Overexpression of a Weed (Solanum americanum) Proteinase Inhibitor in Transgenic Tobacco Results in Increased Glandular Trichome Density and Enhanced Resistance to Helicoverpa armigera and Spodoptera litura

Abstract
:
1. Introduction
2. Results and Discussion
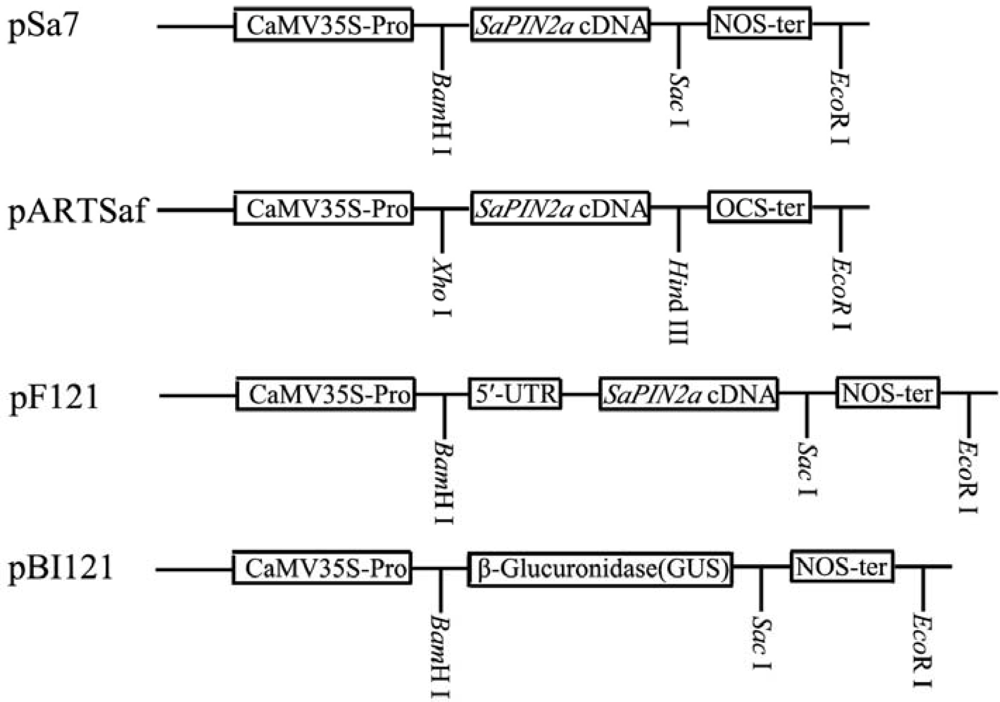
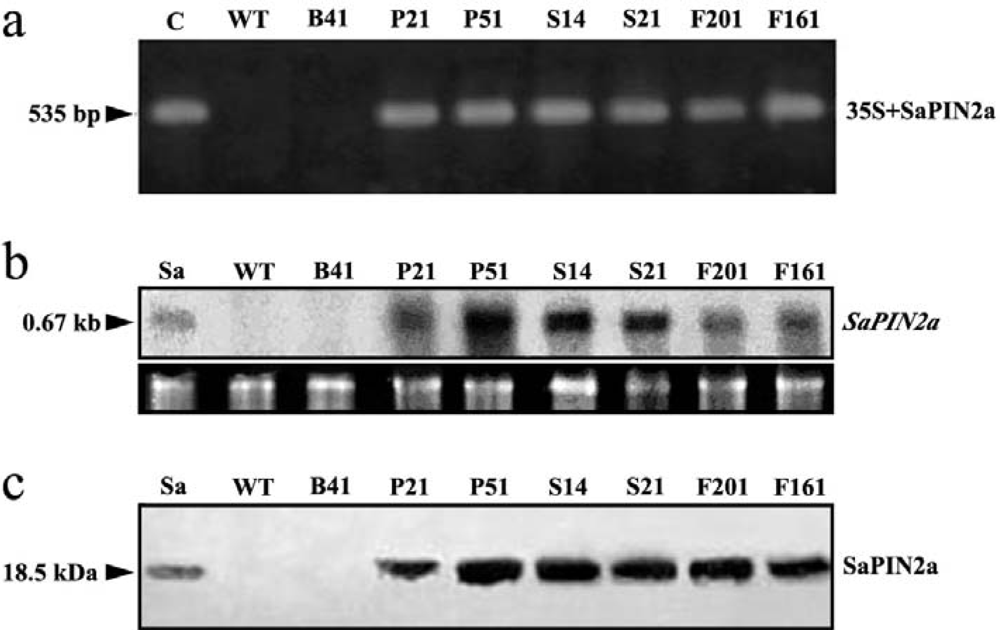
