Screening and Evaluation of Polyhydroxybutyrate-Producing Strains from Indigenous Isolate Cupriavidus taiwanensis Strains


Abstract
:1. Introduction
2. Results and Discussions
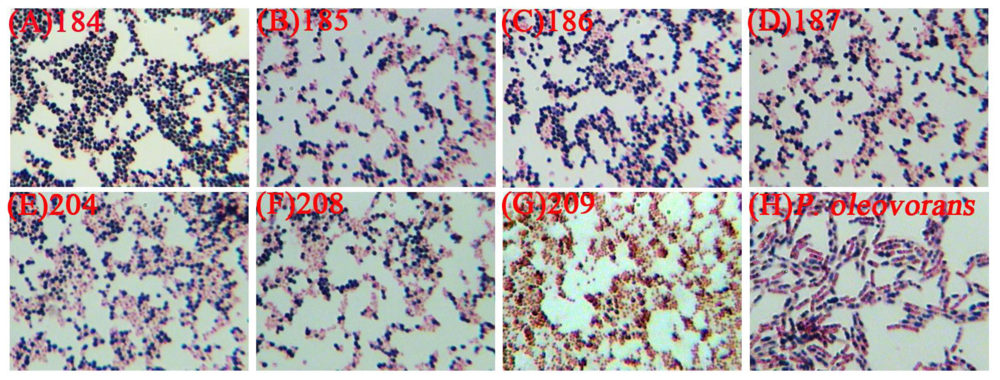
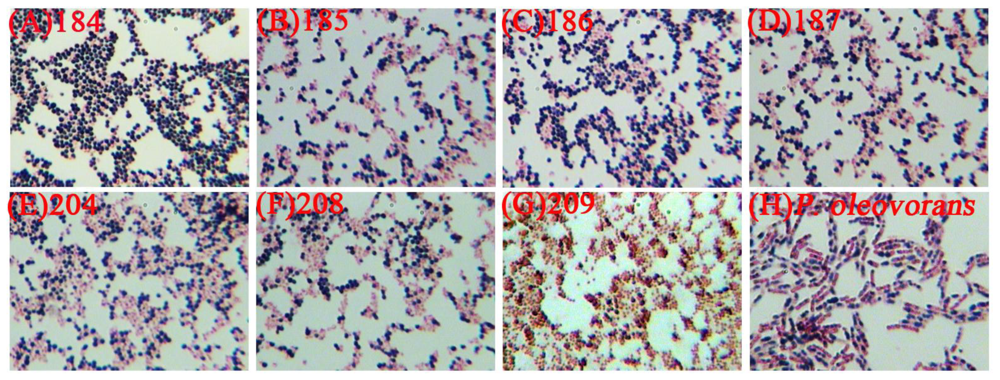
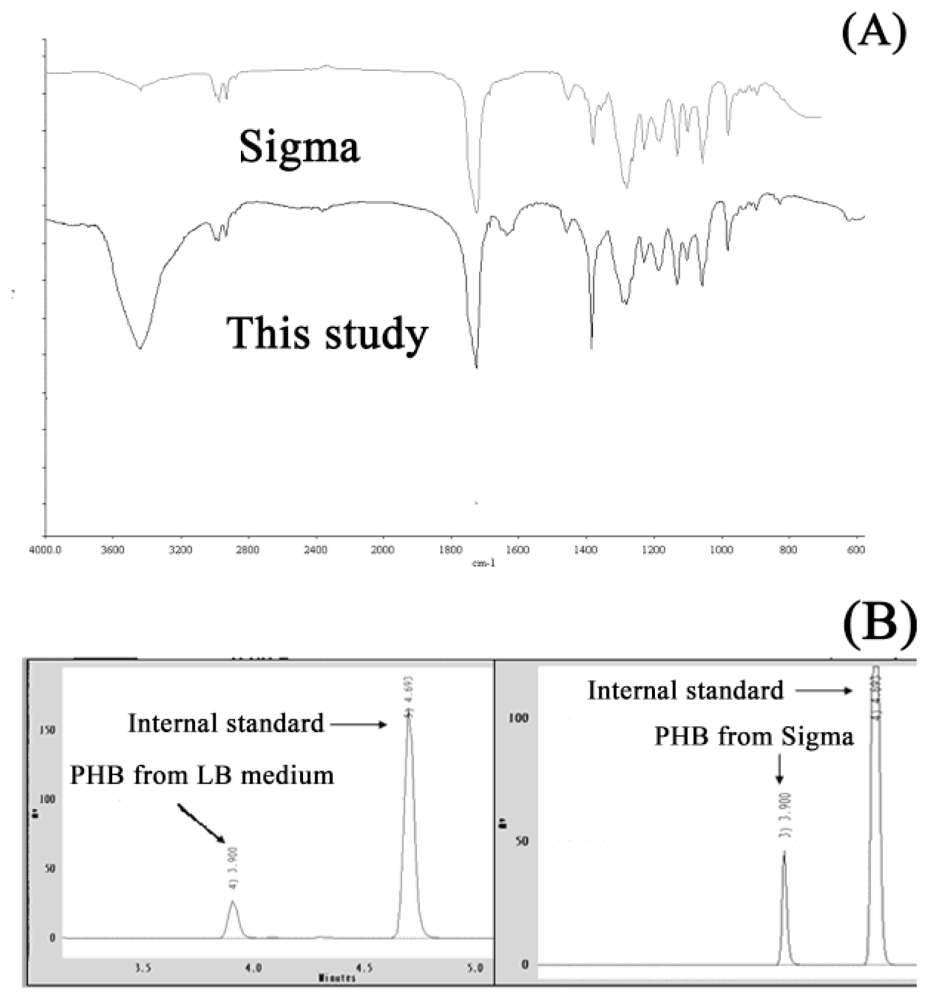
2.1. The Selection of PHB Production Strain
2.2. Effect of Growth Condition on PHB Production by C. taiwanensis 184
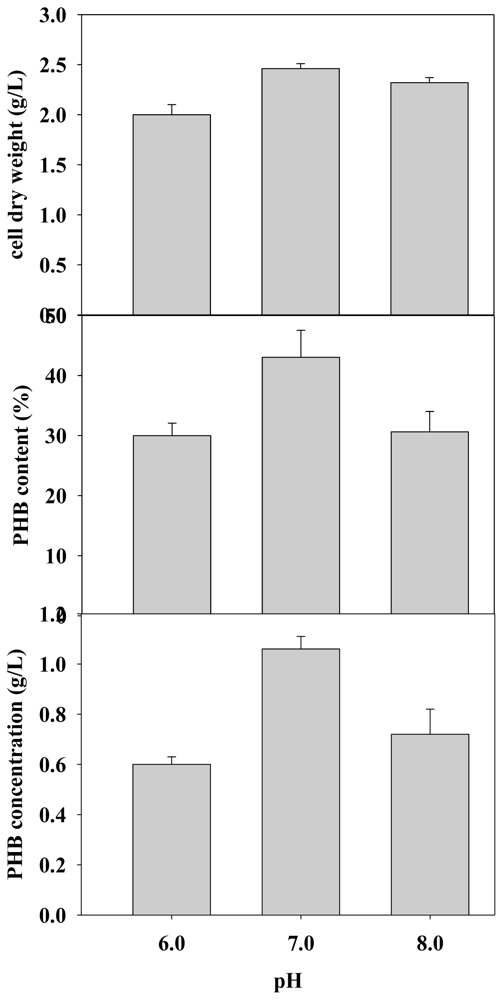
2.2.1. Effect of Initial Culture pH
2.2.2. Effect of Culture Temperature
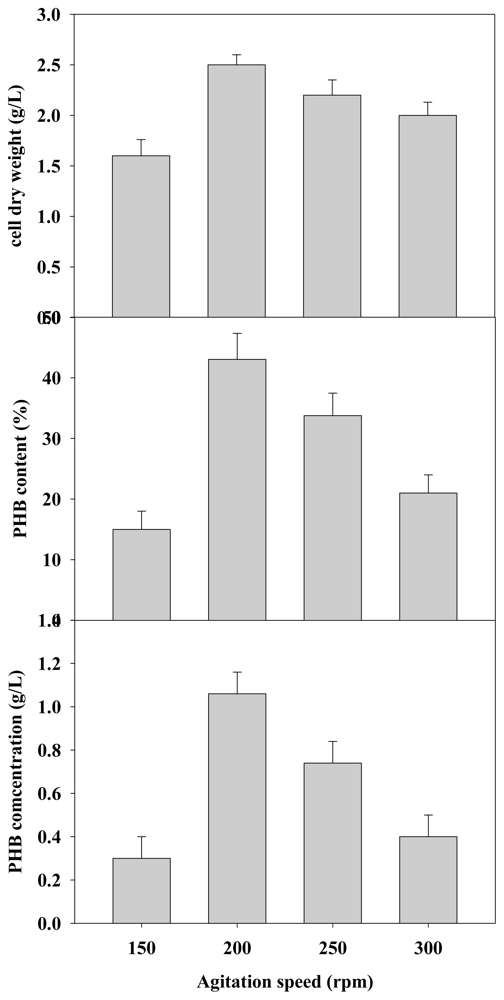
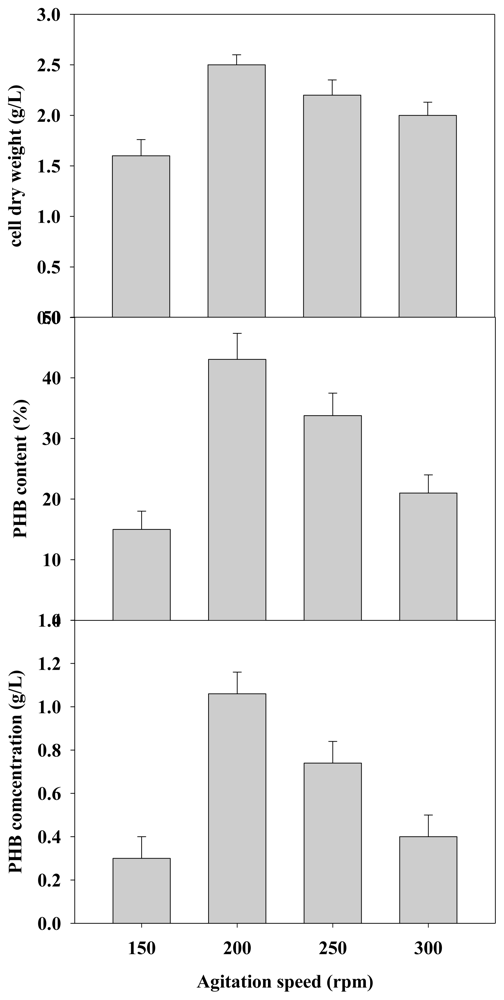
2.2.3. Effect of Agitation Rate
2.3. Media Optimization
2.3.1. Effect of Carbon and Nitrogen Types on PHB Production by C. taiwanensis 184
2.3.2. Effect of Carbon/Nitrogen Ratio on PHB Production by C. taiwanensis 184
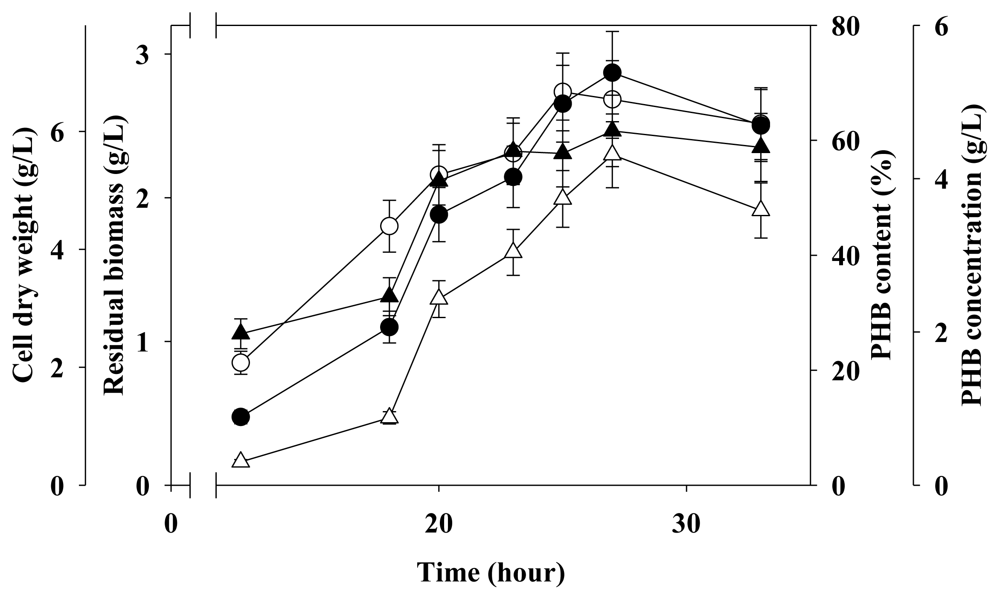
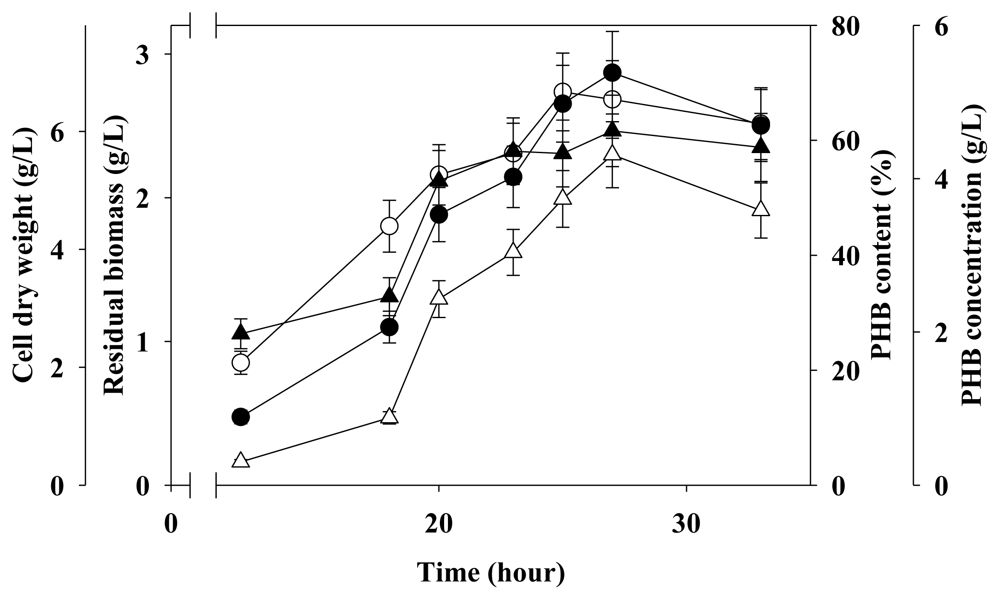
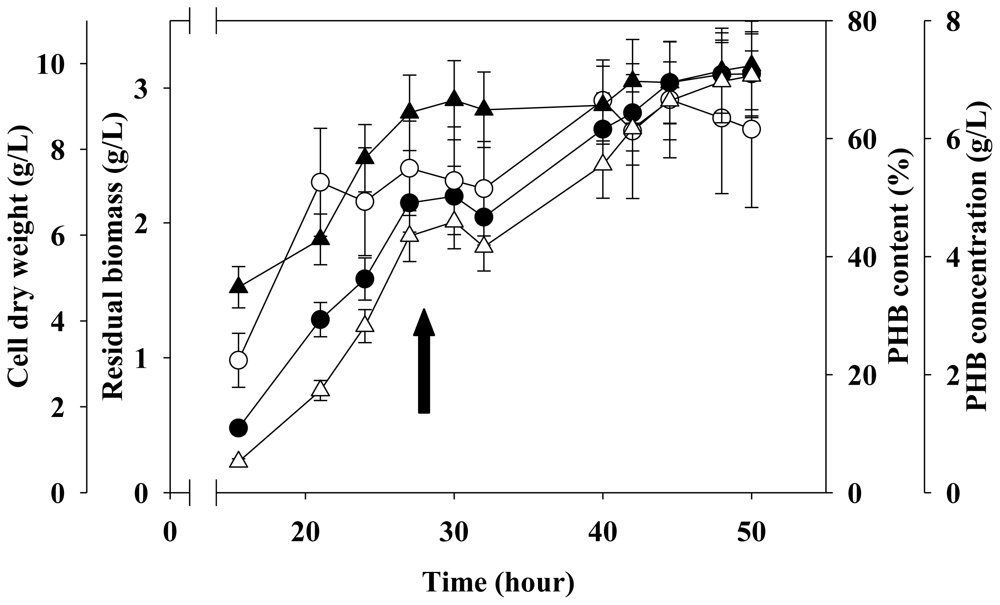
2.4. Two-Stage Fermentation in 5 L Fermenter
3. Experimental Section
3.1. Microorganisms Screening for PHB Production
