Melatonin and Structurally-Related Compounds Protect Synaptosomal Membranes from Free Radical Damage


Abstract
:
1. Introduction
2. Results and Discussion
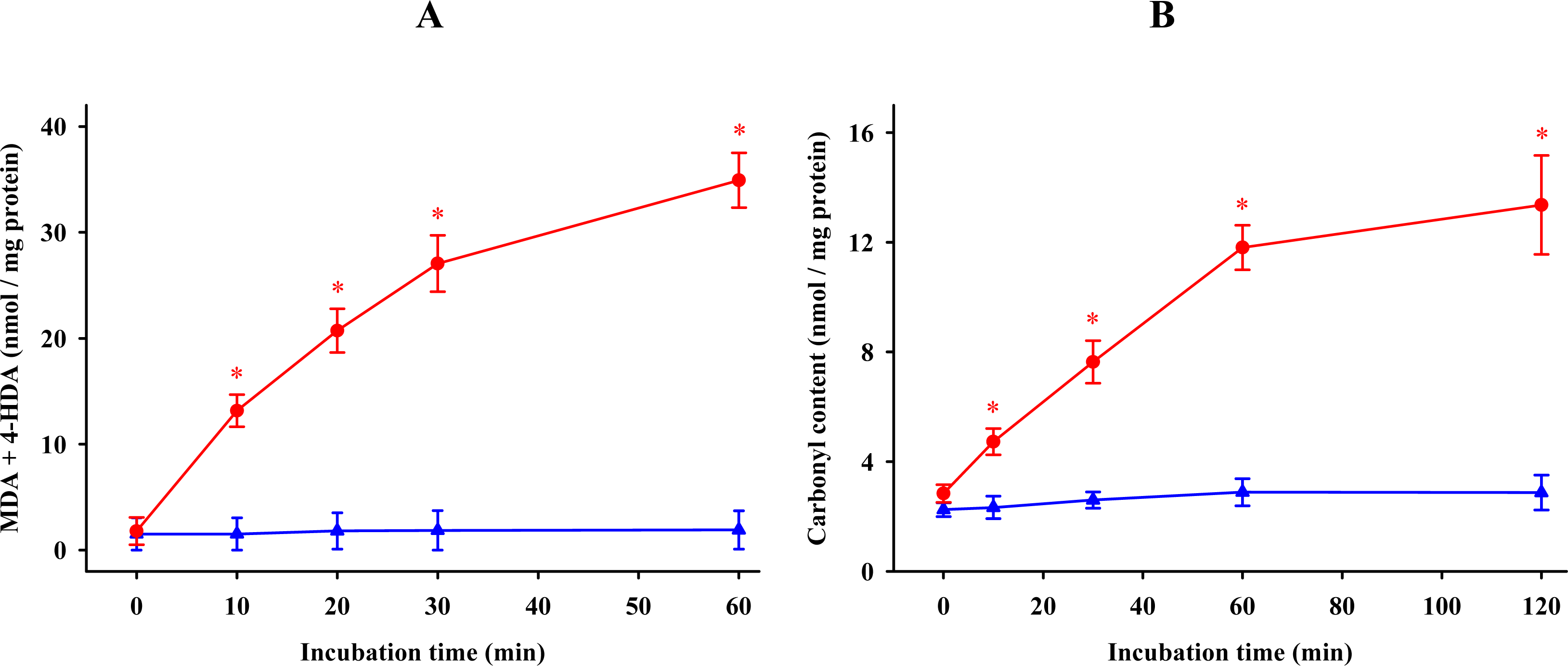
2.1. Time Kinetics of Oxidation in the Synaptosomes
2.2. Tryptophan and 5-hydroxytryptophan
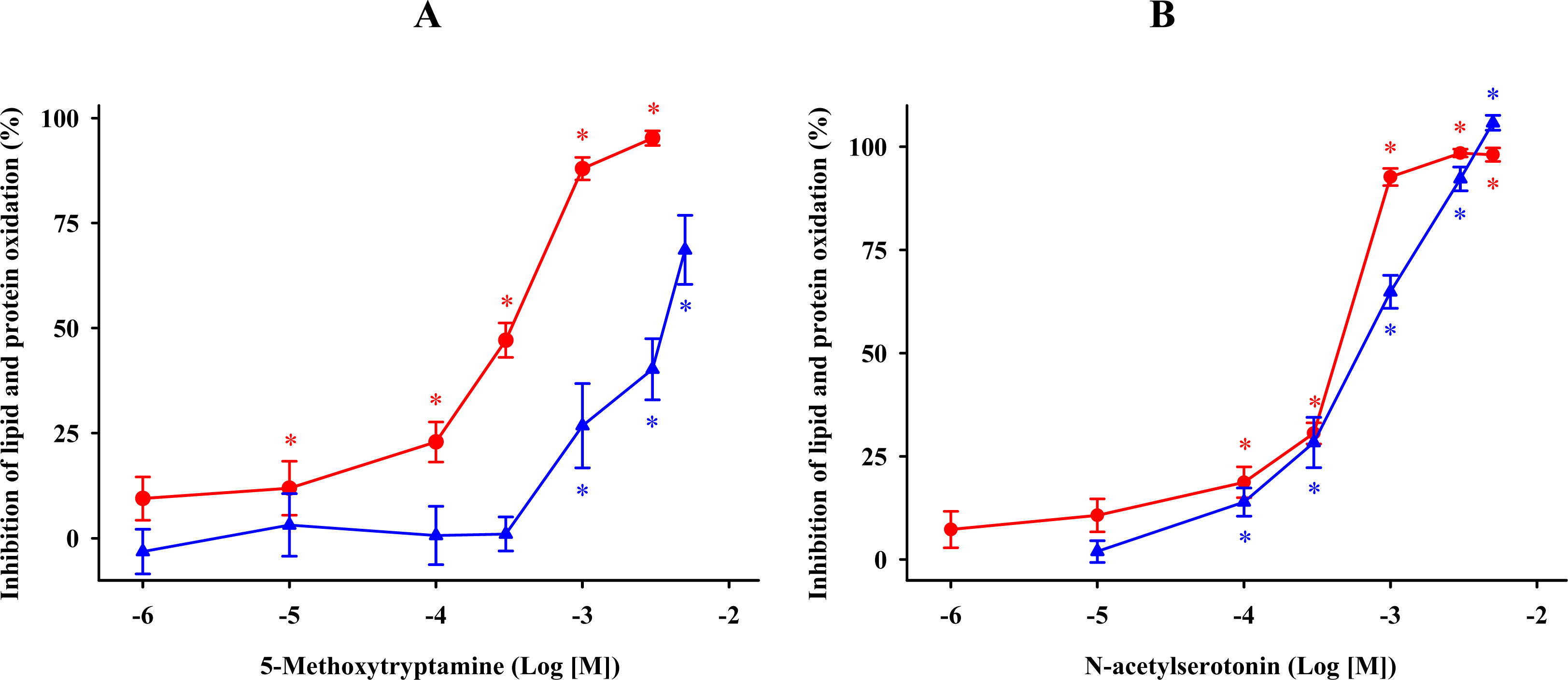
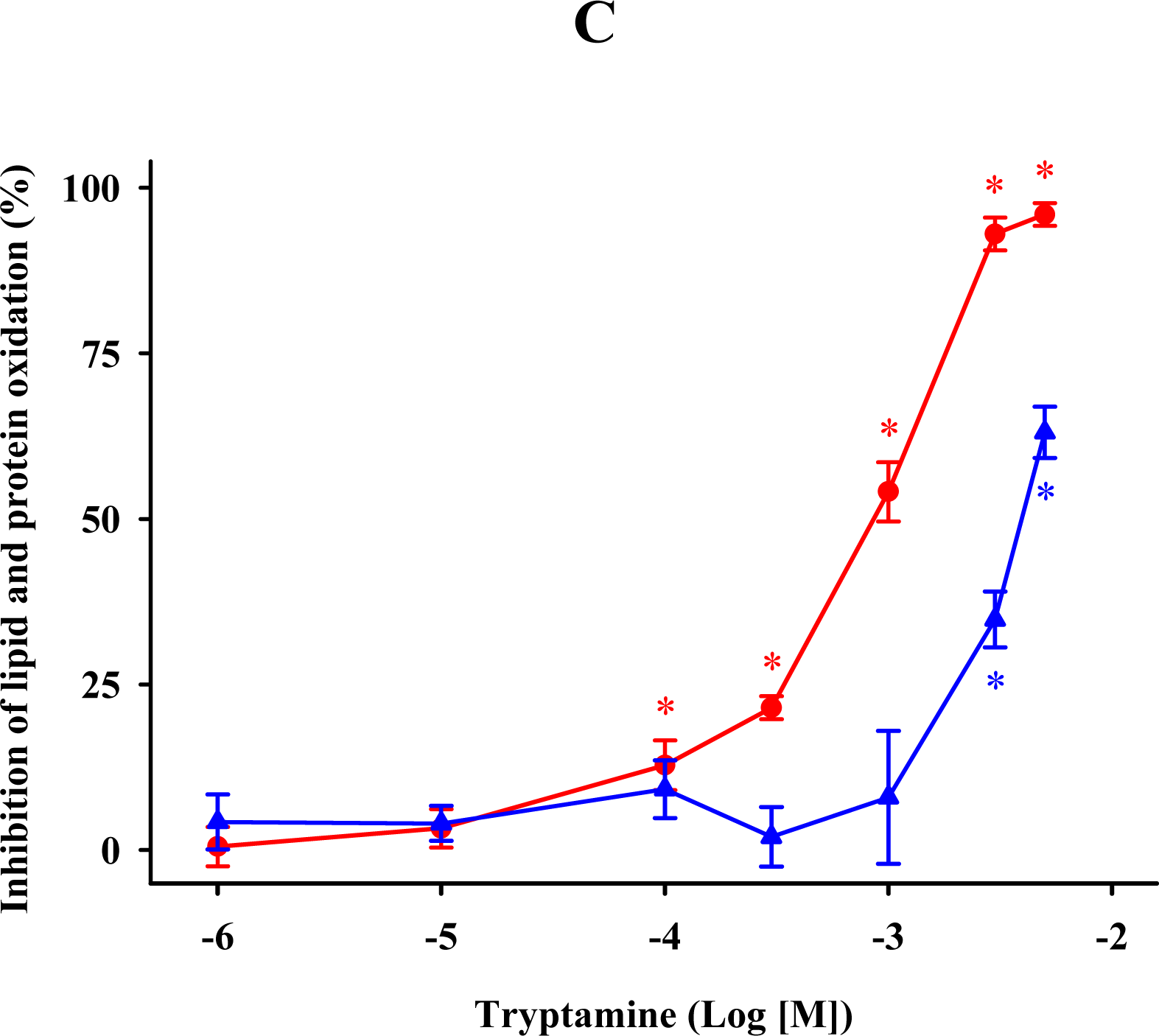
2.3. Methoxytryptamine, N-acetylserotonin, and Tryptamine
2.4. Melatonin
2.5. 5-Methoxytryptophol and 5-methoxy-3-indoleacetic Acid
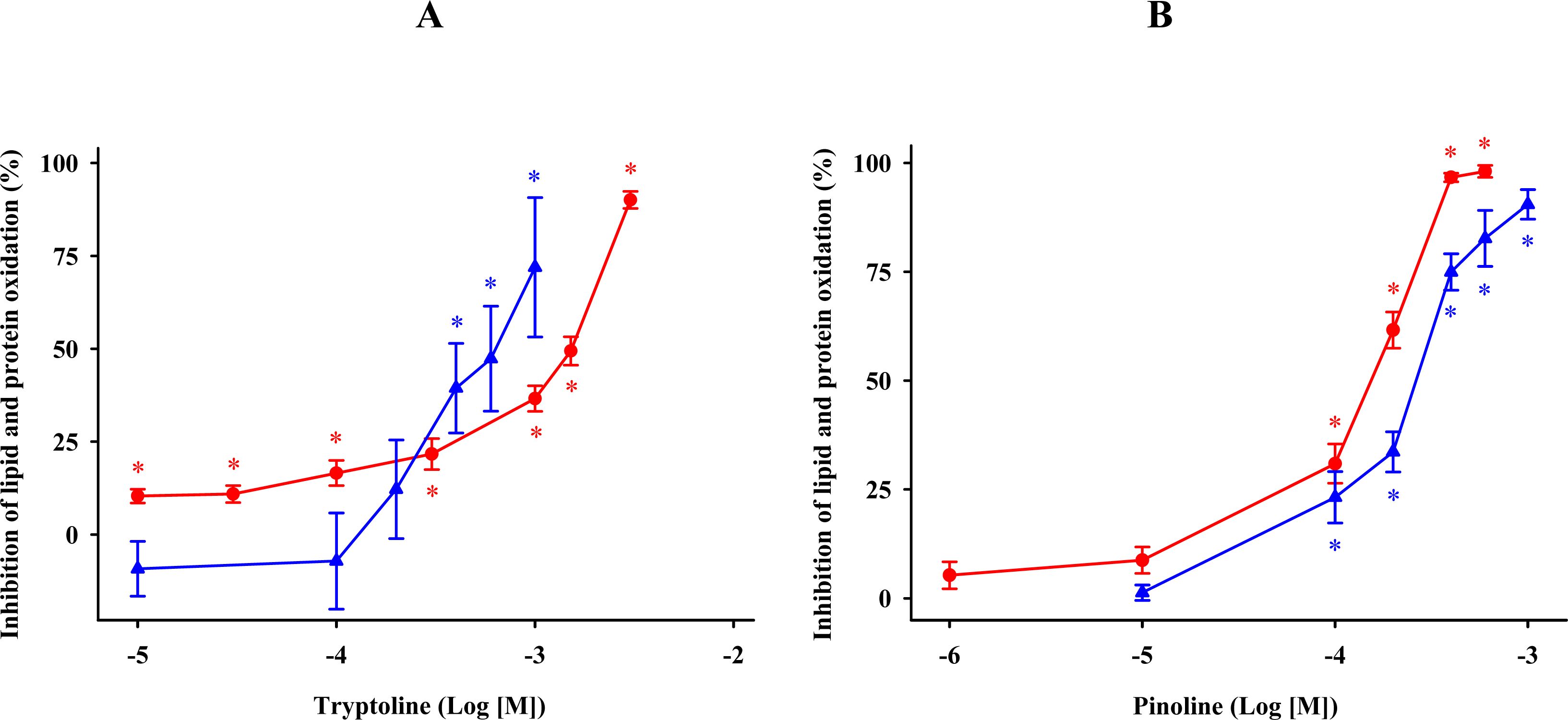
2.6. β-Carbolines
3. Experimental Section
3.1. Chemicals
3.2. Animals and Isolation of Synaptosomes
3.3. Experimental Design
3.4. Analytical Procedures
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Gutteridge, JM. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin. Chem 1995, 41, 1819–1828. [Google Scholar]
- Kanner, J; German, JB; Kinsella, JE. Initiation of lipid peroxidation in biological systems. Crit. Rev. Food Sci. Nutr 1987, 25, 317–364. [Google Scholar]
- Youngman, LD; Park, JY; Ames, BN. Protein oxidation associated with aging is reduced by dietary restriction of protein or calories. Proc. Natl. Acad. Sci. USA 1992, 89, 9112–9116. [Google Scholar]
- Davies, KJ; Delsignore, ME; Lin, SW. Protein damage and degradation by oxygen radicals. II. Modification of amino acids. J. Biol. Chem 1987, 262, 9902–9907. [Google Scholar]
- Reiter, RJ. The pineal and its hormones in the control of reproduction in mammals. Endocr. Rev 1980, 1, 109–131. [Google Scholar]
- Bubenik, GA. Localization of melatonin in the digestive tract of the rat. Effect of maturation, diurnal variation, melatonin treatment and pinealectomy. Horm. Res 1980, 12, 313–323. [Google Scholar]
- Tan, DX; Manchester, LC; Reiter, RJ; Qi, WB; Zhang, M; Weintraub, ST; Cabrera, J; Sainz, RM; Mayo, JC. Identification of highly elevated levels of melatonin in bone marrow: Its origin and significance. Biochim. Biophys. Acta 1999, 1472, 206–214. [Google Scholar]
- Guerrero, JM; Reiter, RJ. Melatonin-immune system relationships. Curr. Top Med. Chem 2002, 2, 167–179. [Google Scholar]
- Reiter, RJ. Oxidative damage in the central nervous system: Protection by melatonin. Prog. Neurobiol 1998, 56, 359–384. [Google Scholar]
- Reiter, RJ; Korkmaz, A; Paredes, SD; Manchester, LC; Tan, DX. Melatonin reduces oxidative/nitrosative stress due to drugs, toxins, metals, and herbicides. Neuro Endocrinol. Lett 2008, 29, 609–613. [Google Scholar]
- García, JJ; Reiter, RJ; Guerrero, JM; Escames, G; Yu, BP; Oh, CS; Muñoz-Hoyos, A. Melatonin prevents changes in microsomal membrane fluidity during induced lipid peroxidation. FEBS Lett 1997, 408, 297–300. [Google Scholar]
- Tan, DX; Chen, LD; Poeggeler, B; Manchester, LC; Reiter, RJ. Melatonin: A potent, endogenous hydroxyl radical scavenger. Endocr. J 1993, 1, 57–60. [Google Scholar]
- Cagnoli, CM; Atabay, C; Kharlamova, E; Manev, H. Melatonin protects neurons from singlet oxygen-induced apoptosis. J. Pineal. Res 1995, 18, 222–226. [Google Scholar]
- Tan, DX; Manchester, LC; Reiter, RJ; Plummer, BF; Hardies, LJ; Weintraub, ST; Vijayalaxmi; Shepherd, AM. A novel melatonin metabolite, cyclic 3-hydroxymelatonin: A biomarker of in vivo hydroxyl radical generation. Biochem. Biophys. Res. Commun 1998, 253, 614–620. [Google Scholar]
