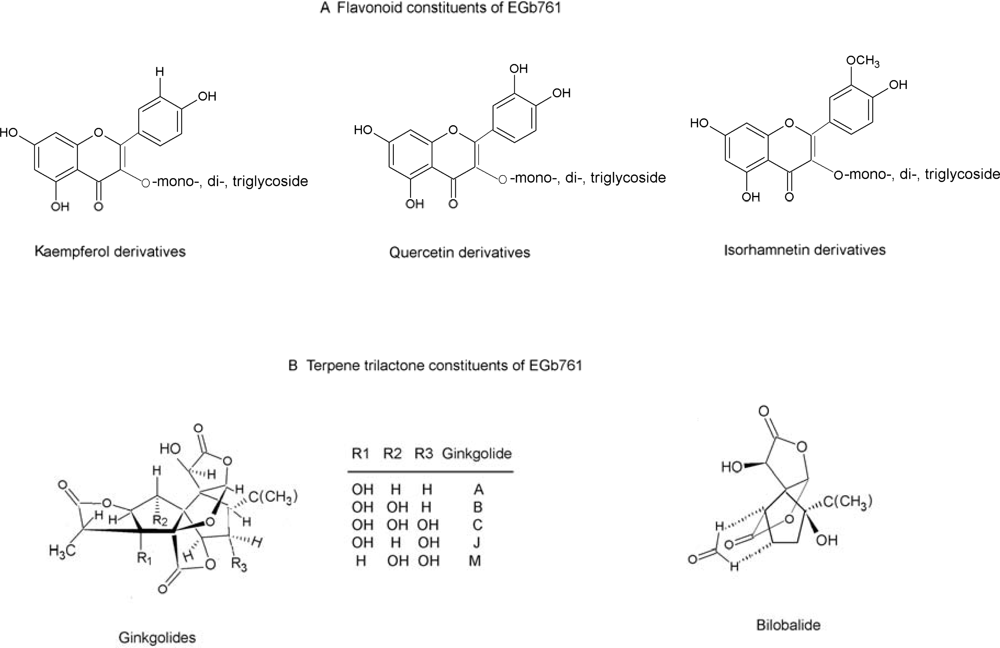
Ginkgo biloba Extract in Alzheimer’s Disease: From Action Mechanisms to Medical Practice

{kind=link}
Abstract
:1. Introduction
2. Mechanism of Neuroprotection by EGb761 and Its Constituents
2.1. Antioxidant Activity
2.2. Protective Effects on Mitochondrial Function
2.3. Anti-Apoptotic Effect
2.4. Anti-Inflammatory Effect
2.5. Protective Effects against Amyloidogenesis and Aβ Aggregation
2.6. Other Mechanisms
3. Current Status of Clinical Use of EGb761: Still a Long Way from Preclinical Promise
4. Conclusions
Acknowledgments
References
- Le Bars, PL. Magnitude of effect and special approach to Ginkgo biloba extract EGb761 in cognitive disorders. Pharmacopsychiatry 2003, 36, S44–S49. [Google Scholar]
- Smith, PF; Maclennan, K; Darlington, CL. The neuroprotective properties of the Ginkgo biloba leaf a review of the possible relationship to platelet-activating factor (PAF). J. Ethnopharmacol 1996, 50, 131–139. [Google Scholar]
- Shi, C; Zhao, L; Zhu, B; Li, Q; Yew, DT; Yao, Z; Xu, J. Protective effects of Ginkgo biloba extract (EGb761) and its constituents quercetin and ginkgolide B against beta-amyloid peptide-induced toxicity in SH-SY5Y cells. Chem. Biol. Interact 2009, 181, 115–123. [Google Scholar]
- Maclennan, KM; Darlington, CL; Smith, PF. The CNS effects of Ginkgo biloba extracts and ginkgolide B. Prog. Neurobiol 2002, 67, 235–257. [Google Scholar]
- Ahlemeyer, B; Krieglstein, J. Neuroprotective effects of Ginkgo biloba extract. Cell Mol. Life Sci 2003, 60, 1779–1792. [Google Scholar]
- Christen, Y. Ginkgo biloba and neurodegenerative disorders. Front Biosci 2004, 9, 3091–3104. [Google Scholar]
- Smith, JV; Luo, Y. Studies on molecular mechanisms of Ginkgo biloba extract. Appl. Microbiol. Biotechnol 2004, 64, 465–472. [Google Scholar]
- Luo, Y. Ginkgo biloba neuroprotection: Therapeutic implications in Alzheimer’s disease. J. Alzheimers Dis 2001, 3, 401–407. [Google Scholar]
- Saleem, S; Zhuang, H; Biswal, S; Christen, Y; Doré, S. Ginkgo biloba extract neuroprotective action is dependent on heme oxygenase 1 in ischemic reperfusion brain injury. Stroke 2008, 39, 3389–3396. [Google Scholar]
- Ponto, LL; Schultz, SK. Ginkgo biloba extract: Review of CNS effects. Ann. Clin. Psychiat 2003, 15, 109–119. [Google Scholar]
- Luo, Y. Alzheimer’s disease, the nematode Caenorhabditis elegans, and ginkgo biloba leaf extract. Life Sci 2006, 78, 2066–2072. [Google Scholar]
- Holstein, N. Ginkgo special extract EGb 761 in tinnitus therapy. An overview of results of completed clinical trials. Fortschr. Med. Orig 2001, 118, 157–164. [Google Scholar]
- Schneider, B. Ginkgo biloba extract in peripheral arterial diseases: Meta-analysis of controlled clinical studies. Arzneimittelforschung 1992, 42, 428–436. [Google Scholar]
- Shi, C; Xu, XW; Forster, EL; Tang, LF; Ge, Z; Yew, DT; Xu, J. Possible role of mitochondrial dysfunction in central neurodegeneration of ovariectomized rats. Cell Biochem. Funct 2008, 26, 172–178. [Google Scholar]
