Constitutive Expression of Thermobifida fusca Thermostable Acetylxylan Esterase Gene in Pichia pastoris

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Amplification and Construction of the AXE Gene in P. pastoris
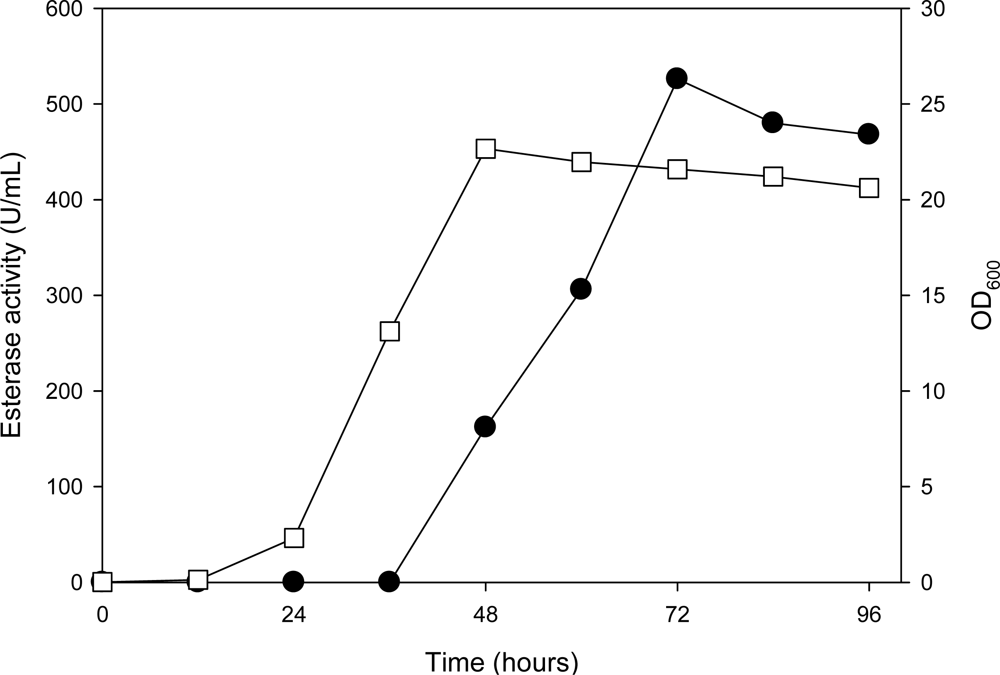
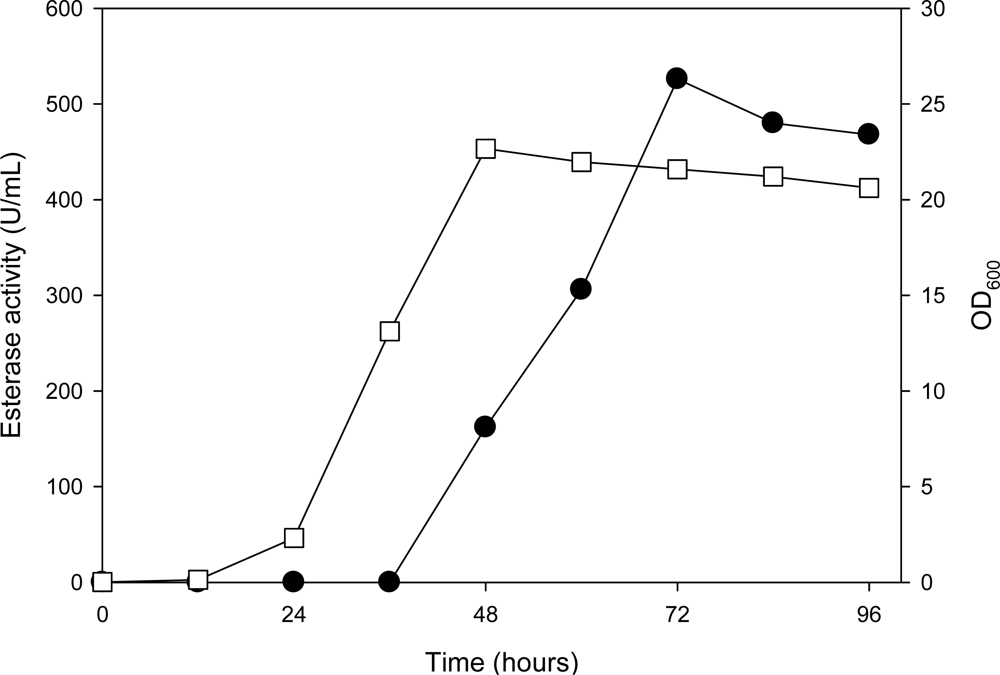
2.2. Expression of the AXE Gene (axe) in P. pastoris
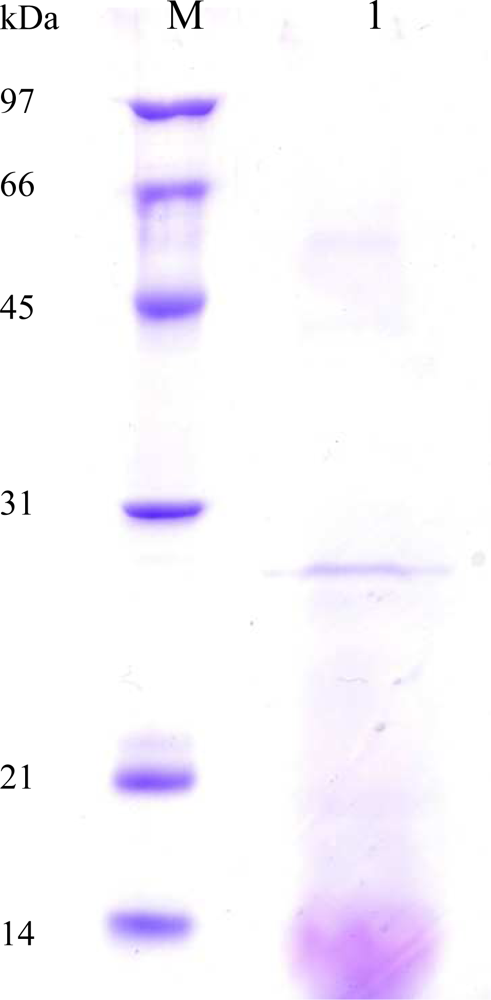
2.3. Purification of AXE from P. pastoris Transformant
2.4. Properties of AXE from P. pastoris Transformant
3. Experimental Section
3.1. Microorganisms and Vectors
3.2. Materials
3.3. Construction of the Amylase Expression Plasmid
3.4. Transformation and Screening of P. pastoris
3.5. Biomass and Esterase Activity Assay
3.6. Cultivation and Expression of AXE in Hinton Flask
3.7. Enzyme Purification
3.8. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
3.9. Deglycosylation of AXE from P. pastoris Transformant
4. Conclusions
Acknowledgments
References
- Coughlan, MP; Hazlewood, GP. β-1,4-d-Xylan-degrading enzyme systems: Biochemistry, molecular biology and applications. Appl. Biochem. Biotechnol 1993, 17, 259–289. [Google Scholar]
- Christov, LP; Prior, BA. Esterases of xylan-degrading microorganisms: Production, properties, and significance. Enzyme Microb. Technol 1993, 15, 460–475. [Google Scholar]
- Kato, Y; Nevins, DJ. Isolation and identification of O-(5-O-feruloyl-α-l-arabinofuranosyl)-1(→3)-O-β-d-xylopyranosyl-(1→4)-d-xylopyranose as a component of Zea shoot cell-walls. Carbohydr. Res 1985, 137, 139–150. [Google Scholar]
- Biely, P; Cote, GL; Kremnicky, L; Greene, RV; Tenkanen, M. Action of acetylxylan esterase from Trichoderma reesei on acetylated methyl glycosides. FEBS Lett 1997, 420, 121–124. [Google Scholar]
- Khan, AW; Lamb, KA; Overend, RP. Comparison of natural hemicellulose and chemically acetylated xylan as substrates for the determination of acetyl-xylan esterase activity in Aspergilli. Enzyme Microb. Technol 1990, 12, 127–131. [Google Scholar]
