Copper Transporter 2 Content Is Lower in Liver and Heart of Copper-Deficient Rats

{kind=link}
Abstract
:1. Introduction
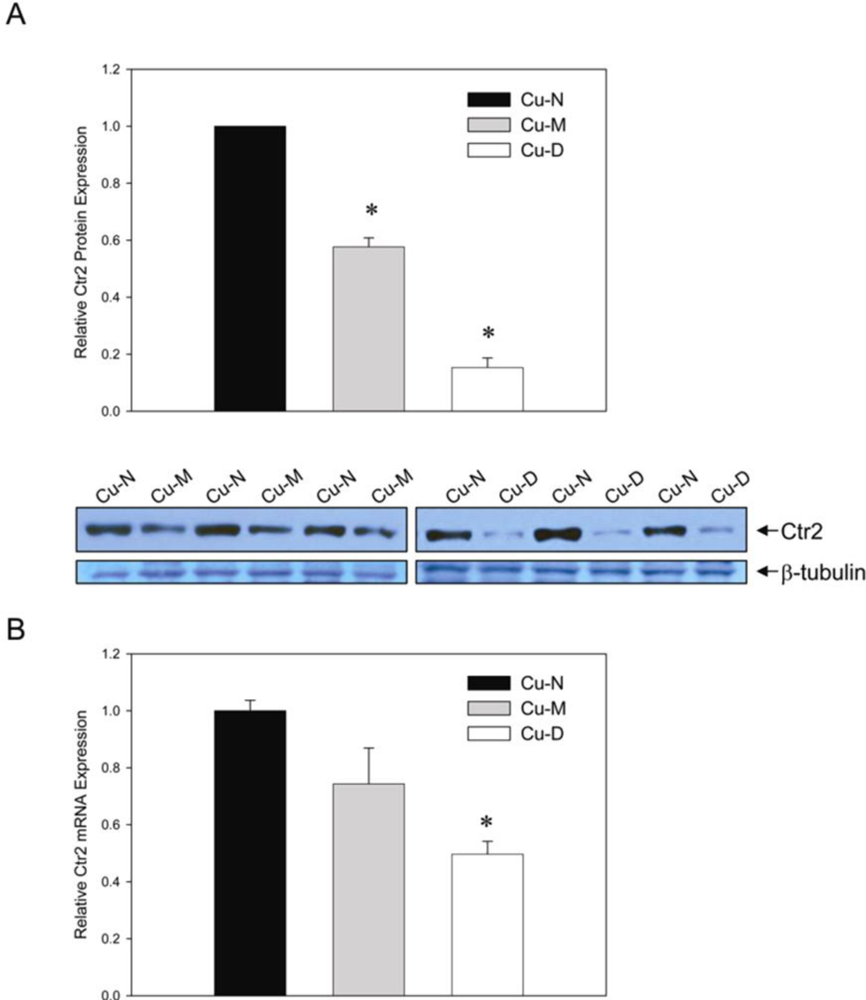
2. Results and Discussion
3. Experimental Section
3.1. Animals and Test Diets
3.2. Tissue Extracts and Western Blotting
3.3. Quantitative PCR (QPCR)
3.4. Statistical Analyses
4. Conclusions
Acknowledgments
References
- Lee, J; Pena, MM; Nose, Y; Thiele, DJ. Biochemical characterization of the human copper transporter Ctr1. J. Biol. Chem 2002, 277, 4380–4387. [Google Scholar]
- Puig, S; Lee, J; Lau, M; Thiele, DJ. Biochemical and genetic analyses of yeast and human high affinity copper transporters suggest a conserved mechanism for copper uptake. J. Biol. Chem 2002, 277, 26021–26030. [Google Scholar]
- Eisses, JF; Kaplan, JH. The mechanism of copper uptake mediated by human CTR1: a mutational analysis. J. Biol. Chem 2005, 280, 37159–37168. [Google Scholar]
- Aller, SG; Unger, VM. Projection structure of the human copper transporter CTR1 at 6-A resolution reveals a compact trimer with a novel channel-like architecture. Proc. Natl. Acad. Sci. USA 2006, 103, 3627–3632. [Google Scholar]
- van den Berghe, PV; Folmer, DE; Malingre, HE; van Beurden, E; Klomp, AE; van de Sluis, B; Merkx, M; Berger, R; Klomp, LW. Human copper transporter 2 is localized in late endosomes and lysosomes and facilitates cellular copper uptake. Biochem. J 2007, 407, 49–59. [Google Scholar]
- Rees, EM; Lee, J; Thiele, DJ. Mobilization of intracellular copper stores by the Ctr2 vacuolar copper transporter. J. Biol. Chem 2004, 279, 54221–54229. [Google Scholar]
- Bellemare, DR; Shaner, L; Morano, KA; Beaudoin, J; Langlois, R; Labbé, S. Ctr6, a vacuolar membrane copper transporter in Schizosaccharomyces pombe. J. Biol. Chem 2002, 277, 46676–46686. [Google Scholar]
- Blair, BG; Larson, CA; Adams, PL; Abada, PB; Safaei, R; Howell, SB. Regulation of copper transporter 2 expression by copper and cisplatin in human ovarian carcinoma cells. Mol. Pharmacol 2010, 77, 912–921. [Google Scholar]
- Bertinato, J; Swist, E; Plouffe, LJ; Brooks, SP; L’Abbé, MR. Ctr2 is partially localized to the plasma membrane and stimulates copper uptake in COS-7 cells. Biochem. J 2008, 409, 731–740. [Google Scholar]
- Howell, SB; Safaei, R; Larson, CA; Sailor, MJ. Copper transporters and the cellular pharmacology of the platinum-containing cancer drugs. Mol. Pharmacol 2010, 77, 887–894. [Google Scholar]
- Safaei, R; Maktabi, MH; Blair, BG; Larson, CA; Howell, SB. Effects of the loss of Atox1 on the cellular pharmacology of cisplatin. J. Inorg. Biochem 2009, 103, 333–341. [Google Scholar]
- Larson, CA; Blair, BG; Safaei, R; Howell, SB. The role of the mammalian copper transporter 1 in the cellular accumulation of platinum-based drugs. Mol. Pharmacol 2009, 75, 324–330. [Google Scholar]