3.2. PHB Production Media and Conditions
3.3. Quantifications of Cell Growth and PHB
3.4. Two-Stage Fermentation Strategy for PHB Production
3.5. Staining of Bacteria with Sudan Black B
3.6. Analytical Methods
4. Conclusion
Acknowledgements
References
- Chakraborty, P; Gibbons, W; Muthukumarappanl, K. Conversion of volatile fatty acids into polyhydroxyalkanoate by Ralstonia eutropha. J. Appl. Microbiol 2009, 106, 1996–2005. [Google Scholar]
- Yu, PH; Chua, H; Huang, AL; Ho, KP. Conversion of industrial food wastes by Alcaligenes latus into polyhydroxyalkanoates. Appl Biochem Biotechnol 1999, 77–79, 445–454. [Google Scholar]
- Zhang, HF; Ma, L; Wang, ZH; Chen, GQ. Biosynthesis and characterization of 3-hydroxyalkanoate terpolyesters with adjustable properties by Aeromonas hydrophila. Biotechnol. Bioeng 2009, 104, 582–589. [Google Scholar]
- Ren, Q; de Roo, G; Witholt, B; Zinn, M; Thöny-Meyer, L. Influence of growth stage on activities of polyhydroxyalkanoate (PHA) polymerase and PHA depolymerase in Pseudomonas putida U. BMC Microbiol 2010, 10, 254. [Google Scholar]