- Castellani, RJ; Lee, HG; Perry, G; Smith, MA. Antioxidant protection and neurodegenerative disease: The role of amyloid-beta and tau. Am. J. Alzheimers Dis. Other Demen 2006, 21, 126–130. [Google Scholar]
- García-Ayllón, MS; Silveyra, MX; Sáez-Valero, J. Association between acetylcholinesterase and beta-amyloid peptide in Alzheimer’s cerebrospinal fluid. Chem. Biol. Interact 2008, 175, 209–215. [Google Scholar]
- Dinamarca, MC; Arrázola, M; Toledo, E; Cerpa, WF; Hancke, J; Inestrosa, NC. Release of acetylcholinesterase (AChE) from beta-amyloid plaques assemblies improves the spatial memory impairments in APP-transgenic mice. Chem. Biol. Interact 2008, 175, 142–149. [Google Scholar]
- Gella, A; Durany, N. Oxidative stress in Alzheimer disease. Cell Adh. Migr 2009, 3, 88–93. [Google Scholar]
- Cameron, B; Landreth, GE. Inflammation, microglia, and alzheimer’s disease. Neurobiol Dis 2009. [Google Scholar]
- Magaki, S; Mueller, C; Dickson, C; Kirsch, W. Increased production of inflammatory cytokines in mild cognitive impairment. Exp. Gerontol 2007, 42, 233–240. [Google Scholar]
- Culpan, D; Cornish, A; Love, S; Kehoe, PG; Wilcock, GK. Protein and gene expression of tumour necrosis factor receptors I and II and their promoter gene polymorphisms in Alzheimer’s disease. Exp. Gerontol 2007, 42, 538–544. [Google Scholar]
- Koutsilieri, E; Riederer, P. Excitotoxicity and new antiglutamatergic strategies in Parkinson’s disease and Alzheimer’s disease. Parkinsonism Relat. Disord 2007, 13, S329–S331. [Google Scholar]
- Bertoni-Freddari, C; Fattoretti, P; Casoli, T; di Stefano, G; Balietti, M; Giorgetti, B; Perretta, G. Neuronal apoptosis in Alzheimer’s disease: The role of age-related mitochondrial metabolic competence. Ann. N. Y. Acad. Sci 2009, 1171, 18–24. [Google Scholar]
- Klegeris, A; McGeer, PL. Non-steroidal anti-inflammatory drugs (NSAIDs) and other anti-inflammatory agents in the treatment of neurodegenerative disease. Curr. Alzheimer Res 2005, 2, 355–365. [Google Scholar]
- Zhang, HY; Zheng, CY; Yan, H; Wang, ZF; Tang, LL; Gao, X; Tang, XC. Potential therapeutic targets of huperzine A for Alzheimer’s disease and vascular dementia. Chem. Biol. Interact 2008, 175, 396–402. [Google Scholar]
- Akasofu, S; Kimura, M; Kosasa, T; Sawada, K; Ogura, H. Study of neuroprotection of donepezil, a therapy for Alzheimer’s disease. Chem. Biol. Interact 2008, 175, 222–226. [Google Scholar]
- Cole, GM; Frautschy, SA. The role of insulin and neurotrophic factor signaling in brain aging and Alzheimer’s Disease. Exp. Gerontol 2007, 42, 10–21. [Google Scholar]
- Steele, M; Stuchbury, G; Münch, G. The molecular basis of the prevention of Alzheimer’s disease through healthy nutrition. Exp. Gerontol 2007, 42, 28–36. [Google Scholar]
- Kovács, T. Therapy of Alzheimer disease. Neuropsychopharmacol. Hung 2009, 11, 27–33. [Google Scholar]
- Zimmermann, M; Colciaghi, F; Cattabeni, F; di Luca, M. Ginkgo biloba extract: From molecular mechanisms to the treatment of Alzhelmer’s disease. Cell Mol. Biol (Noisy-le-grand) 2002, 48, 613–623. [Google Scholar]
