Distribution of Endogenous Farnesyl Pyrophosphate and Four Species of Lysophosphatidic Acid in Rodent Brain

Abstract
:1. Introduction
2. Results and Discussion
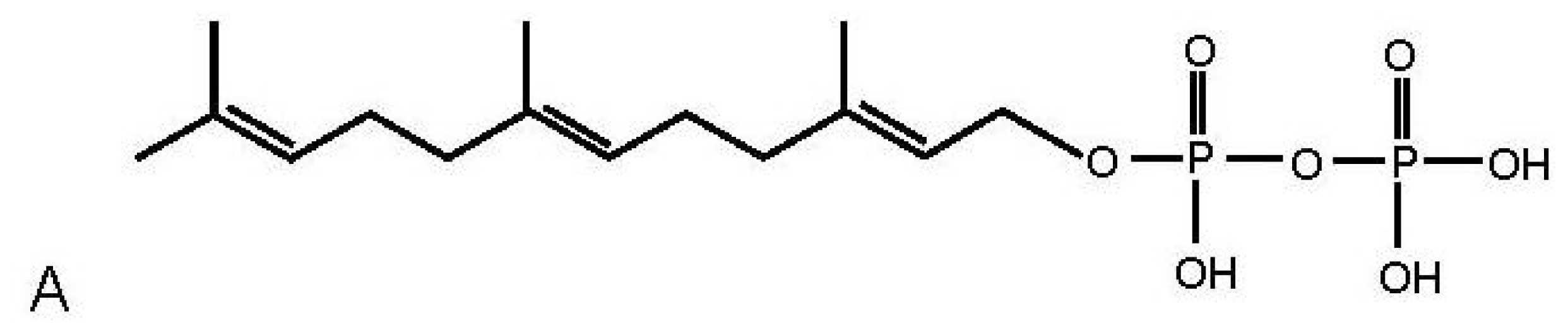
2.1. Distribution of FPP in Rat Brain
2.2. Distribution of Four LPA Species in Rat Brain
2.3. Discussion
3. Experimental Section
3.1. Sample Preparation
3.2. HPLC/MS/MS
3.2.1. FPP Analysis
3.2.2. LPA Analysis
4. Conclusions
Acknowledgements
References
- An, S; Dickens, MA; Bleu, T; Hallmark, OG; Goetzl, EJ. Molecular cloning of the human edg2 protein and its identification as a functional cellular receptor for lysophosphatidic acid. Biochem. Biophys. Res. Commun 1997, 231, 619–622. [Google Scholar]
- Hecht, JH; Weiner, JA; Post, SR; Chun, J. Ventricular zone gene-1 (vzg-1) encodes a lysophosphatidic acid receptor expressed in neurogenic regions of the developing cerebral cortex. J. Cell Biol 1996, 135, 1071–1083. [Google Scholar]
- An, S; Goetzl, EJ; Lee, H. Signaling mechanisms and molecular characteristics of G protein-coupled receptors for lysophosphatidic acid and sphingosine 1-phosphate. J. Cell Biochem. Suppl 1998, 30–31, 147–157. [Google Scholar]
- Bandoh, K; Aoki, J; Hosono, H; Kobayashi, S; Kobayashi, T; Murakami-Murofushi, K; Tsujimoto, M; Arai, H; Inoue, K. Molecular cloning and characterization of a novel human G-protein-coupled receptor, EDG7, for lysophosphatidic acid. J. Biol. Chem 1999, 274, 27776–27785. [Google Scholar]
- Im, D-S; Heise, CE; Harding, MA; George, SR; O'Dowd, BF; Theodorescu, D; Lynch, KR. Molecular cloning and characterization of a lysophosphatidic acid receptor, Edg-7, expressed in prostate. Mol. Pharm 2000, 57, 753–759. [Google Scholar]
- Lee, C-W; Rivera, R; Dubin, AE; Chun, J. LPA4/GPR23 is a lysophosphatidic acid (LPA) receptor utilizing Gs-, Gq/Gi-mediated calcium signaling and G12/13-mediated rho activation. J. Biol. Chem 2007, 282, 4310–4317. [Google Scholar]
- Lee, C-W; Rivera, R; Gardell, S; Dubin, AE; Chun, J. GPR92 as a New G12/13- and Gq-coupled Lysophosphatidic Acid Receptor That Increases cAMP, LPA5. J. Biol. Chem 2006, 281, 23589–23597. [Google Scholar]
- Moolenaar, WH. Bioactive lysophospholipids and their G protein-coupled receptors. Exp. Cell Res 1999, 253, 230–238. [Google Scholar]