- Singh, M; Patel, SK; Kalia, VC. Bacillus subtilis as potential producer for polyhydroxyalkanoates. Microb. Cell Fact 2009, 8, 38. [Google Scholar]
- Chanprateep, S. Current trends in biodegradable polyhydroxyalkanoates. J. Biosci. Bioeng 2010, 110, 621–632. [Google Scholar]
- Chen, GQ. A microbial polyhydroxyalkanoates (PHA) based bio- and materials industry. Chem. Soc. Rev 2009, 38, 2434–2446. [Google Scholar]
- Dong, Y; Li, P; Chen, CB; Wang, ZH; Ma, P; Chen, GQ. The improvement of fibroblast growth on hydrophobic biopolyesters by coating with polyhydroxyalkanoate granule binding protein PhaP fused with cell adhesion motif RGD. Biomaterials 2010, 31, 8921–8930. [Google Scholar]
- Cousley, RR. A stent-guided mini-implant system. J. Clin. Orthod 2009, 43, 403–407. [Google Scholar]
- Xu, XY; Li, XT; Peng, SW; Xiao, JF; Liu, C; Fang, G; Chen, KC; Chen, GQ. The behaviour of neural stem cells on polyhydroxyalkanoate nanofiber scaffolds. Biomaterials 2010, 31, 3967–3975. [Google Scholar]