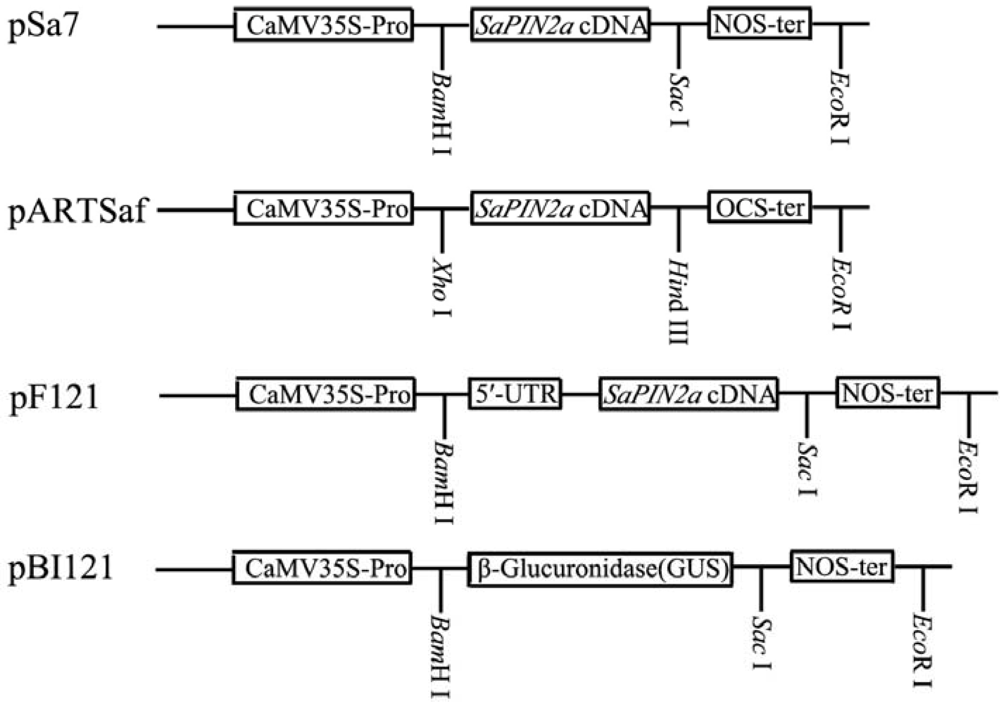
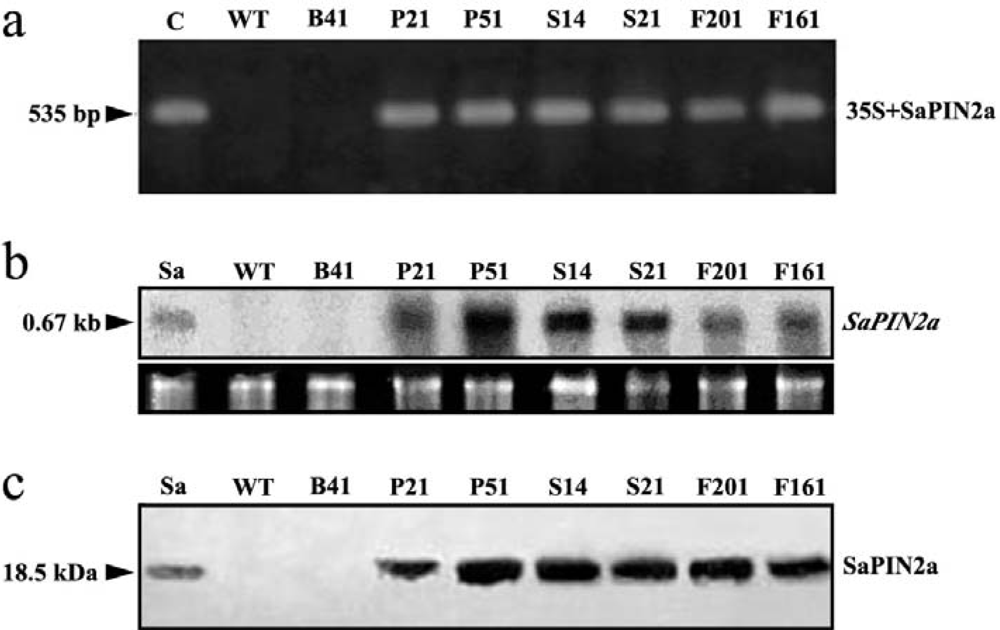
2.1. Expression of SaPIN2a in transgenic tobacco plants
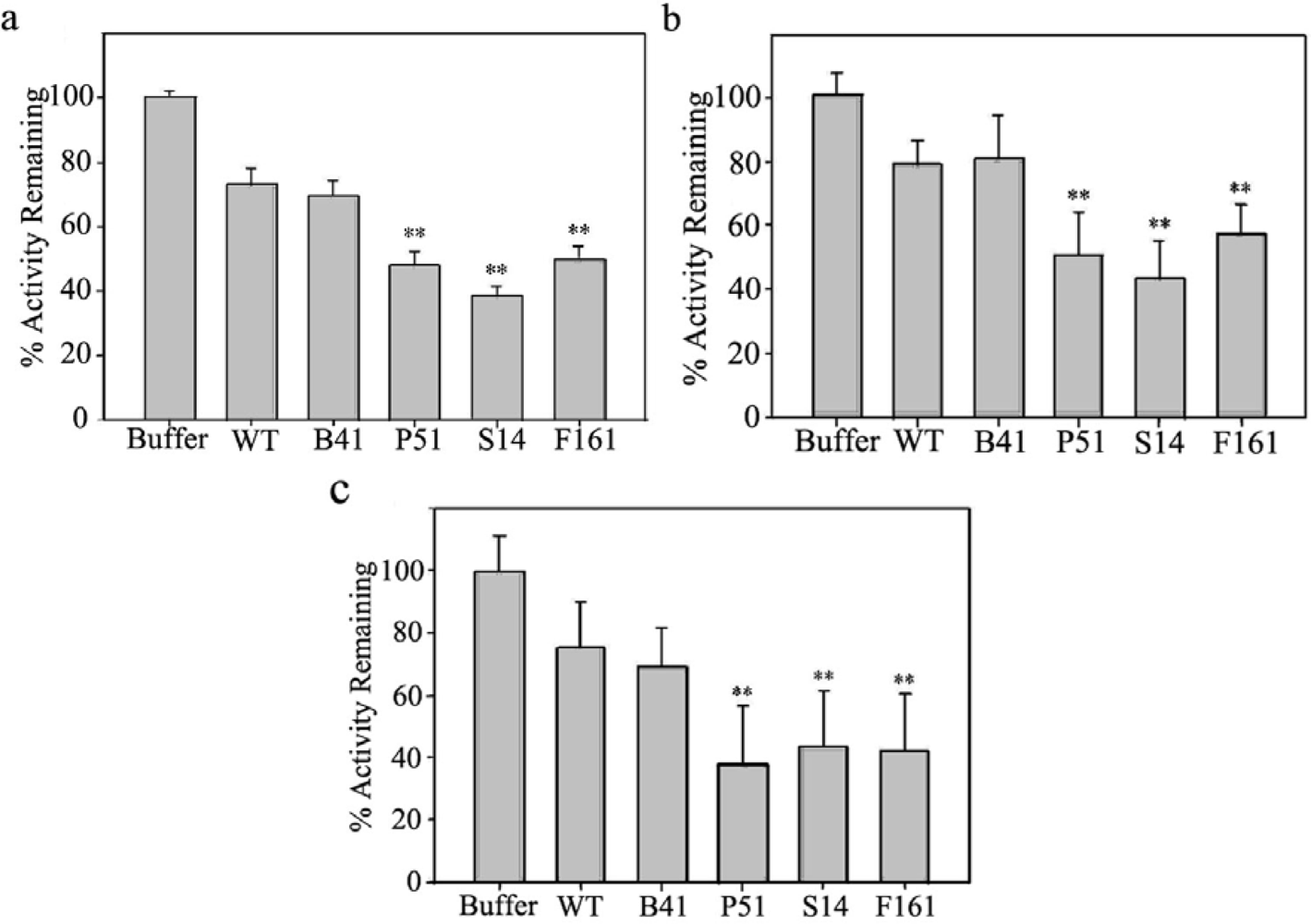
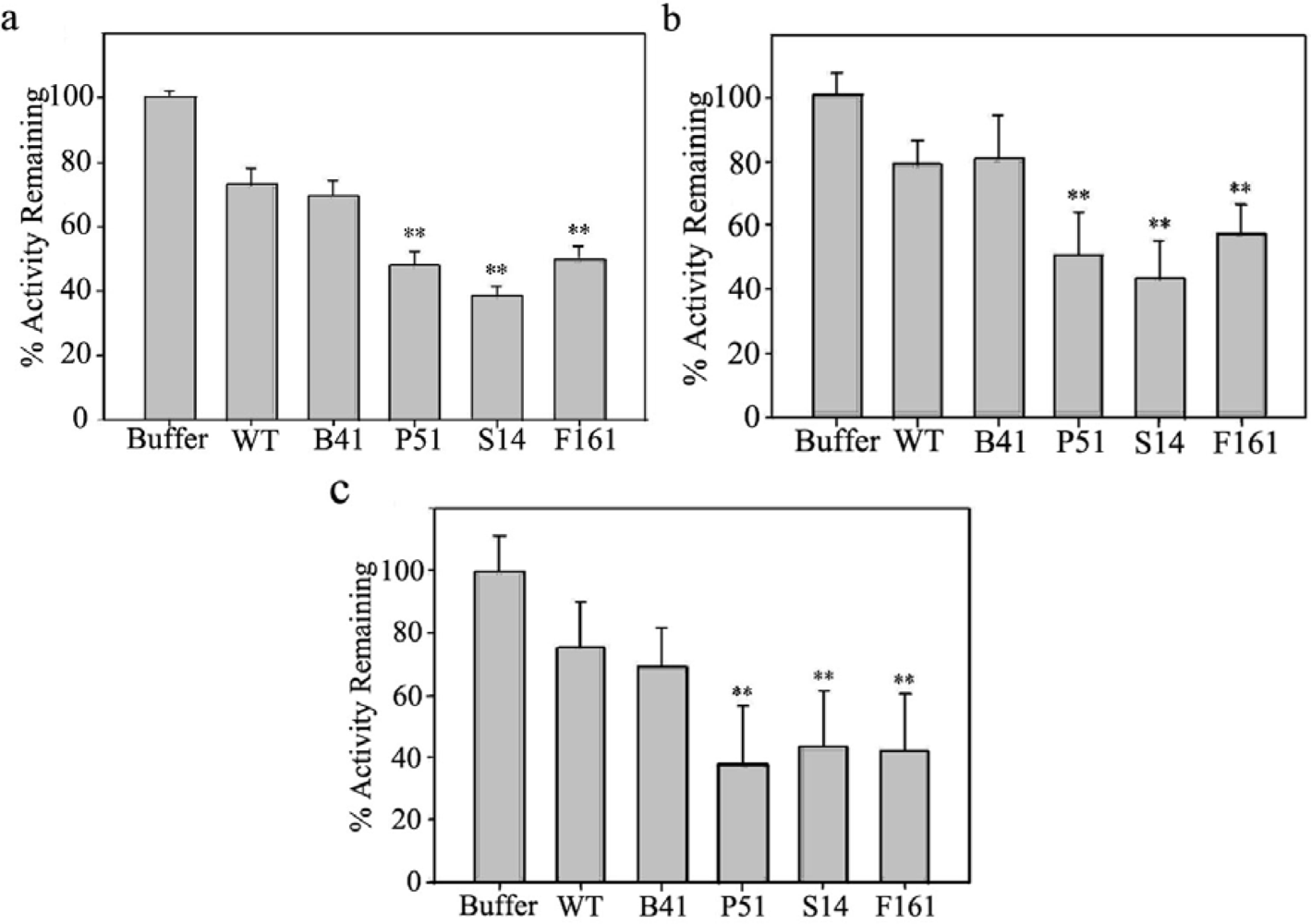
2.2. Proteinase inhibitory activities of transgenic SaPIN2a
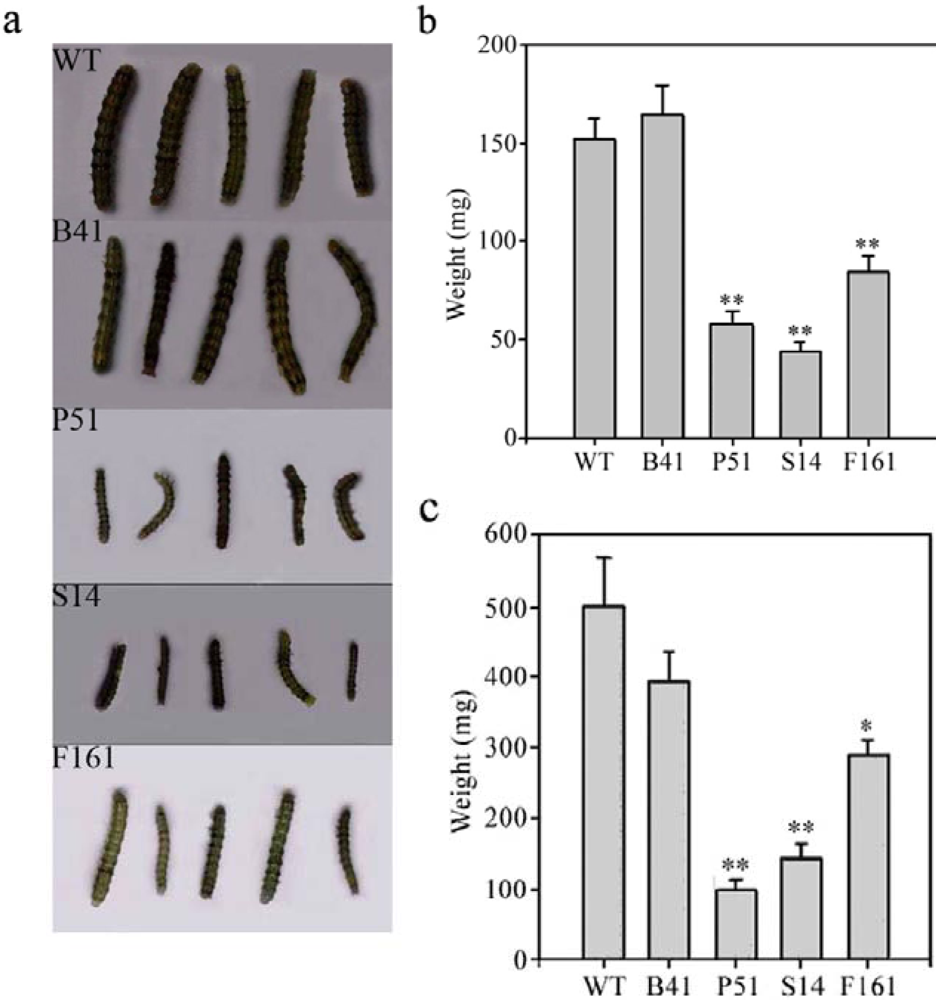