- Zhang, H; Wang, D; Sun, H; Hall, RA; Yun, CC. MAGI-3 regulates LPA-induced activation of Erk and RhoA. Cell. Sign 2007, 19, 261–268. [Google Scholar]
- Wang, Q; Liu, M; Kozasa, T; Rothstein, JD; Sternweis, PC; Neubig, RR. Thrombin and lysophosphatidic acid receptors utilize distinct rhoGEFs in prostate cancer cells. J. Biol. Chem 2004, 279, 28831–28834. [Google Scholar]
- Tigyi, G; Parrill, AL. Molecular mechanisms of lysophosphatidic acid action. Prog. Lipid Res 2003, 42, 498–526. [Google Scholar]
- Kranenburg, O; Moolenaar, WH. Ras-MAP kinase signaling by lysophosphatidic acid and other G protein-coupled receptor agonists. Oncogene 2001, 20, 1540–1546. [Google Scholar]
- Oyesanya, R; Lee, Z; Wu, J; Chen, J; Song, Y; Mukherjee, A; Dent, P; Kordula, T; Zhou, H; Fang, X. Transcriptional and post-transcriptional mechanisms for lysophosphatidic acid-induced cyclooxygenase-2 expression in ovarian cancer cells. FASEB J 2008, 22, 2639–2651. [Google Scholar]
- Chen, Q; Olashaw, N; Wu, J. Participation of reactive oxygen species in the lysophosphatidic acid-stimulated mitogen-activated protein kinase kinase activation pathway. J. Biol. Chem 1995, 270, 28499. [Google Scholar]
- Kim, H; Park, S; Zhou, J; Taglialatela, G; Chung, K; Coggeshall, R; Chung, J. Reactive oxygen species (ROS) play an important role in a rat model of neuropathic pain. Pain 2004, 111, 116–124. [Google Scholar]
- Renbäck, K; Inoue, M; Ueda, H. Lysophosphatidic acid-induced, pertussis toxin-sensitive nociception through a substance P release from peripheral nerve endings in mice. Neurosci. Lett 1999, 270, 59–61. [Google Scholar]
- Pietruck, FBS; Virchow, S; Brockmeyer, N; Siffert, W. Signalling properties of lysophosphatidic acid in primary human skin fibroblasts: Role of pertussis toxin-sensitive GTP-binding proteins. Naunyn. Schmiedebergs. Arch. Pharmacol 1997, 355, 1–7. [Google Scholar]
- Inoue, M; Rashid, M; Fujita, R; Contos, J; Chun, J; Ueda, H. Initiation of neuropathic pain requires lysophosphatidic acid receptor signaling. Nat. Med 2004, 10, 712–718. [Google Scholar]
- Liliom, K; Baksa, A; Tsukahara, T; Tsukahara, R; Zelman-Femiak, M; Swiezewska, E; Tigyi, G. Farnesyl phosphates are endogenous ligands for lysophosphatidic acid receptors. Febs. J 2006, 273, 98–98. [Google Scholar]