- Dellegar, SM; Murphy, SA; Bourne, AE; DiCesare, JC; Purser, GH. Identification of the factors affecting the rate of deactivation of hypochlorous acid by melatonin. Biochem. Biophys. Res. Commun 1999, 257, 431–439. [Google Scholar]
- Zhang, H; Squadrito, GL; Uppu, R; Pryor, WA. Reaction of peroxynitrite with melatonin: A mechanistic study. Chem. Res. Toxicol 1999, 12, 526–534. [Google Scholar]
- Tan, DX; Manchester, LC; Reiter, RJ; Plummer, BF; Limson, J; Weintraub, ST; Qi, W. Melatonin directly scavenges hydrogen peroxide: A potentially new metabolic pathway of melatonin biotransformation. Free Radic. Biol. Med 2000, 29, 1177–1185. [Google Scholar]
- Antolín, I; Rodríguez, C; Sainz, RM; Mayo, JC; Uria, H; Kotler, ML; Rodríguez-Colunga, MJ; Tolivia, D; Menéndez-Peláez, A. Neurohormone melatonin prevents cell damage: Effect on gene expression for antioxidant enzymes. FASEB J 1996, 10, 882–890. [Google Scholar]
- Coto-Montes, A; Hardeland, R. Antioxidative effects of melatonin in Drosophila melanogaster: Antagonization of damage induced by the inhibition of catalase. J. Pineal. Res 1999, 27, 154–158. [Google Scholar]
- Pozo, D; Reiter, RJ; Calvo, JR; Guerrero, JM. Inhibition of cerebellar nitric oxide synthase and cyclic GMP production by melatonin via complex formation with calmodulin. J. Cell Biochem 1997, 65, 430–442. [Google Scholar]
- Sinet, PM; Heikkila, RE; Cohen, G. Hydrogen peroxide production by rat brain in vivo. J. Neurochem 1980, 34, 1421–1428. [Google Scholar]
- Beard, JL; Connor, JR; Jones, BC. Iron in the brain. Nutr. Rev 1993, 51, 157–170. [Google Scholar]
- Cadenas, E; Simic, MG; Sies, H. Antioxidant activity of 5-hydroxytryptophan, 5-hydroxyindole, and DOPA against microsomal lipid peroxidation and its dependence on vitamin E. Free Radic. Res. Commun 1989, 6, 11–17. [Google Scholar]
- Millán-Plano, S; García, JJ; Martínez-Ballarín, E; Reiter, RJ; Ortega-Gutiérrez, S; Lázaro, RM; Escanero, JF. Melatonin and pinoline prevent aluminium-induced lipid peroxidation in rat synaptosomes. J. Trace Elem. Med. Biol 2003, 17, 39–44. [Google Scholar]
- Reyes-Gonzales, MC; Fuentes-Broto, L; Martínez-Ballarín, E; Miana-Mena, FJ; Berzosa, C; García-Gil, FA; Aranda, M; García, JJ. Effects of tryptophan and 5-hydroxytryptophan on the hepatic cell membrane rigidity due to oxidative stress. J. Membr. Biol 2009, 231, 93–99. [Google Scholar]
- Feksa, LR; Latini, A; Rech, VC; Wajner, M; Dutra-Filho, CS; de Souza Wyse, AT; Wannmacher, CM. Promotion of oxidative stress by L-tryptophan in cerebral cortex of rats. Neurochem. Int 2006, 49, 87–93. [Google Scholar]
- Mathur, RL; Sahai, P. Effects of a tryptophan supplemented diet and U.V. radiation on the rat lens. Lens Eye Toxic. Res 1990, 7, 143–160. [Google Scholar]
- Aviram, M; Cogan, U; Mokady, S. Excessive dietary tryptophan enhances plasma lipid peroxidation in rats. Atherosclerosis 1991, 88, 29–34. [Google Scholar]
- Poeggeler, B; Reiter, RJ; Hardeland, R; Tan, DX; Barlow-Walden, LR. Melatonin and structurally-related, endogenous indoles act as potent electron donors and radical scavengers in vitro. Redox Rep 1996, 2, 179–184. [Google Scholar]
- Jaworek, J; Leja-Szpak, A; Bonior, J; Nawrot, K; Tomaszewska, R; Stachura, J; Sendur, R; Pawlik, W; Brzozowski, T; Konturek, SJ. Protective effect of melatonin and its precursor L-tryptophan on acute pancreatitis induced by caerulein overstimulation or ischemia/reperfusion. J. Pineal. Res 2003, 34, 40–52. [Google Scholar]