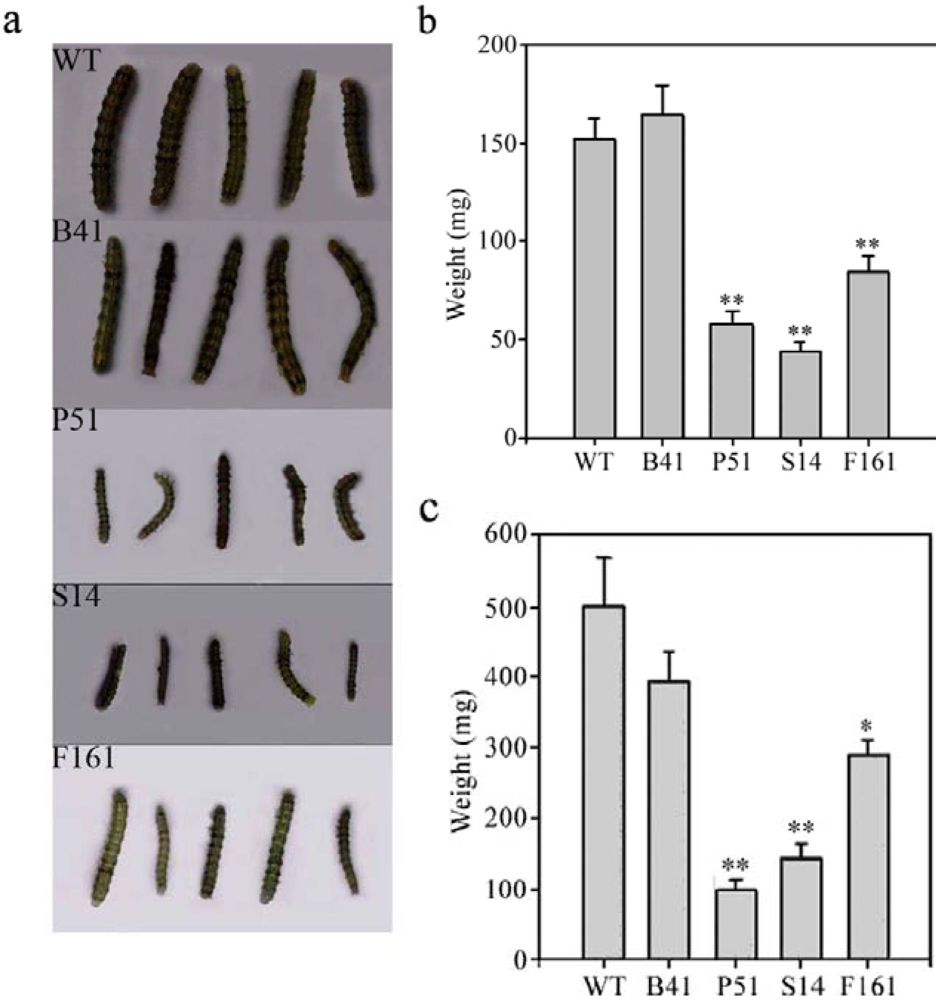
2.3. Insect-resistant activity of transgenic SaPIN2a against H. armigera and S. litura
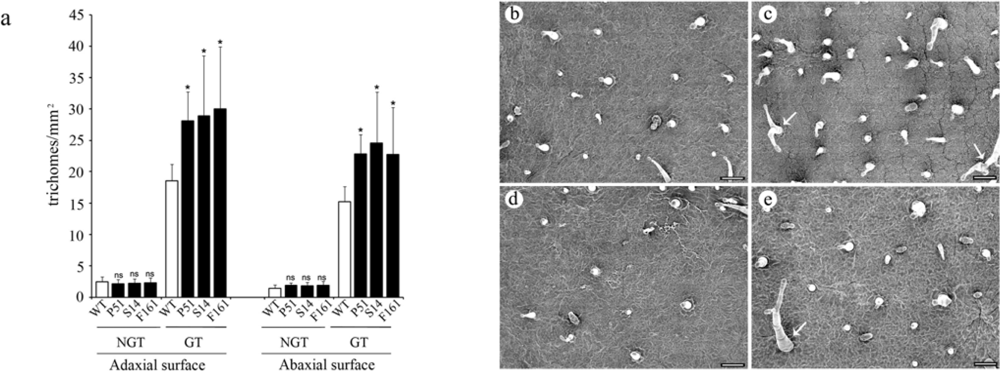
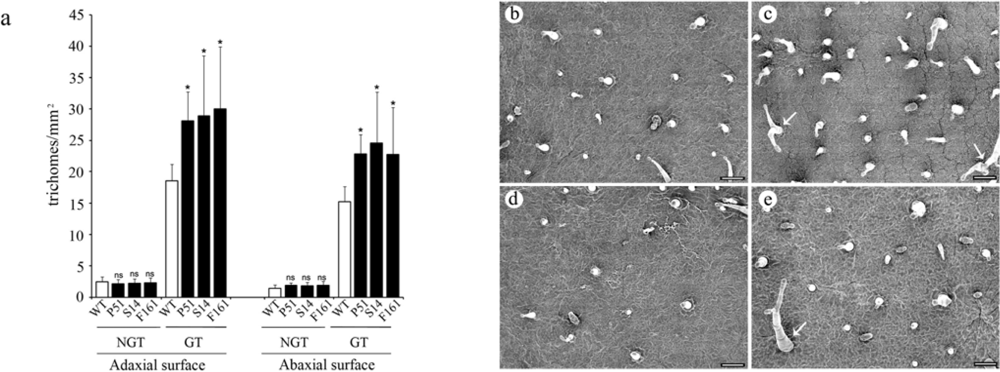
2.4. Effect of transgenic SaPIN2a on trichome development in tobacco
3. Experimental Section