- Samimi, G; Varki, NM; Wilczynski, S; Safaei, R; Alberts, DS; Howell, SB. Increase in expression of the copper transporter ATP7A during platinum drug-based treatment is associated with poor survival in ovarian cancer patients. Clin. Cancer Res 2003, 9, 5853–5859. [Google Scholar]
- Safaei, R; Otani, S; Larson, BJ; Rasmussen, ML; Howell, SB. Transport of cisplatin by the copper efflux transporter ATP7B. Mol. Pharmacol 2008, 73, 461–468. [Google Scholar]
- Liu, JJ; Jamieson, SM; Subramaniam, J; Ip, V; Jong, NN; Mercer, JF; McKeage, MJ. Neuronal expression of copper transporter 1 in rat dorsal root ganglia: association with platinum neurotoxicity. Cancer Chemother. Pharmacol 2009, 64, 847–856. [Google Scholar]
- Blair, BG; Larson, CA; Safaei, R; Howell, SB. Copper transporter 2 regulates the cellular accumulation and cytotoxicity of Cisplatin and Carboplatin. Clin. Cancer Res 2009, 15, 4312–4321. [Google Scholar]
- Blair, BG; Larson, CA; Adams, PL; Abada, PB; Pesce, CE; Safaei, R; Howell, S. Copper Transporter 2 (CTR2) regulates endocytosis and controls tumor growth and sensitivity to cisplatin in vivo. Mol Pharmacol 2010. doi:10.1124/mol.110.068411.. [Google Scholar]
- Gupta, A; Lutsenko, S. Human copper transporters: mechanism, role in human diseases and therapeutic potential. Future Med. Chem 2009, 1, 1125–1142. [Google Scholar]
- Bertinato, J; L’Abbé, MR. Maintaining copper homeostasis: regulation of copper-trafficking proteins in response to copper deficiency or overload. J. Nutr. Biochem 2004, 15, 316–322. [Google Scholar]
- Bertinato, J; L’Abbé, MR. Copper modulates the degradation of copper chaperone for Cu,Zn superoxide dismutase by the 26 S proteosome. J. Biol. Chem 2003, 278, 35071–35078. [Google Scholar]
- Caruano-Yzermans, AL; Bartnikas, TB; Gitlin, JD. Mechanisms of the copper-dependent turnover of the copper chaperone for superoxide dismutase. J. Biol. Chem 2006, 281, 13581–13587. [Google Scholar]
- Bertinato, J; Iskandar, M; L’Abbé, MR. Copper deficiency induces the upregulation of the copper chaperone for Cu/Zn superoxide dismutase in weanling male rats. J. Nutr 2003, 133, 28–31. [Google Scholar]
- Hepburn, JJ; Arthington, JD; Hansen, SL; Spears, JW; Knutson, MD. Technical note: copper chaperone for copper, zinc superoxide dismutase: a potential biomarker for copper status in cattle. J. Anim. Sci 2009, 87, 4161–4166. [Google Scholar]
- West, EC; Prohaska, JR. Cu,Zn-superoxide dismutase is lower and copper chaperone CCS is higher in erythrocytes of copper-deficient rats and mice. Exp. Biol. Med. (Maywood) 2004, 229, 756–764. [Google Scholar]
- Prohaska, JR; Broderius, M; Brokate, B. Metallochaperone for Cu,Zn-superoxide dismutase (CCS) protein but not mRNA is higher in organs from copper-deficient mice and rats. Arch. Biochem. Biophys 2003, 417, 227–234. [Google Scholar]