- Brzozowski, T; Konturek, PC; Konturek, SJ; Pajdo, R; Bielanski, W; Brzozowska, I; Stachura, J; Hahn, EG. The role of melatonin and L-tryptophan in prevention of acute gastric lesions induced by stress, ethanol, ischemia, and aspirin. J. Pineal. Res 1997, 23, 79–89. [Google Scholar]
- Brzozowska, I; Konturek, PC; Brzozowski, T; Konturek, SJ; Kwiecien, S; Pajdo, R; Drozdowicz, D; Pawlik, M; Ptak, A; Hahn, EG. Role of prostaglandins, nitric oxide, sensory nerves and gastrin in acceleration of ulcer healing by melatonin and its precursor, l-tryptophan. J. Pineal. Res 2002, 32, 149–162. [Google Scholar]
- Uchida, K; Enomoto, N; Itakura, K; Kawakishi, S. Formation of diastereoisomeric 3a-hydroxypyrroloindoles from a tryptophan residue analog mediated by iron (II)-EDTA and L-ascorbate. Arch. Biochem. Biophys 1990, 279, 14–20. [Google Scholar]
- Chan, TY; Tang, PL. Characterization of the antioxidant effects of melatonin and related indoleamine in vitro. J. Pineal. Res 1996, 20, 187–191. [Google Scholar]
- Ng, TB; Liu, F; Zhao, L. Antioxidative and free radical scavenging activities of pineal indoles. J. Neural. Transm 2000, 107, 1243–1251. [Google Scholar]
- Wang, HX; Liu, F; Ng, TB. Examination of pineal indoles and 6-methoxy-2-benzoxazolinone for antioxidant and antimicrobial effects. Comp. Biochem. Physiol. C: Toxicol. Pharmacol 2001, 130, 379–388. [Google Scholar]
- Hara, M; Iigo, M; Ohtani-Kaneko, R; Nakamura, N; Suzuki, T; Reiter, RJ; Hirata, K. Administration of melatonin and related indoles prevents exercise-induced cellular oxidative changes in rats. Biol. Signals 1997, 6, 90–100. [Google Scholar]
- Siraki, AG; O'Brien, PJ. Prooxidant activity of free radicals derived from phenol-containing neurotransmitters. Toxicology 2002, 177, 81–90. [Google Scholar]
- Seegar, H; Mueck, AO; Lippert, TH. Effect of melatonin and metabolites on copper-mediated oxidation of flow density lipoprotein. Br. J. Clin. Pharmacol 1997, 44, 283–284. [Google Scholar]
- Gozzo, A; Lesieur, D; Duriez, P; Fruchart, JC; Teissier, E. Structure-activity relationships in a series of melatonin analogues with the low-density lipoprotein oxidation model. Free Radic. Biol. Med 1999, 26, 1538–1543. [Google Scholar]
- Qi, W; Reiter, RJ; Tan, DX; Manchester, LC; Siu, AW; García, JJ. Increased levels of oxidatively damaged DNA induced by chromium(III) and H2O2: Protection by melatonin and related molecules. J. Pineal. Res 2000, 29, 54–61. [Google Scholar]
- Longoni, B; Pryor, WA; Marchiafava, P. Inhibition of lipid peroxidation by N-acetylserotonin and its role in retinal physiology. Biochem. Biophys. Res. Commun 1997, 233, 778–780. [Google Scholar]
- Lezoualc'h, F; Sparapani, M; Behl, C. N-acetyl-serotonin (normelatonin) and melatonin protect neurons against oxidative challenges and suppress the activity of the transcription factor NF-kappaB. J. Pineal. Res 1998, 24, 168–178. [Google Scholar]
- Siu, AW; Reiter, RJ; To, CH. Pineal indoleamines and vitamin E reduce nitric oxide-induced lipid peroxidation in rat retinal homogenates. J. Pineal. Res 1999, 27, 122–128. [Google Scholar]
