Antioxidant Protective Effect of Glibenclamide and Metformin in Combination with Honey in Pancreas of Streptozotocin-Induced Diabetic Rats

Abstract
:1. Introduction
2. Methods
2.1. Processing and Preparation of Tissue
2.2. Superoxide Dismutase (SOD) Assay
2.3. Glutathione Peroxidase (GPx) Assay
2.4. Catalase (CAT) Assay
2.5. Glutathione Reductase (GR) Assay
2.6. Glutathione-S-Transferase (GST) Assay
2.7. Lipid Peroxidation (LPO) Assay
2.8. Protein Assay
2.9. Statistical Analysis
3. Results
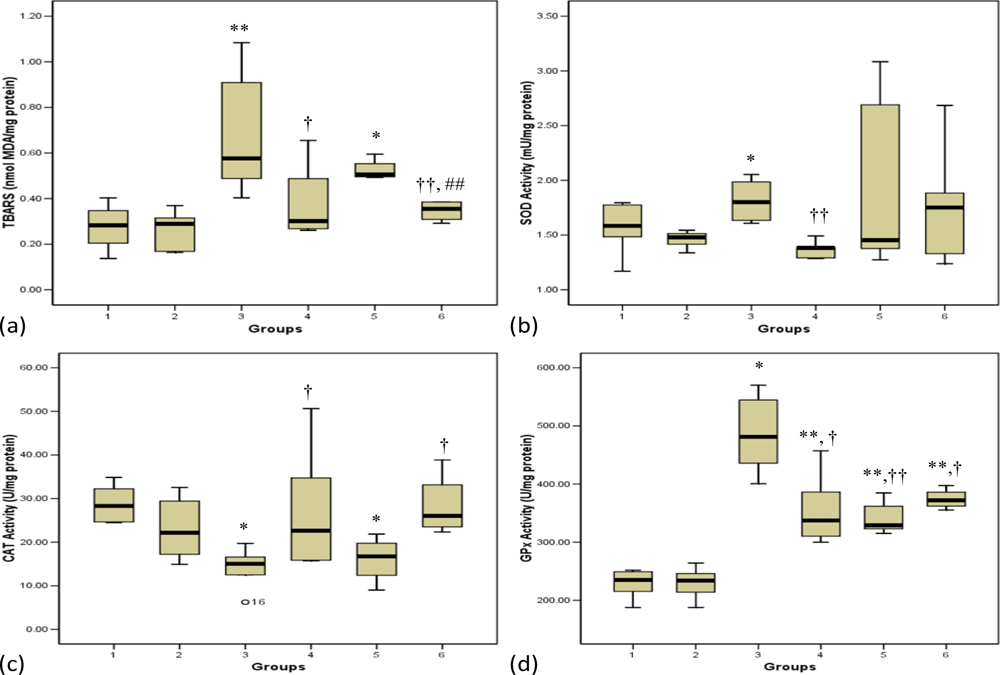
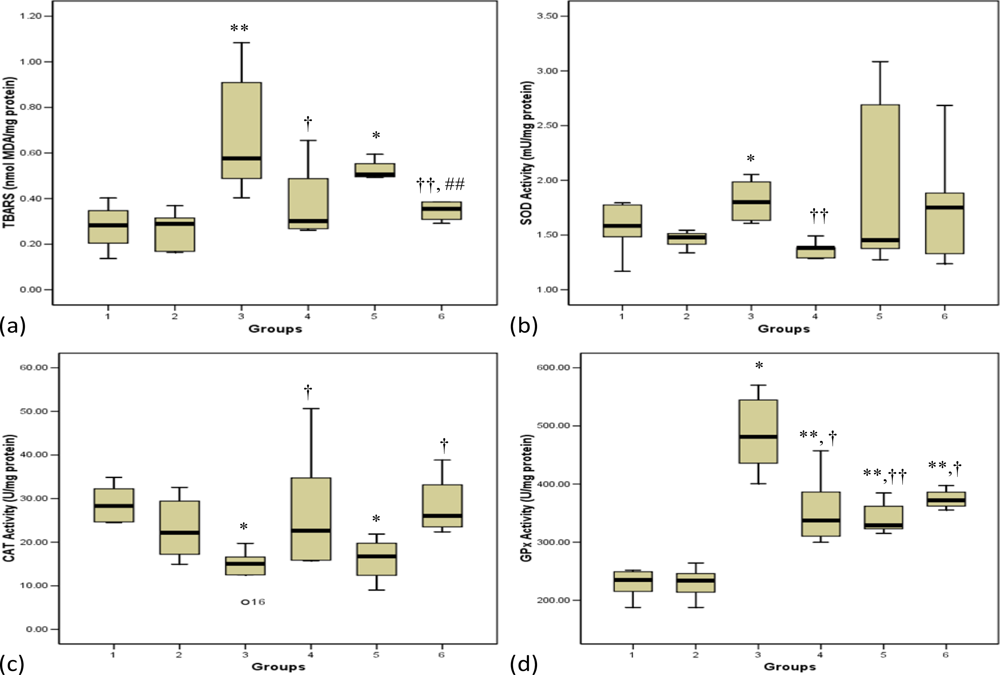
3.1. Lipid Peroxidation
3.2. Antioxidant Enzymes
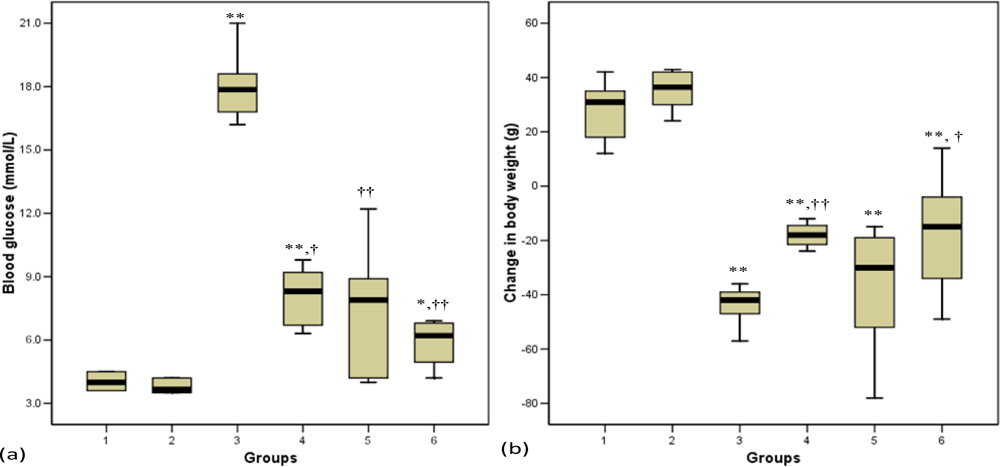
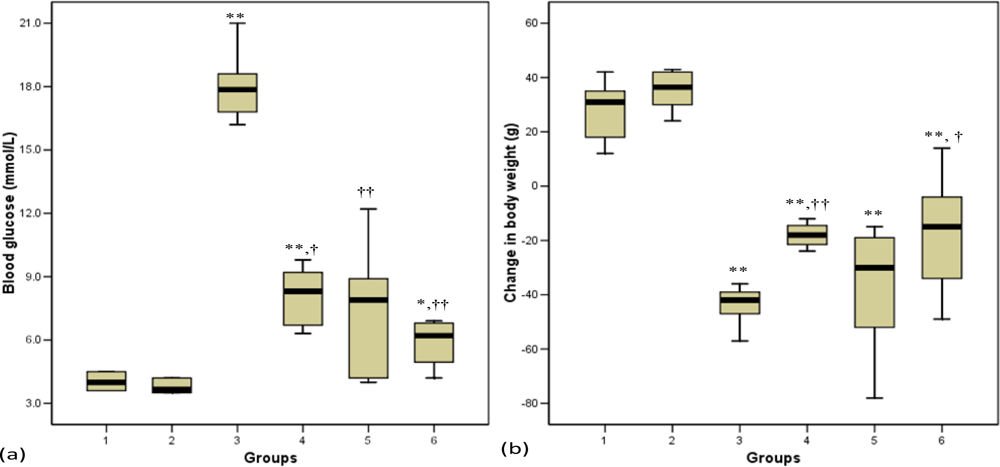
3.3. Blood Glucose
3.4. Body Weight
4. Discussion
5. Conclusions
Acknowledgments
References and notes
- Soret, MG; Dulin, WE; Mathews, J; Gerritsen, GC. Morphologic abnormalities observed in retina, pancreas, and kidney of diabetic Chinese hamsters. Diabetologia 1974, 10, 567–579. [Google Scholar]
- Cristina, L; Roberto, L; Stefano, DP. β-cell failure in type 2 diabetes mellitus. Curr. Diab. Rep 2008, 8, 179–184. [Google Scholar]
- Cook, MN; Girman, CJ; Stein, PP; Alexander, CM; Holman, RR. Glycemic control continues to deteriorate after sulfonylureas are added to metformin among patients with type 2 diabetes. Diabetes Care 2005, 28, 995–1000. [Google Scholar]
- Giugliano, D; Ceriello, A; Paolisso, G. Oxidative stress and diabetic vascular complications. Diabetes Care 1996, 19, 257–267. [Google Scholar]
- Baynes, JW; Thorpe, SR. The role of oxidative stress in diabetic complications. Curr. Opin. Endocrinol 1999, 3, 277–284. [Google Scholar]
- Bonnefont-Rousselot, D; Bastard, JP; Jaudon, MC; Delattre, J. Consequences of the diabetic status on the oxidant / antioxidant balance. Diab. Metab 2000, 26, 163–176. [Google Scholar]
- Telci, A; Cakatay, U; Salman, S; Satman, I; Sivas, A. Oxidative protein damage in early stage in type 1 diabetic patients. Diabetes Res. Clin. Pract 2000, 50, 213–223. [Google Scholar]