- Han, I; Shim, KJ; Kim, JY; Im, SU; Sung, YK; Kim, M; Kang, IK; Kim, JC. Effect of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) nanofiber matrices cocultured with hair follicular epithelial and dermal cells for biological wound dressing. Artif Organs 2007, 31, 801–808. [Google Scholar]
- Yilgor, P; Hasirci, N; Hasirci, V. Sequential BMP-2/BMP-7 delivery from polyester nanocapsules. J. Biomed. Mater. Res A 2010, 93, 528–536. [Google Scholar]
- Zhu, XH; Wang, CH; Tong, YW. In vitro characterization of hepatocyte growth factor release from PHBV/PLGA microsphere scaffold. J. Biomed. Mater. Res A 2009, 89, 411–423. [Google Scholar]
- Xiong, YC; Yao, YC; Zhan, XY; Chen, GQ. Application of polyhydroxyalkanoates nanoparticles as intracellular sustained drug-release vectors. J. Biomater. Sci. Polym. Ed 2010, 21, 127–140. [Google Scholar]
- Burdon, KL. Fatty materials in bacteria and fungi revealed by staining dried, fixed slide preparations. J. Bacteriol 1946, 52, 665–678. [Google Scholar]
- Hartmann, R; Hany, R; Witholt, B; Zinn, M. Simultaneous biosynthesis of two copolymers in Pseudomonas putida GPo1 using a two-stage continuous culture system. Biomacromolecules 2010, 11, 1488–1493. [Google Scholar]