- Westfall, D; Aboushadi, N; Shackelford, JE; Krisans, SK. Metabolism of farnesol: Phosphorylation of farnesol by rat liver microsomal and peroxisomal fractions. Biochem. Biophys. Res. Commun 1997, 230, 562–568. [Google Scholar]
- Liao, J. Isoprenoids as mediators of the biological effects of statins. Am. Soc. Clin. Investig 2002, 110, 285–288. [Google Scholar]
- Santodomingo-Garzon, T; Cunha, TM; Verri, WA, Jr; Valerio, DA; Parada, CA; Poole, S; Ferreira, SH; Cunha, FQ. Atorvastatin inhibits inflammatory hypernociception. Br. J. Pharmacol 2006, 149, 14–22. [Google Scholar]
- Oh da, Y; Yoon, JM; Moon, MJ; Hwang, JI; Choe, H; Lee, JY; Kim, JI; Kim, S; Rhim, H; O'Dell, DK; Walker, JM; Na, HS; Lee, MG; Kwon, HB; Kim, K; Seong, JY. Identification of farnesyl pyrophosphate and N-arachidonylglycine as endogenous ligands for GPR92. J. Biol. Chem 2008, 283, 21054–21064. [Google Scholar]
- Yin, H; Chu, A; Li, W; Wang, B; Shelton, F; Otero, F; Nguyen, DG; Caldwell, JS; Chen, YA. Lipid G protein-coupled receptor ligand identification using beta-arrestin PathHunter assay. J. Biol. Chem 2009, 284, 12328–12338. [Google Scholar]
- Bang, S; Yoo, S; Yang, TJ; Cho, H; Hwang, SW. Farnesyl pyrophosphate is a novel pain-producing molecule via specific activation of TRPV3. J. Biol. Chem 2010, 285, 19362–19371. [Google Scholar]
- Fields, H. Pain modulation: Expectation, opioid analgesia and virtual pain. Prog. Brain Res 2000, 122, 245. [Google Scholar]
- Walker, J; Huang, S. Endocannabinoids in pain modulation. Prostaglandins Leukot. Essent. Fatty Acids 2002, 66, 235–242. [Google Scholar]
- Hohmann, AG; Suplita, RL; Bolton, NM; Neely, MH; Fegley, D; Mangieri, R; Krey, JF; Walker, JM; Holmes, PV; Crystal, JD; Duranti, A; Tontini, A; Mor, M; Tarzia, G; Piomelli, D. An endocannabinoid mechanism for stress-induced analgesia. Nature 2005, 435, 1108–1112. [Google Scholar]
- Walker, JM; Hohmann, AG; Martin, WJ; Strangman, NM; Huang, SM; Tsou, K. The neurobiology of cannabinoid analgesia. Life Sci 1999, 65, 665–673. [Google Scholar]
- Hooff, G; Volmer, D; Wood, W; Müller, W; Eckert, G. Isoprenoid quantitation in human brain tissue: A validated HPLC–fluorescence detection method for endogenous farnesyl-(FPP) and geranylgeranylpyrophosphate (GGPP). Anal. Bioanal. Chem 2008, 392, 673–680. [Google Scholar]
- Laufs, U. Beyond lipid-lowering: Effects of statins on endothelial nitric oxide. Eur. J. Clin. Pharmacol 2003, 58, 719–731. [Google Scholar]