- Turk, HM; Sevinc, A; Camci, C; Cigli, A; Buyukberber, S; Savli, H; Bayraktar, N. Plasma lipid peroxidation products and antioxidant enzyme activities in patients with type 2 diabetes mellitus. Acta Diabetol 2002, 39, 117–122. [Google Scholar]
- Cnop, M; Welsh, N; Jonas, JC; Jörns, A; Lenzen, S; Eizirik, DL. Mechanisms of pancreatic beta-cell death in type 1 and type 2 diabetes: many differences, few similarities. Diabetes 2005, 54, S97–S107. [Google Scholar]
- Evans, JL; Goldfine, ID; Maddux, BA; Grodsky, GM. Are oxidative stress-activated signaling pathways mediators of insulin resistance and β-cell dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar]
- Simmons, RA. Developmental origins of diabetes: the role of oxidative stress. Free Radic. Biol. Med 2006, 40, 917–922. [Google Scholar]
- The Diabetes Control and Complications Trial (DCCT) Research Group. The effect of intensive treatment of diabetes on the development and progression of long-term complications of insulin-dependent diabetes mellitus. N. Engl. J. Med 1993, 329, 977–980. [Google Scholar]
- UK Prospective Diabetes Study (UKPDS) Group. Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33). Lancet 1998, 352, 837–853. [Google Scholar]
- Erejuwa, OO; Sulaiman, SA; Ab Wahab, MS; Sirajudeen, KNS; Salzihan, MS. Effects of Malaysian tualang honey supplementation on glycemia, free radical scavenging enzymes and markers of oxidative stress in kidneys of normal and streptozotocin-induced diabetic rats. Int. J. Cardiol 2009, 137, S45. [Google Scholar]
- Gott, L. A simple method for determination of serum catalase activity and revision of reference range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar]
- Goldberg, DM; Spooner, RJ. Assay of glutathione reductase. In Methods of Enzymatic Analysis; Bergmeyen, HV, Ed.; Verlag Chemie: Weinheim, Germany, 1983; pp. 258–65. [Google Scholar]
- Habig, WH; Pabst, MJ; Jakoby, WB. Glutathione-S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem 1974, 249, 7130–7139. [Google Scholar]
- Ohkawa, H; Ohishi, N; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem 1979, 95, 351–358. [Google Scholar]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem 1976, 72, 248–254. [Google Scholar]