- García, JJ; Reiter, RJ; Karbownik, M; Calvo, JR; Ortiz, GG; Tan, DX; Martínez-Ballarín, E; Acuña-Castroviejo, D. N-acetylserotonin suppresses hepatic microsomal membrane rigidity associated with lipid peroxidation. Eur. J. Pharmacol 2001, 428, 169–175. [Google Scholar]
- Yildirim, N; Ozer, A; Inal, M; Angin, K; Yurdakul, S. The effect of N-acetyl serotonin on ultraviolet-radiation-induced cataracts in rats. Ophthalmologica 2003, 217, 148–153. [Google Scholar]
- Calvo, JR; Reiter, RJ; García, JJ; Ortiz, GG; Tan, DX; Karbownik, M. Characterization of the protective effects of melatonin and related indoles against alpha-naphthylisothiocyanate-induced liver injury in rats. J. Cell Biochem 2001, 80, 461–470. [Google Scholar]
- Stetinova, V; Smetanova, L; Grossmann, V; Anzenbacher, P. In vitro and in vivo assessment of the antioxidant activity of melatonin and related indole derivatives. Gen. Physiol. Biophys 2002, 21, 153–162. [Google Scholar]
- Nakagawa, H; Takusagawa, M; Arima, H; Furukawa, K; Kinoshita, T; Ozawa, T; Ikota, N. Selective scavenging property of the indole moiety for the nitrating species of peroxynitrite. Chem. Pharm. Bull (Tokyo) 2004, 52, 146–149. [Google Scholar]
- Herrera, F; Sainz, RM; Mayo, JC; Martín, V; Antolín, I; Rodríguez, C. Glutamate induces oxidative stress not mediated by glutamate receptors or cystine transporters: Protective effect of melatonin and other antioxidants. J. Pineal. Res 2001, 31, 356–362. [Google Scholar]
- Tan, DX; Manchester, LC; Reiter, RJ; Qi, W; Kim, SJ; El-Sokkary, GH. Melatonin protects hippocampal neurons in vivo against kainic acid-induced damage in mice. J. Neurosci. Res 1998, 54, 382–389. [Google Scholar]
- Cabrera, J; Reiter, RJ; Tan, DX; Qi, W; Sainz, RM; Mayo, JC; García, JJ; Kim, SJ; El-Sokkary, G. Melatonin reduces oxidative neurotoxicity due to quinolinic acid: In vitro and in vivo findings. Neuropharmacology 2000, 39, 507–514. [Google Scholar]
- Tunez, I; Muñoz, C; Feijoo, M; Muñoz-Castaneda, JR; Bujalance, I; Valdelvira, ME; Montilla López, P. Protective melatonin effect on oxidative stress induced by okadaic acid into rat brain. J. Pineal. Res 2003, 34, 265–268. [Google Scholar]
- Pappolla, MA; Sos, M; Omar, RA; Bick, RJ; Hickson-Bick, DL; Reiter, RJ; Efthimiopoulos, S; Robakis, NK. Melatonin prevents death of neuroblastoma cells exposed to the Alzheimer amyloid peptide. J. Neurosci 1997, 17, 1683–1690. [Google Scholar]
- Acuña-Castroviejo, D; Coto-Montes, A; Gaia Monti, M; Ortiz, GG; Reiter, RJ. Melatonin is protective against MPTP-induced striatal and hippocampal lesions. Life Sci 1997, 60, PL23–PL29. [Google Scholar]
- Ortega-Gutiérrez, S; Fuentes-Broto, L; García, JJ; López-Vicente, M; Martínez-Ballarín, E; Miana-Mena, FJ; Millán-Plano, S; Reiter, RJ. Melatonin reduces protein and lipid oxidative damage induced by homocysteine in rat brain homogenates. J. Cell Biochem 2007, 102, 729–735. [Google Scholar]
- Sewerynek, E; Melchiorri, D; Reiter, RJ; Ortiz, GG; Lewinski, A. Lipopolysaccharide-induced hepatotoxicity is inhibited by the antioxidant melatonin. Eur. J. Pharmacol 1995, 293, 327–334. [Google Scholar]
- Sener, G; Tosun, O; Sehirli, AO; Kacmaz, A; Arbak, S; Ersoy, Y; Ayanoglu-Dulger, G. Melatonin and N-acetylcysteine have beneficial effects during hepatic ischemia and reperfusion. Life Sci 2003, 72, 2707–2718. [Google Scholar]