- Wei, T; Ni, Y; Hou, J; Chen, C; Zhao, B; Xin, W. Hydrogen peroxide-induced oxidative damage and apoptosis in cerebellar granule cells: Protection by Ginkgo biloba extract. Pharmacol. Res 2000, 41, 427–433. [Google Scholar]
- Wu, Y; Wu, Z; Butko, P; Christen, Y; Lambert, MP; Klein, WL; Link, CD; Luo, Y. Amyloid-beta-induced pathological behaviors are suppressed by Ginkgo biloba extract EGb 761 and ginkgolides in transgenic Caenorhabditis elegans. J. Neurosci 2006, 26, 13102–13113. [Google Scholar]
- Bridi, R; Crosetti, FP; Steffen, VM; Henriques, AT. The antioxidant activity of standardized extract of Gingko biloba (EGb 761) in rats. Phytother. Res 2001, 15, 449–451. [Google Scholar]
- Colak, Ö; Sahin, A; Alatas, Ö; Inal, M; Yasar, B; Kiper, H. The effect of Ginkgo biloba on the activity of catalase and lipid peroxidation in experimental strangulation ileus. Int. J. Clin. Lab. Res 1998, 28, 69–71. [Google Scholar]
- Sasaki, K; Hatta, S; Wada, K; Ueda, N; Yoshimura, T; Endo, T; Sakata, M; Tanaka, T; Haga, M. Effects of extract of Ginkgo biloba leaves and its constituents on carcinogen-metabolizing enzyme activities and glutathione levels in mouse liver. Life Sci 2002, 70, 1657–1667. [Google Scholar]
- Rimbach, G; Gohil, K; Matsugo, S; Moini, H; Saliou, C; Virgili, F; Weber, SU; Packer, L. Induction of glutathione synthesis in human keratinocytes by Ginkgo biloba extract (EGb 761). Biofactors 2001, 15, 39–52. [Google Scholar]
- Smith, J; Luo, Y. Elevation of oxidative free radicals in Alzheimer’s disease models can be attenuated by Ginkgo biloba extract EGb 761. J. Alzheimers Dis 2003, 5, 287–300. [Google Scholar]
- Maitra, I; Marcocci, L; Droy-Lefaix, MT; Packer, L. Peroxyl radical scavenging activity of Ginkgo biloba extract EGb 761. Biochem. Pharmacol 1995, 45, 1649–1655. [Google Scholar]
- Gohil, K; Packer, L. Global gene expression analysis identifies cell and tissue specific actions of Ginkgo biloba extract, EGb 761. Cell Mol. Biol 2002, 48, 625–631. [Google Scholar]
- Oken, B; Storzbach, D; Kaye, J. The efficacy of Ginkgo biloba on cognitive function in Alzheimer disease. Arch. Neurol 1998, 55, 1409–1415. [Google Scholar]
- Saija, A; Scalese, M; Lanza, M; Marzullo, D; Bonina, F; Castelli, F. Flavonoids as antioxidant agents: Importance of their interaction with biomembranes. Free Radic. Biol. Med 1995, 19, 481–486. [Google Scholar]
- Sestili, P; Guidarelli, A; Dacha, M; Cantoni, O. Quercetin prevents DNA single strand breakage and cytotoxicity caused by tert-butylhydroperoxide: Free radical scavenging versus iron chelating mechanism. Free Radic. Biol. Med 1998, 25, 196–200. [Google Scholar]
- Scholtyssek, H; Damerau, W; Wessel, R; Schimke, I. Antioxidative activity of ginkgolides against superoxide in an aprotic environment. Chem. Biol. Interact 1997, 106, 183–190. [Google Scholar]
