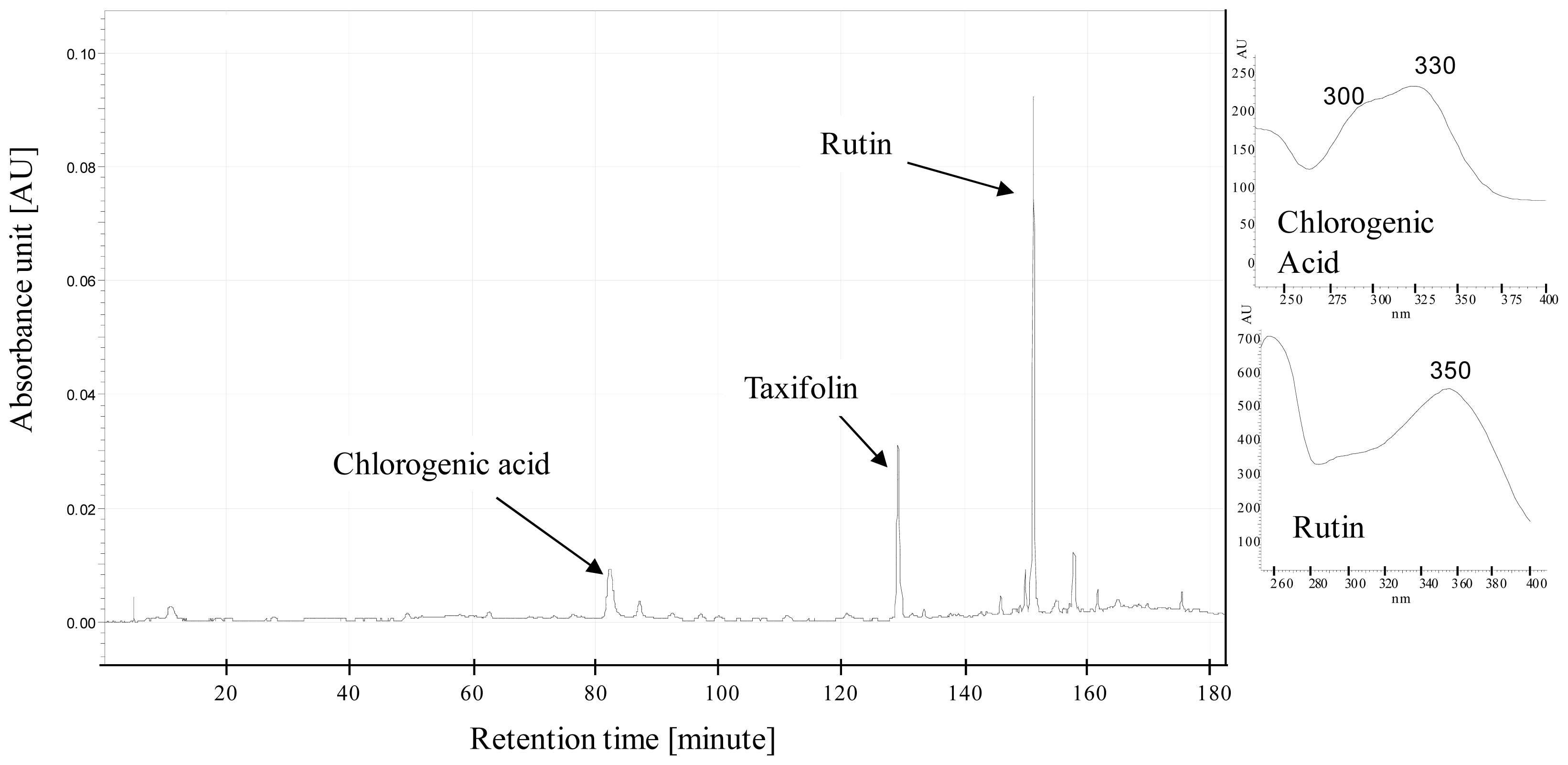
4.2. Plant Harvesting, Sampling and Chemical Analyses
5. Conclusions
References
- Wilkens, R.T.; Spoerke, J.M.; Stamp, N.E. Differential responses of growth and two soluble phenolics of tomato to resource availability. Ecology 1996, 77, 247–258. [Google Scholar]
- Mittelstraβ, K.; Treutter, D.; Pleβl, M.; Heller, W.; Elstner, E.F.; Heiser, L. Modification of primary and secondary metabolism of potato plants by nitrogen application differentially affects resistance to Phytophtora infestans and Alternaria solani. Plant Biol 2006, 8, 653–661. [Google Scholar]
- Niggeweg, R.; Michael, A.; Martin, C. Engineering plants with increased levels of the antioxidant chlorogenic acid. Nat. Biotechnol 2004, 22, 746–754. [Google Scholar]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett 2006, 4, 147–157. [Google Scholar]
- Hoffland, E.; Dicke, M.; Van Tintelen, W.; Dijkman, H.; Van Beusichem, M.L. Nitrogen availability and defense of tomato against two-spotted spider mite. J. Chem. Ecol 2000, 26, 2697–2711. [Google Scholar]
- Johnson, K.S. Plant phenolics as radical scavengers in the context of insect (Manduca sexta) hemolymph and midgut fluid. J. Agric. Food Chem 2005, 53, 10120–10126. [Google Scholar]
- Johnson, K.S.; Felton, G.W. Plant phenolics as dietary antioxidants for herbivorous insects: A test with genetically modified tobacco. J. Chem. Ecol 2001, 27, 2579–2597. [Google Scholar]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol 2006, 9, 436–442. [Google Scholar]
- Lillo, C.; Lea, U.S.; Ruoff, P. Nutrient depletion as a key factor for manipulating gene expression and product formation in different branches of the flavonoid pathway. Plant Cell Environ 2008, 31, 587–601. [Google Scholar]
- Urbanczyk-Wochniak, E.; Fernie, A.R. Metabolic profiling reveals altered nitrogen nutrient regimes have diverse effects on the metabolism of hydroponically-grown tomato (Solanum lycopersicum) plants. J. Exp. Bot 2005, 56, 309–321. [Google Scholar]
- Dumas, Y.; Dadomo, M.; Di Lucca, G.; Grolier, P. Review of the influence of major environmental and agronomic factors on the lycopene content of tomato fruit. Acta Hortic 2002, 579, 595–601. [Google Scholar]
- Le Bot, J.; Benard, C.; Robin, C.; Bourgaud, F.; Adamowicz, S. The “trade-off” between synthesis of primary and secondary compounds in young tomato leaves is altered by nitrate nutrition: experimental evidence and model consistency. J. Exp. Bot 2009, 60, 4301–4314. [Google Scholar]