- Rodríguez, C; Mayo, JC; Sainz, RM; Antolín, I; Herrera, F; Martín, V; Reiter, RJ. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal. Res 2004, 36, 1–9. [Google Scholar]
- Pinol-Ripoll, G; Fuentes-Broto, L; Millán-Plano, S; Reyes-Gonzales, M; Mauri, JA; Martínez-Ballarín, E; Reiter, RJ; García, JJ. Protective effect of melatonin and pinoline on nitric oxide-induced lipid and protein peroxidation in rat brain homogenates. Neurosci. Lett 2006, 405, 89–93. [Google Scholar]
- Albendea, CD; Gómez-Trullén, EM; Fuentes-Broto, L; Miana-Mena, FJ; Millán-Plano, S; Reyes-Gonzales, MC; Martínez-Ballarín, E; García, JJ. Melatonin reduces lipid and protein oxidative damage in synaptosomes due to aluminium. J. Trace Elem. Med. Biol 2007, 21, 261–268. [Google Scholar]
- Kim, SJ; Reiter, RJ; Qi, W; Tan, DX; Cabrera, J. Melatonin prevents oxidative damage to protein and lipid induced by ascorbate-Fe3+-EDTA: Comparison with glutathione and alpha-tocopherol. Neuro Endocrinol. Lett 2000, 21, 269–276. [Google Scholar]
- Okatani, Y; Wakatsuki, A; Reiter, RJ; Miyahara, Y. Melatonin reduces oxidative damage of neural lipids and proteins in senescence-accelerated mouse. Neurobiol. Aging 2002, 23, 639–644. [Google Scholar]
- Hardeland, R; Reiter, RJ; Poeggeler, B; Tan, DX. The significance of the metabolism of the neurohormone melatonin: Antioxidative protection and formation of bioactive substances. Neurosci. Biobehav. Rev 1993, 17, 347–357. [Google Scholar]
- García, JJ; Reiter, RJ; Cabrera, JJ; Pie, J; Mayo, JC; Sainz, RM; Tan, DX; Qi, W; Acuña-Castroviejo, D. 5-methoxytryptophol preserves hepatic microsomal membrane fluidity during oxidative stress. J. Cell Biochem 2000, 76, 651–657. [Google Scholar]
- Ortega-Gutiérrez, S; García, JJ; Martínez-Ballarín, E; Reiter, RJ; Millán-Plano, S; Robinson, M; Acuña-Castroviejo, D. Melatonin improves deferoxamine antioxidant activity in protecting against lipid peroxidation caused by hydrogen peroxide in rat brain homogenates. Neurosci. Lett 2002, 323, 55–59. [Google Scholar]
- Callaway, JC; Gynther, J; Poso, A; Vepsalainen, J; Airaksinen, MM. The Pictet-Spengler reaction and biogenic tryptamines: Formation of tetrahydro-beta-carbolines at physicological pH. J. Heterocyclic. Chem 1994, 31, 431–435. [Google Scholar]
- García, JJ; Martínez-Ballarín, E; Robinson, M; Allué, JL; Reiter, RJ; Osuna, C; Acuña-Castroviejo, D. Protective effect of beta-carbolines and other antioxidants on lipid peroxidation due to hydrogen peroxide in rat brain homogenates. Neurosci. Lett 2000, 294, 1–4. [Google Scholar]
- García, JJ; Reiter, RJ; Pié, J; Ortiz, GG; Cabrera, J; Sainz, RM; Acuñ-Castroviejo, D. Role of pinoline and melatonin in stabilizing hepatic microsomal membranes against oxidative stress. J. Bioenerg. Biomembr 1999, 31, 609–616. [Google Scholar]
- Pless, G; Frederiksen, TJ; García, JJ; Reiter, RJ. Pharmacological aspects of N-acetyl-5-methoxytryptamine (melatonin) and 6-methoxy-1,2,3,4-tetrahydro-beta-carboline (pinoline) as antioxidants: Reduction of oxidative damage in brain region homogenates. J. Pineal. Res 1999, 26, 236–246. [Google Scholar]
- Tang, GY; Ip, AK; Siu, AW. Pinoline and N-acetylserotonin reduce glutamate-induced lipid peroxidation in retinal homogenates. Neurosci. Lett 2007, 412, 191–194. [Google Scholar]