- Pietri, S; Maurelli, E; Drieu, K; Culcasi, M. Cardioprotective and anti-oxidant effects of the terpenoid constituents of Ginkgo biloba extract (EGb 761). J. Mol. Cell. Cardiol 1997, 29, 733–742. [Google Scholar]
- Castellani, R; Hirai, K; Aliev, G; Drew, KL; Nunomura, A; Takeda, A; Cash, AD; Obrenovich, ME; Perry, G; Smith, MA. Role of mitochondrial dysfunction in Alzheimer’s disease. J. Neurosci. Res 2002, 70, 357–360. [Google Scholar]
- Tendi, EA; Bosetti, F; Dasgupta, SF; Stella, AM; Drieu, K; Rapoport, SI. Ginkgo biloba extracts (EGb 761) and bilobalide increase NADH dehydrogenase mRNA level and mitochondrial respiratory control ratio in PC12 cells. Neurochem. Res 2002, 27, 319–323. [Google Scholar]
- Shi, C; Fang, L; Yew, DT; Yao, Z; Xu, J. Ginkgo Biloba extract EGb761 protects against mitochondrial dysfunction in platelets and hippocampi in ovariectomized rats. Platelets 2009. [Google Scholar]
- Shi, C; Liu, J; Guo, KH; Wu, FM; Yew, DT; Yao, ZB; Xu, J. Ginkgo biloba extract EGb761 protects against aging-associated mitochondrial dysfunction in platelets and hippocampi of SAMP8 mice. Platelets 2009. [Google Scholar]
- Yuan, J; Yankner, BA. Apoptosis in the nervous system. Nature 2000, 407, 802–809. [Google Scholar]
- Shi, C; Zhao, L; Zhu, B; Li, Q; Yew, DT; Yao, Z; Xu, J. Dosage effects of EGb761 on hydrogen peroxide-induced cell death in SH-SY5Y cells. Chem. Biol. Interact 2009, 18, 389–397. [Google Scholar]
- Wang, L; Matsushita, K; Araki, I; Takeda, M. Inhibition of c-Jun N-terminal kinase ameliorates apoptosis induced by hydrogen peroxide in the kidney tubule epithelial cells (NRK-52E). Nephron 2002, 91, 142–147. [Google Scholar]
- Uchida, K; Shiraishi, M; Naito, Y; Torii, Y; Nakamura, Y; Osawa, T. Activation of stress signaling pathways by the end product of lipid peroxidation 4-hydroxy-2-nonenal is a potential inducer of intracellular peroxide production. J. Biol. Chem 1999, 274, 2234–2242. [Google Scholar]
- Spencer, JP; Rice-Evans, C; Williams, RJ. Modulation of Pro-survival Akt/Protein Kinase B and ERK1/2 Signaling Cascades by Quercetin and Its in vivo Metabolites Underlie Their Action on Neuronal Viability. J. Biol. Chem 2003, 278, 34783–34793. [Google Scholar]
- Ishikawa, Y; Kitamura, M. Anti-apoptotic effect of quercetin: Intervention in the JNK-and ERK-mediated apoptotic pathways. Kidney Int 2000, 58, 1078–1087. [Google Scholar]
- Kong, AN; Yu, R; Chen, C; Mandlekar, S; Primiano, T. Signal transduction events elicited by natural products: Role of MAPK and caspase pathways in homeostatic response and induction of apoptosis. Arch. Pharm. Res 2000, 23, 1–16. [Google Scholar]
- Ahlemeyer, B; Möwes, A; Krieglstein, J. Inhibition of serum deprivationand staurosporine-induced neuronal apoptosis by Ginkgo biloba extract and some of its constituents. Eur. J. Pharmacol 1999, 367, 423–430. [Google Scholar]
- Defeudis, FV. Bilobalide and neuroprotection. Pharmacol. Res 2002, 46, 565–568. [Google Scholar]
- Ahlemeyer, B; Krieglstein, J. Pharmacological studies supporting the therapeutic use of Ginkgo biloba extract for Alzheimer’s disease. Pharmacopsychiatry 2003, 36, S8–S14. [Google Scholar]