- Petris, MJ; Smith, K; Lee, J; Thiele, DJ. Copper-stimulated endocytosis and degradation of the human copper transporter, hCtr1. J. Biol. Chem 2003, 278, 9639–9646. [Google Scholar]
- Kuo, YM; Gybina, AA; Pyatskowit, JW; Gitschier, J; Prohaska, JR. Copper transport protein (Ctr1) levels in mice are tissue specific and dependent on copper status. J. Nutr 2006, 136, 21–26. [Google Scholar]
- Nose, Y; Wood, LK; Kim, BE; Prohaska, JR; Fry, RS; Spears, JW; Thiele, DJ. Ctr1 is an apical copper transporter in mammalian intestinal epithelial cells in vivo that is controlled at the level of protein stability. J. Biol. Chem 2010, 285, 32385–32392. [Google Scholar]
- Danks, DM. Copper deficiency in humans. Annu. Rev. Nutr 1988, 8, 235–257. [Google Scholar]
- Cordano, A. Clinical manifestations of nutritional copper deficiency in infants and children. Am. J. Clin. Nutr 1998, 67, 1012S–1016S. [Google Scholar]
- Fujita, M; Itakura, T; Takagi, Y; Okada, A. Copper deficiency during total parenteral nutrition: clinical analysis of three cases. J. Parenter. Enteral. Nutr 1989, 13, 421–425. [Google Scholar]
- Bonham, M; O’Connor, JM; Hannigan, BM; Strain, JJ. The immune system as a physiological indicator of marginal copper status? Br. J. Nutr 2002, 87, 393–403. [Google Scholar]
- Smith, AD; Botero, S; Levander, OA. Copper deficiency increases the virulence of amyocarditic and myocarditic strains of coxsackievirus B3 in mice. J. Nutr 2008, 138, 849–855. [Google Scholar]
- Hopkins, RG; Failla, ML. Chronic intake of a marginally low copper diet impairs in vitro activities of lymphocytes and neutrophils from male rats despite minimal impact on conventional indicators of copper status. J. Nutr 1995, 125, 2658–2668. [Google Scholar]
- Prohaska, JR; Lukasewycz, OA. Copper deficiency suppresses the immune response of mice. Science 1981, 213, 559–561. [Google Scholar]
- Dhar, S; Gu, FX; Langer, R; Farokhzad, OC; Lippard, SJ. Targeted delivery of cisplatin to prostate cancer cells by aptamer functionalized Pt(IV) prodrug-PLGA-PEG nanoparticles. Proc. Natl. Acad. Sci. USA 2008, 105, 17356–17361. [Google Scholar]

© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bertinato, J.; Duval, S.; L’Abbé, M.R. Copper Transporter 2 Content Is Lower in Liver and Heart of Copper-Deficient Rats. Int. J. Mol. Sci. 2010, 11, 4741-4749. https://doi.org/10.3390/ijms11114741
Bertinato J, Duval S, L’Abbé MR. Copper Transporter 2 Content Is Lower in Liver and Heart of Copper-Deficient Rats. International Journal of Molecular Sciences. 2010; 11(11):4741-4749. https://doi.org/10.3390/ijms11114741
Chicago/Turabian StyleBertinato, Jesse, Sébastien Duval, and Mary R. L’Abbé. 2010. "Copper Transporter 2 Content Is Lower in Liver and Heart of Copper-Deficient Rats" International Journal of Molecular Sciences 11, no. 11: 4741-4749. https://doi.org/10.3390/ijms11114741