- Wong, SF; Halliwell, B; Richmond, R; Skowroneck, WR. The role of superoxide and hydroxyl radicals in the degradation of hyaluronic acid induced by metal ions and by ascorbic acid. J. Inorg. Biochem 1981, 14, 127–134. [Google Scholar]
- Sahu, SC; Washington, MC. Effect of ascorbic acid and curcumin on quercetin-induced nuclear DNA damage, lipid peroxidation and protein degradation. Cancer Lett 1992, 63, 237–241. [Google Scholar]
- Janero, DR. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med 1990, 9, 515–540. [Google Scholar]
- Lowry, OH; Rosebrough, NJ; Farr, AL; Randall, RJ. Protein measurement with the Folin phenol reagent. J. Biol. Chem 1951, 193, 265–275. [Google Scholar]
- Levine, RL; Garland, D; Oliver, CN; Amici, A; Climent, I; Lenz, AG; Ahn, BW; Shaltiel, S; Stadtman, ER. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol 1990, 186, 464–478. [Google Scholar]
 ) or absence (
) or absence (  ) of 0.1 mM FeCl3 and 0.1 mM ascorbic acid. Results are given as means ± SE (n = 6). * P < 0.05, compared to controls at the same time.
) or absence ( ) of 0.1 mM FeCl3 and 0.1 mM ascorbic acid. Results are given as means ± SE (n = 6). * P < 0.05, compared to controls at the same time.
) of 0.1 mM FeCl3 and 0.1 mM ascorbic acid. Results are given as means ± SE (n = 6). * P < 0.05, compared to controls at the same time.
) or absence ( ) of 0.1 mM FeCl3 and 0.1 mM ascorbic acid. Results are given as means ± SE (n = 6). * P < 0.05, compared to controls at the same time.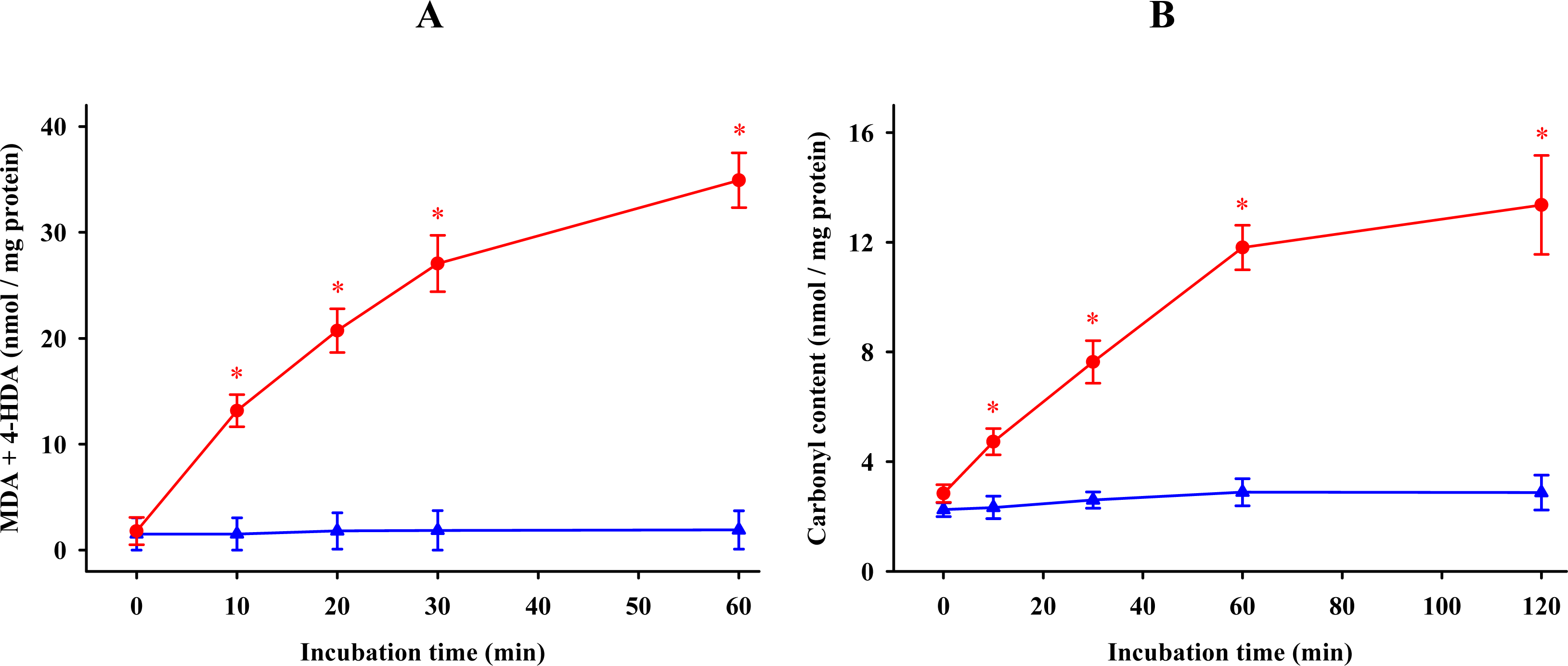 ) and protein ( ) oxidations in synaptosomes isolated from rat brains. The incubation time was 30 min for lipid peroxidation and 1 h for protein oxidation. Both indoleamines are expressed as the logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are expressed as a percentage of control membrane preparations. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidations in synaptosomes isolated from rat brains. The incubation time was 30 min for lipid peroxidation and 1 h for protein oxidation. Both indoleamines are expressed as the logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are expressed as a percentage of control membrane preparations. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidations in synaptosomes isolated from rat brains. The incubation time was 30 min for lipid peroxidation and 1 h for protein oxidation. Both indoleamines are expressed as the logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are expressed as a percentage of control membrane preparations. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidations in synaptosomes isolated from rat brains. The incubation time was 30 min for lipid peroxidation and 1 h for protein oxidation. Both indoleamines are expressed as the logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are expressed as a percentage of control membrane preparations. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.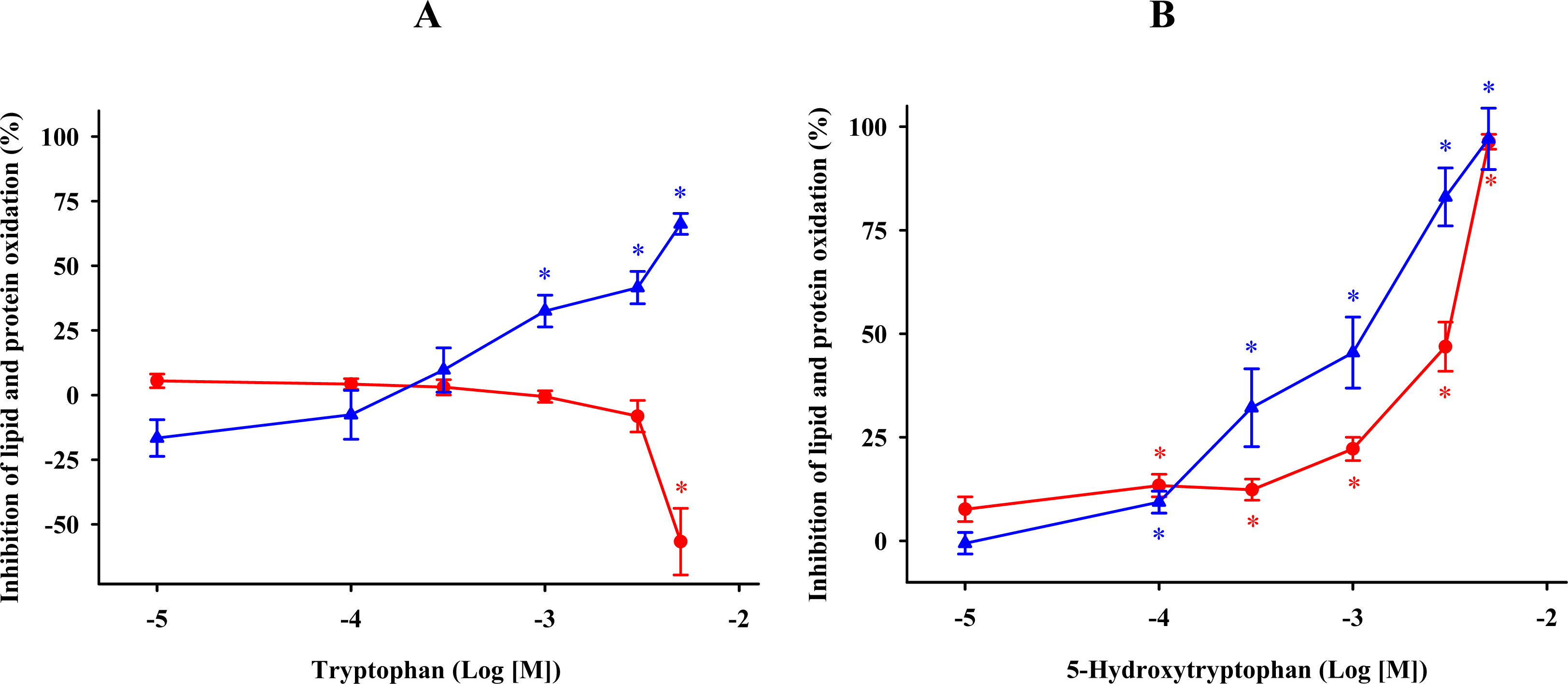 ) and protein ( ) oxidations in synaptosomal membranes. 5-methoxytryptamine, N-acetylserotonin and tryptamine are expressed as a logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are normalized with respect to control membrane preparations that were not exposed to oxidative conditions. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidations in synaptosomal membranes. 