- Sparks, DL; Connor, DJ; Browne, PJ; Lopez, JE; Sabbagh, MN. HMG-CoA reductase inhibitors (statins) in the treatment of Alzheimer's disease and why it would be ill-advise to use one that crosses the blood-brain barrier. J. Nutr. Health Aging 2002, 6, 324–331. [Google Scholar]
- Millan, MJ. The induction of pain: An integrative review. Prog. Neurobiol 1999, 57, 1–164. [Google Scholar]
- Guo, RT; Cao, R; Liang, PH; Ko, TP; Chang, TH; Hudock, MP; Jeng, WY; Chen, CK; Zhang, Y; Song, Y; Kuo, CJ; Yin, F; Oldfield, E; Wang, AH. Bisphosphonates target multiple sites in both cis- and trans-prenyltransferases. Proc. Natl. Acad. Sci. USA 2007, 104, 10022–10027. [Google Scholar]
- Walker, K; Medhurst, SJ; Kidd, BL; Glatt, M; Bowes, M; Patel, S; McNair, K; Kesingland, A; Green, J; Chan, O; Fox, AJ; Urban, LA. Disease modifying and anti-nociceptive effects of the bisphosphonate, zoledronic acid in a model of bone cancer pain. Pain 2002, 100, 219–229. [Google Scholar]
- Ueda, H. Molecular mechanisms of neuropathic pain-phenotypic switch and initiation mechanisms. Pharmacol. Ther 2006, 109, 57–77. [Google Scholar]
- Choi, JW; Herr, DR; Noguchi, K; Yung, YC; Lee, C-W; Mutoh, T; Lin, M-E; Teo, ST; Park, KE; Mosley, AN; Chun, J. LPA Receptors: Subtypes and biological actions. Annu. Rev. Pharmacol. Toxicol 2010, 50, 157–186. [Google Scholar]
- Chen, Q; Olashaw, N; Wu, J. Participation of reactive oxygen species in the lysophosphatidic acid-stimulated mitogen-activated protein kinase kinase activation pathway. J. Biol. Chem 1995, 270, 28499–28502. [Google Scholar]
- Zeinab, K; Tao, L; Robert, DH. Free radicals contribute to the reduction in peripheral vascular responses and the maintenance of thermal hyperalgesia in rats with chronic constriction injury. Pain 1999, 79, 31–37. [Google Scholar]
- Salter, M; Strijbos, PJLM; Neale, S; Duffy, C; Follenfant, RL; Garthwaite, J. The nitric oxide-cyclic GMP pathway is required for nociceptive signalling at specific loci within the somatosensory pathway. Neuroscience 1996, 73, 649–655. [Google Scholar]
- Sun, R; Yan, J; Willis, WD. Activation of protein kinase B/Akt in the periphery contributes to pain behavior induced by capsaicin in rats. Neuroscience 2007, 144, 286–294. [Google Scholar]