- Chan, WH; Hsuuw, YD. Dosage effects of ginkgolide B on ethanol-induced cell death in human hepatoma G2 cells. Ann. N. Y. Acad. Sci 2007, 1095, 388–398. [Google Scholar]
- Rapin, JR; Zaibi, M; Drieu, K. In vitro and in vivo effects of an extract of Ginkgo biloba (EGb 761), ginkgolide B, and bilobalide on apoptosis in primary cultures of rat hippocampal neurons. Drug Dev. Res 1998, 45, 23–29. [Google Scholar]
- Xin, W; Wei, T; Chen, C; Ni, Y; Zhao, B; Hou, J. Mechanisms of apoptosis in rat cerebellar granule cells induced by hydroxyl radicals and the effects of EGb761 and its constituents. Toxicology 2000, 148, 103–110. [Google Scholar]
- de Toledo, M. Inflammation and Alzheimer’s disease. Rev. Neurol 2006, 42, 433–438. [Google Scholar]
- Braquet, P. The ginkgolides: Potent platelet-activating factor antagonists isolated from ginkgo-biloba L.:Chemistry, pharmacology and clinical applications. Drugs Future 1987, 12, 643–688. [Google Scholar]
- Braquet, P; Hosford, D. Ethnopharmacology and the development of natural PAF antagonists as therapeutic agents. J. Ethnopharmacol 1991, 32, 135–139. [Google Scholar]
- Chan, PC; Xia, Q; Fu, PP. Ginkgo biloba leave extract: Biological, medicinal, and toxicological effects. J. Environ. Sci. Health C. Environ. Carcinog. Ecotoxicol. Rev 2007, 25, 211–244. [Google Scholar]
- Bonavida, B; Mencia-Huerta, JM. Platelet-activating factor and the cytokine network in inflammatory processes. Clin. Rev. Allergy 1994, 12, 381–395. [Google Scholar]
- Hynes, N; Bishai, I; Lees, J; Coceani, F. Leukotrienes in brain: Natural occurrence and induced changes. Brain Res 1991, 553, 4–13. [Google Scholar]
- Aihara, M; Ishii, S; Kume, K; Shimizu, T. Interaction between neurone and microglia mediated by platelet-activating factor. Genes Cells 2000, 5, 397–406. [Google Scholar]
- Farooqui, AA; Horrocks, LA. Phospholipase A2-Generated Lipid Mediators in the Brain: The Good, the Bad, and the Ugly. The Neuroscientist 2006, 12, 245–260. [Google Scholar]
- Bate, C; Tayebi, M; Williams, A. Ginkgolides protect against amyloid-beta1-42-mediated synapse damage in vitro. Mol. Neurodegener 2008, 3, 1. [Google Scholar]
- Kunievsky, B; Yavin, E. Platelet-activating factor stimulates arachidonic acid release and enhances thromboxane B2 production in intact fetal rat brain ex vivo. J. Pharmacol. Exp. Ther 1992, 263, 562–568. [Google Scholar]
- Du, ZY; Li, XY. Effects of ginkgolides on interleukin-1, tumor necrosis factor-alpha and nitric oxide production by rat microglia stimulated with lipopolysaccharidesin vitro. Arzneim.—Forsch 1998, 48, 1126–1130. [Google Scholar]
- Gerlai, R. Alzheimer’s disease: Beta-amyloid hypothesis strengthened! Trends Neurosci 2001, 24, 199. [Google Scholar]
- Bastianetto, S; Ramassamy, C; Doré, S; Christen, S; Poirier, J; Quirion, R. The Ginkgo biloba extract (EGb 761) protects hippocampal neurons against cell death induced by beta-amyloid. Eur. J. Neurosci 2000, 12, 1882–1890. [Google Scholar]