5-methoxytryptamine, N-acetylserotonin and tryptamine are expressed as a logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are normalized with respect to control membrane preparations that were not exposed to oxidative conditions. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidations in synaptosomal membranes. 5-methoxytryptamine, N-acetylserotonin and tryptamine are expressed as a logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are normalized with respect to control membrane preparations that were not exposed to oxidative conditions. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidations in synaptosomal membranes. 5-methoxytryptamine, N-acetylserotonin and tryptamine are expressed as a logarithm of its molar concentration. Results are given as means ± SE (n = 6) and are normalized with respect to control membrane preparations that were not exposed to oxidative conditions. * P < 0.05 vs. membranes treated only with FeCl3 and ascorbic acid.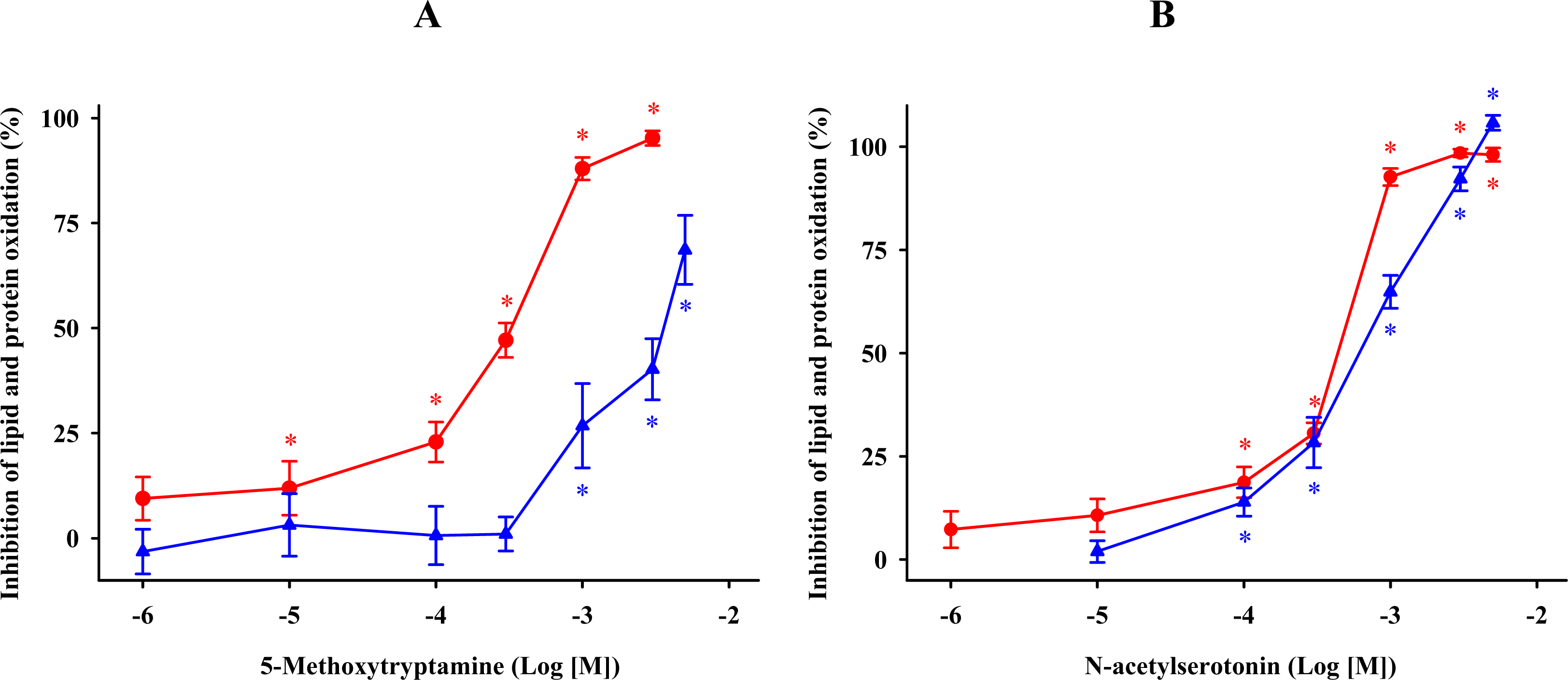
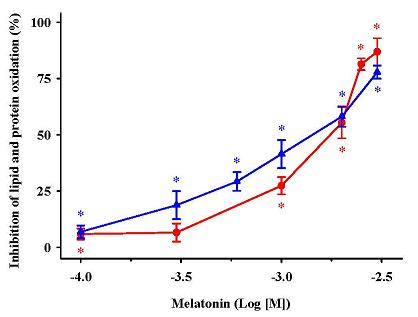
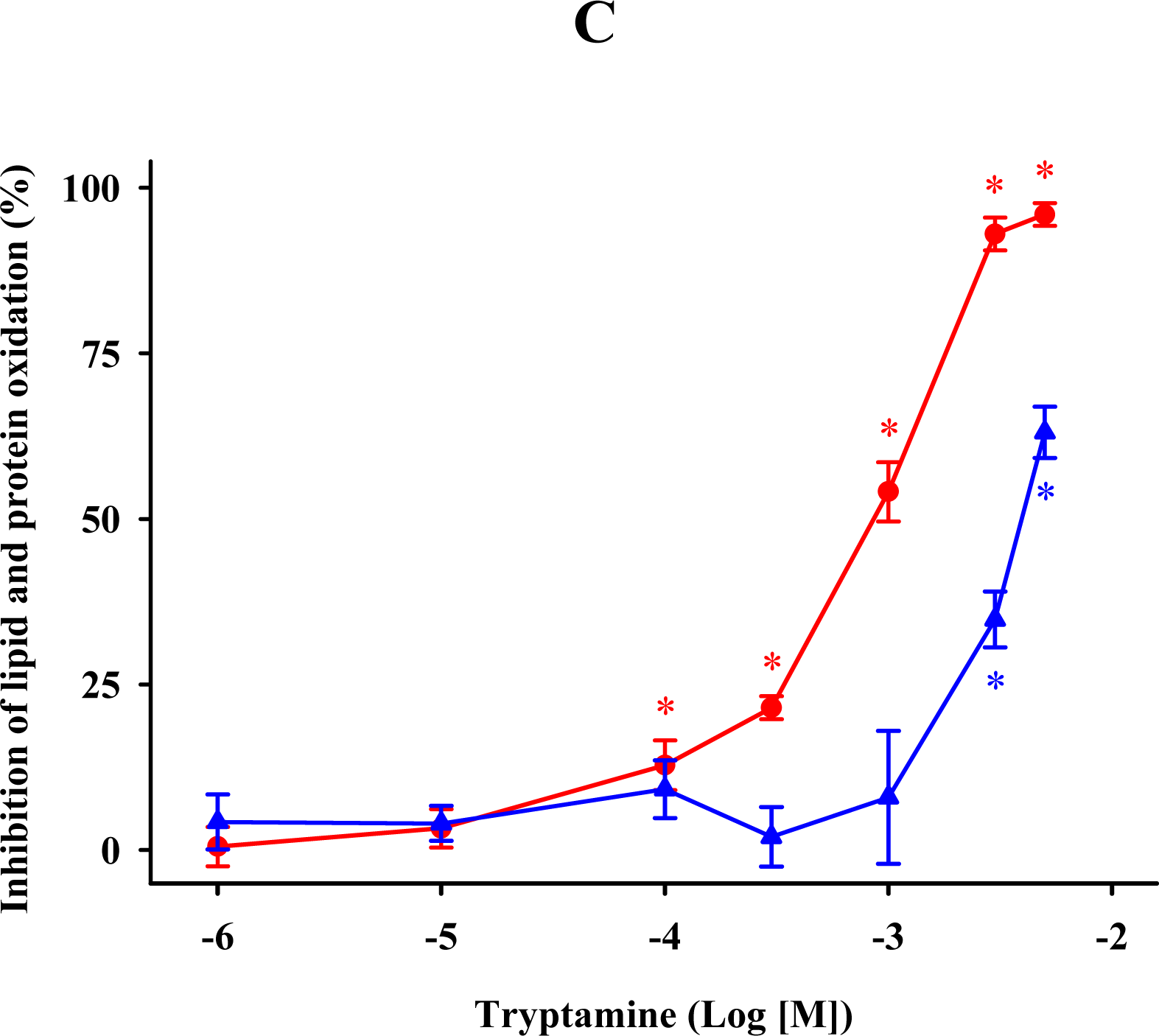 ) MDA + 4-HDA concentrations; ( ) carbonyl contents. Melatonin is expressed as the logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to comparable control membrane preparations. * P < 0.05 vs. membranes treated exclusively with FeCl3 and ascorbic acid.
) MDA + 4-HDA concentrations; ( ) carbonyl contents. Melatonin is expressed as the logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to comparable control membrane preparations. * P < 0.05 vs. membranes treated exclusively with FeCl3 and ascorbic acid.
) MDA + 4-HDA concentrations; ( ) carbonyl contents. Melatonin is expressed as the logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to comparable control membrane preparations. * P < 0.05 vs. membranes treated exclusively with FeCl3 and ascorbic acid.
) MDA + 4-HDA concentrations; ( ) carbonyl contents. Melatonin is expressed as the logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to comparable control membrane preparations. * P < 0.05 vs. membranes treated exclusively with FeCl3 and ascorbic acid.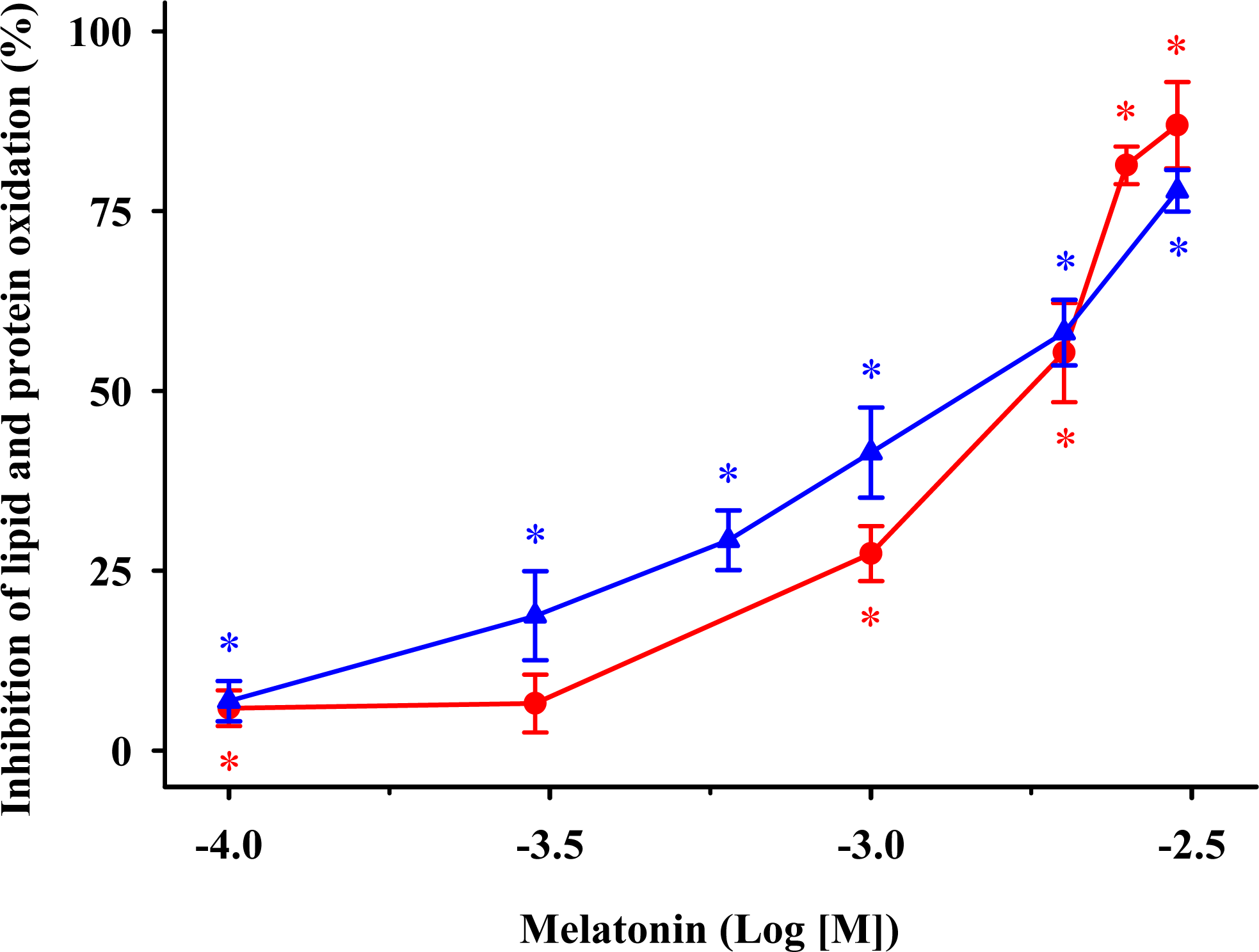 ) and protein ( ) oxidation by 5-methoxytryptophol (A) or 5-methoxy-3-indoleacetic acid (B) induced by FeCl3 and ascorbic acid. The concentrations of these indoleamines are expressed as the decimal logarithm of its molar concentration. Percentage inhibitions are shown as means ± SE (n = 6) and are expressed relative to control membrane preparations. * P < 0.05 vs membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidation by 5-methoxytryptophol (A) or 5-methoxy-3-indoleacetic acid (B) induced by FeCl3 and ascorbic acid. The concentrations of these indoleamines are expressed as the decimal logarithm of its molar concentration. Percentage inhibitions are shown as means ± SE (n = 6) and are expressed relative to control membrane preparations. * P < 0.05 vs membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidation by 5-methoxytryptophol (A) or 5-methoxy-3-indoleacetic acid (B) induced by FeCl3 and ascorbic acid. The concentrations of these indoleamines are expressed as the decimal logarithm of its molar concentration. Percentage inhibitions are shown as means ± SE (n = 6) and are expressed relative to control membrane preparations. * P < 0.05 vs membranes treated only with FeCl3 and ascorbic acid.