- Palleroni, NJ; Palleroni, AV. Alcaligenes latus, a new species of hydrogen-utilizing bacteria. Int. J. Syst. Bacteriol 1978, 28, 416–424. [Google Scholar]
- Chanprateep, S; Katakura, Y; Visetkoop, S; Shimizu, H; Kulpreecha, S; Shioya, S. Characterization of new isolated Ralstonia eutropha strain A-04 and kinetic study of biodegradable copolyester poly(3-hydroxybutyrate-co-4-hydroxybutyrate) production. J. Ind. Microbiol. Biotechnol 2008, 35, 1205–1215. [Google Scholar]
- Sun, Z; Ramsay, JA; Guay, M; Ramsay, B. Increasing the yield of MCL-PHA from nonanoic acid by co-feeding glucose during the PHA accumulation stage in two-stage fed-batch fermentations of Pseudomonas putida KT2440. J. Biotechnol 2007, 132, 280–282. [Google Scholar]
- Albuquerque, MG; Eiroa, M; Torres, C; Nunes, BR; Reis, MA. Strategies for the development of a side stream process for polyhydroxyalkanoate (PHA) production from sugar cane molasses. J. Biotechnol 2007, 130, 411–421. [Google Scholar]
- Liu, HY; Hall, PV; Darby, JL; Coats, ER; Green, PG; Thompson, DE; Loge, FJ. Production of polyhydroxyalkanoate during treatment of tomato cannery wastewater. Water Environ. Res 2008, 80, 367–372. [Google Scholar]
- Lee, SY; Wong, HH; Choi, J; Lee, SH; Lee, SC; Han, CS. Production of medium-chainlength polyhydroxyalkanoates by high-cell-density cultivation of Pseudomonas putida under phosphorus limitation. Biotechnol. Bioeng 2000, 68, 466–470. [Google Scholar]
- Valappil, SP; Rai, R; Bucke, C; Roy, I. Polyhydroxyalkanoate biosynthesis in Bacillus cereus SPV under varied limiting conditions and an insight into the biosynthetic genes involved. J. Appl. Microbiol 2008, 104, 1624–1635. [Google Scholar]
- Wang, F; Lee, SY. Poly(3-hydroxybutyrate) production with high productivity and high polymer content by a Fed-batch culture of Alcaligenes latus under nitrogen limitation. Appl Environ Microbiol 1997, 63, 3703–3706. [Google Scholar]
- Albuquerque, MG; Torres, CA; Reis, MA. Polyhydroxyalkanoate (PHA) production by a mixed microbial culture using sugar molasses: effect of the influent substrate concentration on culture selection. Water Res 2010 44, 3419–3433.