- Chavez, R; Bull, P; Eyzaguirre, J. The xylanolytic enzyme system from the genus Penicillium. J. Biotechnol 2006, 123, 413–433. [Google Scholar]
- Biely, P; MacKenzie, CR; Schneider, H. Production of acetyl xylan esterase by Trichoderma reesei and Schizophllum commune. Can. J. Microbiol 1988, 34, 767–772. [Google Scholar]
- Lee, H; To, RJ; Latta, RK; Biely, P; Schneider, H. Some properties of extracellular acetylxylan esterase produced by the yeast Rhodotorula mucilaginosa. Appl. Environ. Microbiol 1987, 53, 2831–2834. [Google Scholar]
- Dupont, C; Daigneault, N; Shareck, F; Morosoli, R; Kluepfel, D. Purification and characterization of an acetyl xylan esterase produced by Streptomyces lividans. Biochem. J 1996, 319, 881–886. [Google Scholar]
- McDermid, KP; MacKenzie, CR; Forsberg, CW. Esterase Activities of Fibrobacter succinogenes subsp. succinogenes S85. Appl. Environ. Microbiol 1990, 56, 127–132. [Google Scholar]
- Krastanova, I; Guarnaccia, C; Zahariev, S; Degrassi, G; Lamba, D. Heterologous expression, purification, crystallization, X-ray analysis and phasing of the acetyl xylan esterase from Bacillus pumilus. Biochim. Biophys. Acta 2005, 1748, 222–230. [Google Scholar]
- Li, XL; Skory, CD; Cotta, MA; Puchart, V; Biely, P. Novel family of carbohydrate esterases, based on identification of the Hypocrea jecorina acetyl esterase gene. Appl. Environ. Microbiol 2008, 74, 7482–7489. [Google Scholar]
- Liu, X; Ding, S. Molecular characterization of a new acetyl xylan esterase (AXEII) from edible straw mushroom Volvariella volvacea with both de-O-acetylation and de-N-acetylation activity. FEMS Microbiol. Lett 2009, 295, 50–56. [Google Scholar]
- Pai, CK; Wu, ZY; Chen, MJ; Zeng, YF; Chen, JW; Duan, CH; Li, ML; Liu, JR. Molecular cloning and characterization of a bifunctional xylanolytic enzyme from Neocallimastix patriciarum. Appl. Microbiol. Biotechnol 2010, 85, 1451–1462. [Google Scholar]