- Smith, JV; Luo, Y. Elevation of oxidative free radicals in Alzheimer’s disease models can be attenuated by Ginkgo biloba extract EGb 761. J. Alzheimers Dis 2003, 5, 287–300. [Google Scholar]
- Colciaghi, F; Borroni, B; Zimmermann, M; Bellone, C; Longhi, A; Padovani, A; Cattabeni, F; Christen, Y; Di Luca, M. Amyloid precursor protein metabolism is regulated toward alpha-secretase pathway by Ginkgo biloba extracts. Neurobiol. Dis 2004, 16, 454–460. [Google Scholar]
- Yao, ZX; Han, Z; Drieu, K; Papadopoulos, V. Ginkgo biloba extract (EGb 761) inhibits beta-amyloid production by lowering free cholesterol levels. J. Nutr. Biochem 2004, 15, 749–756. [Google Scholar]
- Ramassamy, C. Emerging role of polyphenolic compounds in the treatment of neurodegenerative diseases: A review of their intracellular targets. Eur. J. Pharmacol 2006, 545, 51–64. [Google Scholar]
- Stackman, RW; Eckenstein, F; Frei, B; Kulhanek, D; Nowlin, J; Quinn, JF. Prevention of age-related spatial memory deficits in a transgenic mouse model of Alzheimer’s disease by chronic Ginkgo biloba treatment. Exp Neurol 2003, 184, 510–520. [Google Scholar]
- Bodovitz, S; Klein, WL. Cholesterol modulates alpha-secretase cleavage of amyloid precursor protein. J. Biol. Chem 1996, 271, 4436–4440. [Google Scholar]
- Howland, DS; Trusko, SP; Savage, MJ; Reaume, AG; Lang, DM; Hirsch, JD; Maeda, N; Siman, R; Greenberg, BD; Scott, RW; Flood, DG. Modulation of secreted beta-amyloid precursor protein and amyloid beta-peptide in brain by cholesterol. J. Biol. Chem 1998, 273, 16576–16582. [Google Scholar]
- Simons, M; Keller, P; de Strooper, B; Beyreuther, K; Dotti, CG; Simons, K. Cholesterol depletion inhibits the generation of beta-amyloid in hippocampal neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 6460–6464. [Google Scholar]
- Pike, CJ; Walencewicz, AJ; Glabe, CG; Cotman, CW. In vitro aging of beta-amyloid protein causes peptide aggregation and neurotoxicity. Brain Res 1991, 563, 311–314. [Google Scholar]
- Simmons, LK; May, PC; Tomaselli, KJ; Rydel, RE; Fuson, KS; Brigham, EF; Wright, S; Lieberburg, I; Becker, GW; Brems, DN. Secondary structure of amyloid beta peptide correlates with neurotoxic activity in vitro. Mol. Pharmacol 1994, 45, 373–379. [Google Scholar]
- Soto, C; Sigurdsson, EM; Morelli, L; Kumar, RA; Castano, EM; Frangione, B. Beta-sheet breaker peptides inhibit fibrillogenesis in a rat brain model of amyloidosis: Implications for Alzheimer’s therapy. Nat. Med 1998, 4, 822–826. [Google Scholar]
- Luo, Y; Smith, JV; Paramasivam, V; Burdick, A; Curry, KJ; Buford, JP; Khan, I; Netzer, WJ; Xu, H; Butko, P. Inhibition of amyloid-beta aggregation and caspase-3 activation by the Ginkgo biloba extract EGb761. Proc. Natl. Acad. Sci. USA 2002, 99, 12197–12202. [Google Scholar]
- Watanabe, CM; Wolffram, S; Ader, P; Rimbach, G; Packer, L; Maguire, JJ; Schultz, PG; Gohil, K. The in vivo neuromodulatory effects of the herbal medicine ginkgo biloba. Proc. Natl. Acad. Sci. USA 2001, 98, 6577–6580. [Google Scholar]