- Renbäck, K; Inoue, M; Ueda, H. Lysophosphatidic acid-induced, pertussis toxin-sensitive nociception through a substance P release from peripheral nerve endings in mice. Neuroscience Letters 1999, 270, 59–61. [Google Scholar]
- Lotz, M; Vaughan, J; Carson, D. Effect of neuropeptides on production of inflammatory cytokines by human monocytes. Science 1988, 241, 1218–1221. [Google Scholar]
- Taub, DD; Oppenheim, JJ. Chemokines, inflammation and the immune system. Ther. Immunol 1994, 1, 229–246. [Google Scholar]


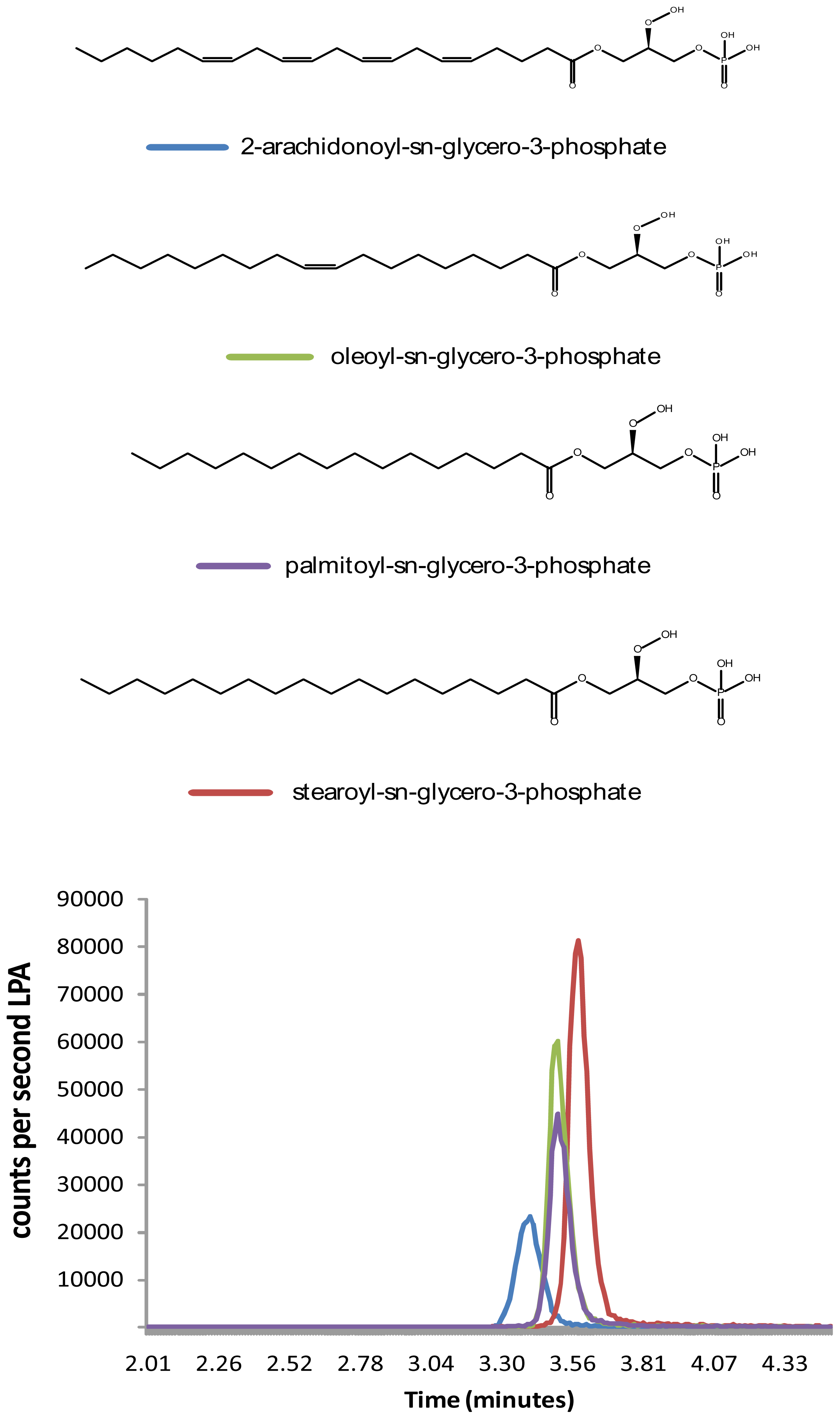

{kind=link}
{kind=link}
{kind=link}
| P-LPA in nmols/gr | A-LPA in pmols/gr | ||
|---|---|---|---|
| Hypothalamus | 4.22 ± 0.81 | Cortex | 220 ± 30.2 |
| Hippocampus | 6.16 ± 0.37 | Cerebellum | 285 ± 25.1 |
| Cortex | 6.96 ± 1.19 | Striatum | 299 ± 23.2 |
| Striatum | 7.88 ± 1.11 | Hippocampus | 316 ± 15.3 |
| Cerebellum | 13.4 ± 1.58 | Hypothalamus | 339 ± 67.6 |
| Thalamus | 19.6 ± 0.58 | Brain Stem | 364 ± 16.3 |
| Brain Stem | 25 ± 3.17 | Thalamus | 478 ± 29.6 |
| Midbrain | 28.6 ± 2.26 | Midbrain | 536 ± 41.8 |
| S-LPA in nmols/gr | O-LPA in nmols/gr | ||
| Hypothalamus | 4.34 ± 1.64 | Hypothalamus | 7.58 ± 1.83 |
| Hippocampus | 6.30 ± 0.62 | Hippocampus | 9.76 ± 1.22 |
| Striatum | 7.68 ± 0.46 | Striatum | 11.4 ± 1.01 |
| Cortex | 10.7 ± 0.55 | Cortex | 17.5 ± 3.27 |
| Cerebellum | 11.2 ± 0.51 | Thalamus | 37.8 ± 4.05 |
| Brain Stem | 13 ± 0.56 | Cerebellum | 28.1 ± 5.34 |
| Midbrain | 14 ± 1.99 | Midbrain | 63.6 ± 7.48 |
| Thalamus | 15.7 ± 2.24 | Brain Stem | 69.5 ± 4.30 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, S.H.; Raboune, S.; Walker, J.M.; Bradshaw, H.B. Distribution of Endogenous Farnesyl Pyrophosphate and Four Species of Lysophosphatidic Acid in Rodent Brain. Int. J. Mol. Sci. 2010, 11, 3965-3976. https://doi.org/10.3390/ijms11103965
Lee SH, Raboune S, Walker JM, Bradshaw HB. Distribution of Endogenous Farnesyl Pyrophosphate and Four Species of Lysophosphatidic Acid in Rodent Brain. International Journal of Molecular Sciences. 2010; 11(10):3965-3976. https://doi.org/10.3390/ijms11103965
Chicago/Turabian StyleLee, Sung Ha, Siham Raboune, J. Michael Walker, and Heather B. Bradshaw. 2010. "Distribution of Endogenous Farnesyl Pyrophosphate and Four Species of Lysophosphatidic Acid in Rodent Brain" International Journal of Molecular Sciences 11, no. 10: 3965-3976. https://doi.org/10.3390/ijms11103965