- Stout, M.J.; Brovont, R.A.; Duffey, S.S. Effect of nitrogen availability on expression of constitutive and inducible chemical defenses in tomato, Lycopersicon esculentum. J. Chem. Ecol 1998, 24, 945–963. [Google Scholar]
- Lovdal, T.; Olsen, K.M.; Slimestad, R.; Verheul, M.; Lillo, C. Synergetic effects of nitrogen depletion, temperature, and light on the content of phenolic compounds and gene expression in leaves of tomato. Phytochemistry 2010, 71, 605–613. [Google Scholar]
- Le Bot, J.; Jeannequin, B.; Fabre, R. Impact of N-deprivation on the yield and nitrogen budget of rockwool grown tomatoes. Agronomie 2001, 21, 341–350. [Google Scholar]
- Toth, V.R.; Meszaros, I.; Palmer, S.J.; Veres, S.; Precsenyi, I. Nitrogen deprivation induces changes in the leaf elongation zone of maize seedlings. Biol. Plant 2002, 45, 241–247. [Google Scholar]
- Warner, J.; Zhang, T.; Hao, X. Effects of nitrogen fertilization on fruit yields and quality of processing tomatoes. Can. J. Plant Sci 2004, 84, 865–871. [Google Scholar]
- Murshed, R.; Lopez-Lauri, F.; Keller, C.; Monnet, F.; Sallanon, H. Acclimation to drought stress enhances oxidative stress tolerance in Solanum Lycopersicum L. fruits. Plant Stress 2008, 2, 145–151. [Google Scholar]
- Walter, J.; Nagy, L.; Heinb, R.; Rascherc, U.; Beierkuhnleinb, C.; Willnerd, E.; Jentsche, A. Do plants remember drought? Hints towards a drought-memory in grasses. Environ. Exp. Bot 2011, 71, 34–40. [Google Scholar]
- Scheible, W.R.; Morcuende, R.; Czechowski, T.; Fritz, C.; Osuna, D.; Palacios-Rojas, N.; Schindelasch, D.; Thimm, O.; Udvardi, M.K.; Stitt, M. Genome-wide reprogramming of primary and secondary metabolism, protein synthesis, cellular growth processes, and the regulatory infrastructure of Arabidopsis in response to nitrogen. Plant Physiol 2004, 136, 2483–2499. [Google Scholar]
- Brouwer, R. Distribution of dry matter in the plant. Neth. J. Agric. Sci 1962, 10, 361–376. [Google Scholar]
- Lemaire, F. Comparative action of nitrogen supply on growth of root system and aerial parts of plants. Ann. Agron 1975, 26, 59–73. [Google Scholar]
- Cartelat, A.; Cerovic, Z.G.; Goulas, Y.; Meyer, S.; Lelarge, C.; Prioul, J.L.; Barbottin, A.; Jeuffroy, M.H.; Gate, P.; Agati, G.; et al. Optically assessed contents of leaf polyphenolics and chlorophyll as indicators of nitrogen deficiency in wheat (Triticum aestivum L.). Field Crops Res 2005, 91, 35–49. [Google Scholar]