- Tsuzuki, K; Yamaguchi, H; Tateno, M; Imai, K; Fujii, N; Yamauchi, T. Transthyretin binds amyloid beta peptides, Abeta1-42 and Abeta1-40 to form complex in the autopsied human kidney -possible role of transthyretin for abeta sequestration. Neurosci. Lett 2000, 281, 171–174. [Google Scholar]
- Berrocal, M; Marcos, D; Sepúlveda, MR; Pérez, M; Avila, J; Mata, AM. Altered Ca2+ dependence of synaptosomal plasma membrane Ca2+-ATPase in human brain affected by Alzheimer’s disease. FASEB J 2009, 23, 1826–1834. [Google Scholar]
- Shirwany, NA; Payette, D; Xie, J; Guo, Q. The amyloid beta ion channel hypothesis of Alzheimer’s disease. Neuropsychiatr. Dis. Treat 2007, 3, 597–612. [Google Scholar]
- Liu, F; Zaidi, T; Iqbal, K; Grundke-Iqbal, I; Merkle, RK; Gong, CX. Role of glycosylation in hyperphosphorylation of tau in Alzheimer’s disease. FEBS Lett 2002, 512, 101–106. [Google Scholar]
- Scott, SA; Mufson, EJ; Weingartner, JA; Skau, KA; Crutcher, KA. Nerve growth factor in Alzheimer’s disease: Increased levels throughout the brain coupled with declines in nucleus basalis. J. Neurosci 1995, 15, 6213–6221. [Google Scholar]
- Peng, S; Garzon, DJ; Marchese, M; Klein, W; Ginsberg, SD; Francis, BM; Mount, HT; Mufson, EJ; Salehi, A; Fahnestock, M. Decreased brain-derived neurotrophic factor depends on amyloid aggregation state in transgenic mouse models of Alzheimer’s disease. J. Neurosci 2009, 29, 9321–9329. [Google Scholar]
- Pierre, S; Jamme, I; Droy-Lefaix, MT; Nouvelot, A; Maixent, JM. Ginkgo biloba extract (EGb 761) protects Na,K-ATPase activity during cerebral ischemia. Neuroreport 1999, 10, 47–51. [Google Scholar]
- Zheng, SX; Zhou, LJ; Chen, ZL; Yin, ML; Zhu, XZ. Bilobalide promotes expression of glial cell line-derived growth neurotrophic factor and vascular endothelial growth factor in rat astrocytes. Acta. Pharmacol. Sin 2000, 21, 151–155. [Google Scholar]
- Birks, J; Grimley Evans, J. Ginkgo biloba for cognitive impairment and dementia. Cochrane Database Syst Rev 2009, 1, D003120. [Google Scholar]
- DeKosky, ST; Williamson, JD; Fitzpatrick, AL; Kronmal, RA; Ives, DG; Saxton, JA; Lopez, OL; Burke, G; Carlson, MC; Fried, LP; Kuller, LH; Robbins, JA; Tracy, RP; Woolard, NF; Dunn, L; Snitz, BE; Nahin, RL; Furberg, CD. Ginkgo Evaluation of Memory (GEM) Study Investigators, Ginkgo biloba for prevention of dementia: A randomized controlled trial. JAMA 2008, 300, 2253–2262. [Google Scholar]
- Kanowski, S; Herrmann, WM; Stephan, K; Wierich, W; Hörr, R. Proof of efficacy of the ginkgo biloba special extract EGb 761 in outpatients suffering from mild to moderate primary degenerative dementia of the Alzheimer type or multi-infarct dementia. Pharmacopsychiatry 1996, 29, 47–56. [Google Scholar]
- Hofferberth, B. The efficacy of EGb 761 in patients with senile dementia of the Alzheimer type. A double-blind, placebo controlled study on different levels of investigation. Hum. Psychopharmacol 1994, 9, 215–222. [Google Scholar]