- Evans, JL; Goldfine, ID; Maddux, BA; Grodsky, GM. Are oxidative stress-activated signaling pathways mediators of insulin resistance and β-cell dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar]
- Lenzen, S. Oxidative stress: The vulnerable beta-cell. Biochem. Soc. Trans 2008, 36, 343–347. [Google Scholar]
- McCord, JM; Keele, BB; Fridovich, I. An enzymebased theory of obligate anaerobiosis: the physiological function of superoxide dismutase. Proc. Natl. Acad. Sci. USA 1971, 68, 1024–1027. [Google Scholar]
- Gardner, R; Salvador, A; Moradas-Ferreira, P. Why does SOD overexpression sometimes enhance, sometimes decrease, hydrogen peroxide production? A minimalist explanation. Free Radic. Biol. Med 2002, 32, 1351–1357. [Google Scholar]
- de Haan, JB; Cristiano, F; Iannello, R; Bladier, C; Kelner, MJ; Kola, I. Elevation in the ratio of Cu/Zn-superoxide dismutase to glutathione peroxidase activity induces features of cellular senescence and this effect is mediated by hydrogen peroxide. Hum. Mol. Genet 1996, 5, 283–292. [Google Scholar]
- Kono, Y; Fridovich, I. Superoxide radical inhibits catalase. J. Biol. Chem 1982, 257, 5751–5754. [Google Scholar]
- Christophersen, BO. Formation of monohydroxy-polyenic fatty acids from lipid peroxides by a glutathione peroxidase. Biochim. Biophys. Acta 1968, 164, 35–46. [Google Scholar]
- Robertson, RP; Harmon, JS. Pancreatic islet β-cell and oxidative stress: The importance of glutathione peroxidase. FEBS Lett 2007, 581, 3743–3748. [Google Scholar]
- Hauggard, N. Cellular mechanism of oxygen toxicity. Physiol. Rev 1968, 48, 311–373. [Google Scholar]
- Halliwell, B. Lipid peroxidation, antioxidants and cardiovascular disease: how should we more forward? Cardiovasc. Res 2000, 47, 410–448. [Google Scholar]
- Tjälve, H. Streptozotocin: distribution, metabolism and mechanisms of action. Uppsala J. Med. Sci 1983, 39, 145–147. [Google Scholar]
- Chen, V; Ianuzzo, CD. Dosage effect of streptozotocin on rat tissue enzyme activities and glycogen concentration. Can. J. Physiol. Pharmacol 1982, 60, 1251–1256. [Google Scholar]