- Kovacik, J.; Klejdus, B.; Backor, M.; Repcak, M. Phenylalanine ammonia-lyase activity and phenolic compounds accumulation in nitrogen-deficient Matricaria chamomilla leaf rosettes. Plant Sci 2007, 172, 393–399. [Google Scholar]
- Herms, D.A.; Mattson, W.J. The dilemma of plants: to grow or defend. Quart. Rev. Biol 1992, 67, 283–335. [Google Scholar]
- Baidez, A.G.; Gomez, P.; Del Rio, J.A.; Ortuno, A. Antifungal capacity of major phenolic compounds of Olea europaea L. against Phytophthora megasperma Drechsler and Cylindrocarpon destructans (Zinssm.) Scholten. Physiol. Mol. Plant Pathol 2006, 69, 224–229. [Google Scholar]
- Lattanzio, V.; Lattanzio, V.M.T.; Cardinali, A. Role of Phenolics in the Resistance Mechanisms of Plants against Fungal Pathogens and Insects. In Phytochemistry: Advances in Research; Research Signpost: Kerala, India, 2006; pp. 23–67. [Google Scholar]
- Benard, C.; Gautier, H.; Bourgaud, F.; Grasselly, D.; Navez, B.; Caris-Veyrat, C.; Weiss, M.; Genard, M. Effects of low nitrogen supply on tomato (Solanum lycopersicum) fruit yield and quality with special emphasis on sugars, acids, ascorbate, carotenoids, and phenolic compounds. J. Agric. Food Chem 2009, 57, 4112–4123. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound/Weight (g) | KH2PO4 | K2SO4 | Ca(NO3)2, 4H2O | CaSO4, 2H2O | MgSO4, 7H2O | EDTA-Fe, 1H2O | Oligoelements (mL/L) | |
|---|---|---|---|---|---|---|---|---|
| NO3− (mM) | 0 | 0.136 | 0.174 | 0 | 0.603 | 0.37 | 0.017 | 0.1 |
| 7 | 0.827 | 0 |
| Compound | MoO3 | MnCl2, 4H2O | ZnSO4, 7H2O | CuSO4, 5H2O | H3BO3 | FeSO4 | Na2EDTA |
|---|---|---|---|---|---|---|---|
| Weight (g) | 0.40 | 23.06 | 9.32 | 1.18 | 12.75 | 29.87 | 59.56 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bénard, C.; Bourgaud, F.; Gautier, H. Impact of Temporary Nitrogen Deprivation on Tomato Leaf Phenolics. Int. J. Mol. Sci. 2011, 12, 7971-7981. https://doi.org/10.3390/ijms12117971
Bénard C, Bourgaud F, Gautier H. Impact of Temporary Nitrogen Deprivation on Tomato Leaf Phenolics. International Journal of Molecular Sciences. 2011; 12(11):7971-7981. https://doi.org/10.3390/ijms12117971
Chicago/Turabian StyleBénard, Camille, Frédéric Bourgaud, and Hélène Gautier. 2011. "Impact of Temporary Nitrogen Deprivation on Tomato Leaf Phenolics" International Journal of Molecular Sciences 12, no. 11: 7971-7981. https://doi.org/10.3390/ijms12117971