- Shi, C; Yao, Z; Xu, J; Yew, DT. Effects of Gingko Extract (EGb761) on oxidative damage under different conditions of serum supply. J. Bioenerg. Biomembr 2009, 41, 61–69. [Google Scholar]
- Biber, A. Pharmacokinetics of Ginkgo biloba extracts. Pharmacopsychiatry 2003, 36, S32–S37. [Google Scholar]
- Amri, H; Drieu, K; Papadopoulos, V. Transcriptional suppression of the adrenal cortical peripheral-type benzodiazepine receptor gene and inhibition of steroid synthesis by ginkgolide B. Biochem. Pharmacol 2003, 65, 717–729. [Google Scholar]
- Farrall, AJ; Wardlaw, JM. Blood–brain barrier: Ageing and microvascular disease—systematic review and meta-analysis. Neurobiol. Aging 2009, 30, 337–352. [Google Scholar]
- Popescu, BO; Toescu, EC; Popescu, LM; Bajenaru, O; Muresanu, DF; Schultzberg, M; Bogdanovic, N. Blood–brain barrier alterations in ageing and dementia. J. Neurol. Sci 2009, 283, 99–106. [Google Scholar]
- Kanowski, S; Herrmann, WM; Stephan, K; Wierich, W; Herr, R. Proof of efficacy of the Ginkgo biloba special extract EGB 761 in outpatients suffering from mild to moderate primary degenerative dementia of the Alzheimer type or multi-infarct dementia. Phannacopsychiatry 1996, 29, 47–56. [Google Scholar]
- Garcia-Alloza, M; Dodwell, SA; Meyer-Luehmann, M; Hyman, BT; Bacskai, BJ. Plaque-derived oxidative stress mediates distorted neurite trajectories in the Alzheimer mouse model. J. Neuropathol. Exp. Neurol 2006, 65, 1082–1089. [Google Scholar]
- D’Aurelio, M; Merlo Pich, M; Catani, L; Sgarbi, GL; Bovina, C; Formiggini, G; Parenti Castelli, G; Baum, H; Tura, S; Lenaz, G. Decreased Pasteur effect in platelets of aged individuals. Mech. Ageing Dev 2001, 122, 823–833. [Google Scholar]
- Henderson, VW; Buckwalter, JG. Cognitive deficits of men and women with Alzheimer disease. Neurology 1994, 44, 90–96. [Google Scholar]
- Wilson, AC; Clemente, L; Liu, T; Bowen, RL; Meethal, SV; Atwood, CS. Reproductive hormones regulate the selective permeability of the blood-brain barrier. Biochim. Biophys. Acta 2008, 1782, 401–417. [Google Scholar]
- Cipolla, MJ; Godfrey, JA; Wiegman, MJ. The effect of ovariectomy and estrogen on penetrating brain arterioles and blood-brain barrier permeability. Microcirculation 2009, 9, 1–10. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shi, C.; Liu, J.; Wu, F.; Yew, D.T. Ginkgo biloba Extract in Alzheimer’s Disease: From Action Mechanisms to Medical Practice. Int. J. Mol. Sci. 2010, 11, 107-123. https://doi.org/10.3390/ijms11010107
Shi C, Liu J, Wu F, Yew DT. Ginkgo biloba Extract in Alzheimer’s Disease: From Action Mechanisms to Medical Practice. International Journal of Molecular Sciences. 2010; 11(1):107-123. https://doi.org/10.3390/ijms11010107
Chicago/Turabian StyleShi, Chun, Jun Liu, Fengming Wu, and David T. Yew. 2010. "Ginkgo biloba Extract in Alzheimer’s Disease: From Action Mechanisms to Medical Practice" International Journal of Molecular Sciences 11, no. 1: 107-123. https://doi.org/10.3390/ijms11010107