- Rajkumar, L; Govindarajulu, P. Increased degradation of dermal collagen in diabetic rats. Indian J. Exp. Biol 1991, 29, 1081–1083. [Google Scholar]

{kind=link}
{kind=link}
| Group | Treatment |
|---|---|
| 1 | (Normal) Distilled water (0.5 mL) |
| 2 | (Normal) Tualang honey (1.0 g/kg/body weight) |
| 3 | (Diabetic) Distilled water (0.5 mL) |
| 4 | (Diabetic) Tualang honey (1.0 g/kg/body weight) |
| 5 | (Diabetic) Glibenclamide (0.6 mg/kg/body weight) + metformin (100 mg/kg/body weight) |
| 6 | (Diabetic) Glibenclamide (0.6 mg/kg/body weight) + metformin (100 mg/kg/body weight) + tualang honey (1.0 g/kg/body weight) |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.A.; Sirajudeen, K.N.S.; Salleh, M.S.M.; Gurtu, S. Antioxidant Protective Effect of Glibenclamide and Metformin in Combination with Honey in Pancreas of Streptozotocin-Induced Diabetic Rats. Int. J. Mol. Sci. 2010, 11, 2056-2066. https://doi.org/10.3390/ijms11052056
Erejuwa OO, Sulaiman SA, Wahab MSA, Sirajudeen KNS, Salleh MSM, Gurtu S. Antioxidant Protective Effect of Glibenclamide and Metformin in Combination with Honey in Pancreas of Streptozotocin-Induced Diabetic Rats. International Journal of Molecular Sciences. 2010; 11(5):2056-2066. https://doi.org/10.3390/ijms11052056
Chicago/Turabian StyleErejuwa, Omotayo Owomofoyon, Siti Amrah Sulaiman, Mohd Suhaimi Abdul Wahab, Kuttulebbai Nainamohammed Salam Sirajudeen, Md Salzihan Md Salleh, and Sunil Gurtu. 2010. "Antioxidant Protective Effect of Glibenclamide and Metformin in Combination with Honey in Pancreas of Streptozotocin-Induced Diabetic Rats" International Journal of Molecular Sciences 11, no. 5: 2056-2066. https://doi.org/10.3390/ijms11052056
APA StyleErejuwa, O. O., Sulaiman, S. A., Wahab, M. S. A., Sirajudeen, K. N. S., Salleh, M. S. M., & Gurtu, S. (2010). Antioxidant Protective Effect of Glibenclamide and Metformin in Combination with Honey in Pancreas of Streptozotocin-Induced Diabetic Rats. International Journal of Molecular Sciences, 11(5), 2056-2066. https://doi.org/10.3390/ijms11052056