- Shang, L; Jiang, M; Chang, HN. Poly(3-hydroxybutyrate) synthesis in fed-batch culture of Ralstonia eutropha with phosphate limitation under different glucose concentrations. Biotechnol. Lett 2003, 25, 1415–1419. [Google Scholar]
- Lütke-Eversloh, T; Bergander, K; Luftmann, H; Steinbüchel, A. Identification of a new class of biopolymer: bacterial synthesis of a sulfur-containing polymer with thioester linkages. Microbiology 2001, 147, 11–19. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | PHB content (%) | PHB concentration (g/L) |
|---|---|---|
| C. taiwanensis 184 | 9.67 ± 0.11 | 0.15 ± 0.03 |
| C. taiwanensis 185 | 3.43 ± 0.09 | 0.07 ± 0.01 |
| C. taiwanensis 186 | 2.08 ± 0.02 | 0.09 ± 0.02 |
| C. taiwanensis 187 | 0.45 ± 0.06 | 0.03 ± 0.01 |
| C. taiwanensis 204 | 0.54 ± 0.08 | 0.04 ± 0.01 |
| C. taiwanensis 208 | 0.25 ± 0.03 | 0.01 ± 0.01 |
| C. taiwanensis 209 | 0.63 ± 0.06 | 0.02 ± 0.01 |
| P. oleovorans ATCC 29347 | 0.09 ± 0.01 | 0.03 ± 0.01 |
| Carbon types | Nitrogen types | ||||
|---|---|---|---|---|---|
| Carbohydrates | Hydrocarbons | Inorganic nitrogen | |||
| Carbon source | PHB production | Carbon source | PHB production (g/L) | Nitrogen source | PHB production (g/L) |
| Glucose | + | Ethanol | − | NH4Cl | ++ |
| Glucose (food) | + | Gluconic acid | ++ | CH3COONH2 | + |
| Lactose | − | Oxalic acid | − | NH4NO3 | + |
| Raffinose | − | Malic acid | − | (NH4)2SO4 | + |
| Sucrose | − | Galacturoni c acid | − | NH2CONH2 | + |
| Galactose | − | ||||
| Dulcitol | − | ||||
| Mannitol | − | ||||
| C/N (mol/mol) | CDW (g/L) | PHB Concentration (g/L) | PHB Content (%) |
|---|---|---|---|
| 1/1 | 1.42 ± 0.28 | 0.42 ± 0.05 | 29.21 ± 2.33 |
| 2/1 | 2.42 ± 0.62 | 1.04 ± 0.12 | 43.10 ± 3.15 |
| 4/1 | 2.45 ± 0.57 | 1.20 ± 0.14 | 48.86 ± 2.19 |
| 8/1 | 4.15 ± 0.91 | 2.44 ± 0.36 | 58.81 ± 3.93 |
| 20/1 | 1.69 ± 0.43 | 0.89 ± 0.02 | 52.77 ± 2.62 |
© 2011 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wei, Y.-H.; Chen, W.-C.; Huang, C.-K.; Wu, H.-S.; Sun, Y.-M.; Lo, C.-W.; Janarthanan, O.-M. Screening and Evaluation of Polyhydroxybutyrate-Producing Strains from Indigenous Isolate Cupriavidus taiwanensis Strains. Int. J. Mol. Sci. 2011, 12, 252-265. https://doi.org/10.3390/ijms12010252
Wei Y-H, Chen W-C, Huang C-K, Wu H-S, Sun Y-M, Lo C-W, Janarthanan O-M. Screening and Evaluation of Polyhydroxybutyrate-Producing Strains from Indigenous Isolate Cupriavidus taiwanensis Strains. International Journal of Molecular Sciences. 2011; 12(1):252-265. https://doi.org/10.3390/ijms12010252
Chicago/Turabian StyleWei, Yu-Hong, Wei-Chuan Chen, Chin-Kuei Huang, Ho-Shing Wu, Yi-Ming Sun, Chi-Wei Lo, and Om-Murugan Janarthanan. 2011. "Screening and Evaluation of Polyhydroxybutyrate-Producing Strains from Indigenous Isolate Cupriavidus taiwanensis Strains" International Journal of Molecular Sciences 12, no. 1: 252-265. https://doi.org/10.3390/ijms12010252