- Bachmann, SL; McCarthy, AJ. Purification and cooperative activity of enzymes constituting the xylan-degrading system of Thermomonospora fusca. Appl. Environ. Microbiol 1991, 57, 2121–2130. [Google Scholar]
- Liu, WH; Yang, CH. The isolation and identification of a lignocellulolytic and thermophilic actinomycete. Food Sci. Agric. Chem 2002, 4, 89–94. [Google Scholar]
- Yang, CH; Liu, WH. Purification and properties of an acetylxylan esterase from Thermobifida fusca. Enzyme Microb. Technol 2008, 42, 181–186. [Google Scholar]
- Huang, YC; Chen, GH; Chen, YF; Chen, WL; Yang, CH. Heterologous expression of thermostable acetylxylan esterase gene from Thermobifida fusca and its synergistic action with xylanase for the production of xylooligosaccharides. Biochem Biophys Res Commun 2010. [Google Scholar] [CrossRef]
- Zamost, BL; Nielsen, HK; Starnes, RL. Thermostable enzymes for industrial application. J. Ind. Microbiol Biotechnol 1991, 8, 71–82. [Google Scholar]
- Cheng, YF; Yang, CH; Liu, WH. Cloning and expression of Thermobifida xylanase gene in the methylotrophic yeast Pichia pastoris. Enzyme Microb. Technol 2005, 37, 541–546. [Google Scholar]
- Cereghino, JL; Cregg, JM. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiol. Rev 2000, 24, 45–66. [Google Scholar]
- Zhang, AL; Zhang, TY; Luo, JX; Chen, SC; Guan, WJ; Fu, CY; Peng, SQ; Li, HL. Constitutive expression of human angiostatin in Pichia pastoris by high-density cell culture. J. Ind. Microbiol. Biotechnol 2007, 34, 117–122. [Google Scholar]
- Cos, O; Ramón, R; Montesinos, JL; Valero, F. Operational strategies, monitoring and control of heterologous protein production in the methylotrophic yeast Pichia pastoris under different promoters: A review. Microb Cell Fact 2006, 5, 17. [Google Scholar] [CrossRef]
- Goodrick, JC; Xu, M; Finnegan, R; Schilling, BM; Schiavi, S; Hoppe, H; Wan, NC. High-level expression and stabilization of recombinant human chitinase produced in a continuous constitutive Pichia pastoris expression system. Biotechnol. Bioeng 2001, 74, 492–497. [Google Scholar]