) and protein ( ) oxidation by 5-methoxytryptophol (A) or 5-methoxy-3-indoleacetic acid (B) induced by FeCl3 and ascorbic acid. The concentrations of these indoleamines are expressed as the decimal logarithm of its molar concentration. Percentage inhibitions are shown as means ± SE (n = 6) and are expressed relative to control membrane preparations. * P < 0.05 vs membranes treated only with FeCl3 and ascorbic acid.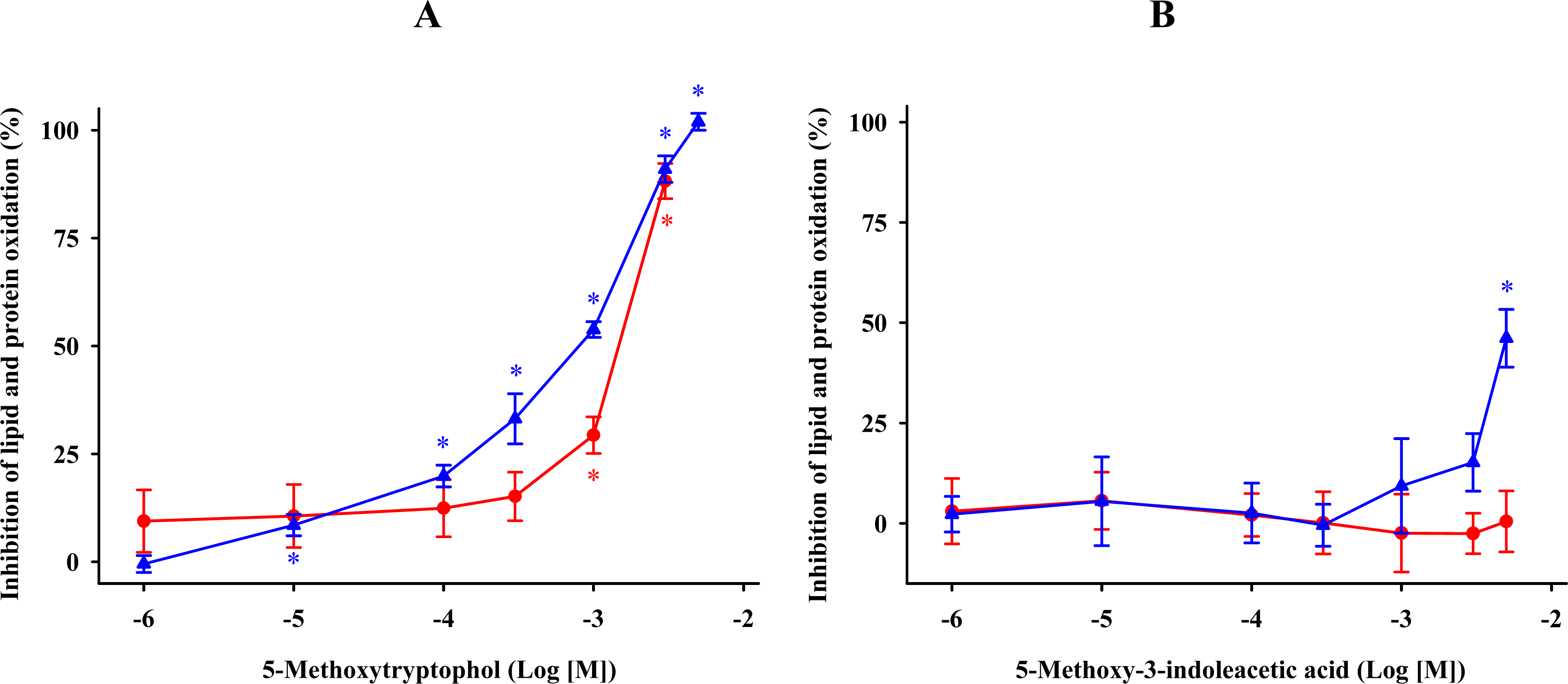 ) and protein ( ) oxidation after oxidative stress. The concentrations of these β-carbolines are expressed as the decimal logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to control synaptosome preparations. * P < 0.05 vs membranes treated exclusively with FeCl3 and ascorbic acid.
) and protein ( ) oxidation after oxidative stress. The concentrations of these β-carbolines are expressed as the decimal logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to control synaptosome preparations. * P < 0.05 vs membranes treated exclusively with FeCl3 and ascorbic acid.
) and protein ( ) oxidation after oxidative stress. The concentrations of these β-carbolines are expressed as the decimal logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to control synaptosome preparations. * P < 0.05 vs membranes treated exclusively with FeCl3 and ascorbic acid.
) and protein ( ) oxidation after oxidative stress. The concentrations of these β-carbolines are expressed as the decimal logarithm of its molar concentration. Values are means ± SE (n = 6) and are expressed relative to control synaptosome preparations. * P < 0.05 vs membranes treated exclusively with FeCl3 and ascorbic acid.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance (n = 6) | Lipid peroxidation | Protein oxidation |
|---|---|---|
| Tryptophan | - | 2.92 |
| 5-Hydroxytryptophan | 3.05 | 1.01 |
| 5-Methoxytryptamine | 0.31 | 3.40 |
| N-acetylserotonin | 0.42 | 0.65 |
| Tryptamine | 0.84 | 3.96 |
| Melatonin | 1.75 | 1.40 |
| 5-Methoxy-3-indoleacetic acid | - | - |
| 5-Methoxytryptophol | 1.75 | 0.76 |
| Tryptoline | 1.49 | 0.63 |
| Pinoline | 0.15 | 0.25 |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Millán-Plano, S.; Piedrafita, E.; Miana-Mena, F.J.; Fuentes-Broto, L.; Martínez-Ballarín, E.; López-Pingarrón, L.; Sáenz, M.A.; García, J.J. Melatonin and Structurally-Related Compounds Protect Synaptosomal Membranes from Free Radical Damage. Int. J. Mol. Sci. 2010, 11, 312-328. https://doi.org/10.3390/ijms11010312
Millán-Plano S, Piedrafita E, Miana-Mena FJ, Fuentes-Broto L, Martínez-Ballarín E, López-Pingarrón L, Sáenz MA, García JJ. Melatonin and Structurally-Related Compounds Protect Synaptosomal Membranes from Free Radical Damage. International Journal of Molecular Sciences. 2010; 11(1):312-328. https://doi.org/10.3390/ijms11010312
Chicago/Turabian StyleMillán-Plano, Sergio, Eduardo Piedrafita, Francisco J. Miana-Mena, Lorena Fuentes-Broto, Enrique Martínez-Ballarín, Laura López-Pingarrón, María A. Sáenz, and Joaquín J. García. 2010. "Melatonin and Structurally-Related Compounds Protect Synaptosomal Membranes from Free Radical Damage" International Journal of Molecular Sciences 11, no. 1: 312-328. https://doi.org/10.3390/ijms11010312