- Yang, CH; Huang, YC; Chen, CY; Wen, CY. Expression of Thermobifida fusca thermostable raw starch digesting alpha-amylase in Pichia pastoris and its application in raw sago starch hydrolysis. J. Ind. Microbiol. Biotechnol 2010, 37, 401–406. [Google Scholar]
- Reddy, ST; Kumar, SN; Haas, AL; Dahms, NM. Biochemical and functional properties of the full-length cation-dependent mannose 6-phosphate receptor expressed in Pichia pastoris. Biochem. Biophys. Res. Commun 2003, 309, 643–651. [Google Scholar]
- Kademi, A; Ait-Abdelkader, N; Fakhreddine, L; Baratti, JC. A thermostable esterase activity from newly isolated moderate thermophilic bacterial strains. Enzyme Microb. Technol 1999, 24, 332–338. [Google Scholar]

{kind=link}
{kind=link}
| 0 | Total activity (U) | Total protein (mg) | Specific activity (U/mg) | Purification (fold) | Yield (%) |
|---|---|---|---|---|---|
| Culture | 469,698.8 | 637.1 | 737.2 | 1.0 | 100.0 |
| Ultrafiltration | 32,729.1 | 283.5 | 115.4 | 0.2 | 6.97 |
| Ni-Sepharose™ | 3,214.0 | 0.167 | 19,245.5 | 26.1 | 0.68 |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yang, C.-H.; Lin, K.-I.; Chen, G.-H.; Chen, Y.-F.; Chen, C.-Y.; Chen, W.-L.; Huang, Y.-C. Constitutive Expression of Thermobifida fusca Thermostable Acetylxylan Esterase Gene in Pichia pastoris. Int. J. Mol. Sci. 2010, 11, 5143-5151. https://doi.org/10.3390/ijms11125143
Yang C-H, Lin K-I, Chen G-H, Chen Y-F, Chen C-Y, Chen W-L, Huang Y-C. Constitutive Expression of Thermobifida fusca Thermostable Acetylxylan Esterase Gene in Pichia pastoris. International Journal of Molecular Sciences. 2010; 11(12):5143-5151. https://doi.org/10.3390/ijms11125143
Chicago/Turabian StyleYang, Chao-Hsun, Kun-I Lin, Gen-Hung Chen, Yu-Fen Chen, Cheng-Yu Chen, Wei-Lin Chen, and Yu-Chun Huang. 2010. "Constitutive Expression of Thermobifida fusca Thermostable Acetylxylan Esterase Gene in Pichia pastoris" International Journal of Molecular Sciences 11, no. 12: 5143-5151. https://doi.org/10.3390/ijms11125143