3.1. RNA ligase-mediated rapid amplification of 5′ cDNA end (RLM-5′ RACE)
3.2. Binary vectors and plant transformation
3.3. PCR analysis of transgenic plants
3.4. Northern blot and western blot analysis
3.5. Proteinase inhibitory activity assay
3.6. Scanning electron microscopy
3.7. Insect feeding trials
4. Conclusions
Acknowledgments
References
- Boulter, D. Insect pest control by copying nature using genetically engineered crops. Phytochemistry 1993, 34, 1453–1466. [Google Scholar]
- Mosolov, VV; Valueva, TA. Proteinase inhibitors in plant biotechnology: A review. Appl. Biochem. Microbiol 2008, 44, 233–240. [Google Scholar]
- Ryan, CA. Proteinase inhibitor gene families: Strategies for transformation to improve plant defenses against herbivores. Bioessays 1989, 10, 20–24. [Google Scholar]
- Brzin, J; Kidric, M. Proteinases and their inhibitors in plants: role in normal growth and in response to various stress conditions. Biotechnol. Genet. Eng. Rev 1995, 13, 421–467. [Google Scholar]
- Hilder, VA; Gatehouse, AMR; Sheerman, SE; Barker, RF; Boulter, D. A novel mechanism of insect resistance engineered into tobacco. Nature 1987, 330, 160–163. [Google Scholar]
- Johnson, R; Narvaez, J; An, G; Ryan, C. Expression of proteinase inhibitors I and II in transgenic tobacco plants: effects on natural defense against Manduca sexta larvae. Proc. Natl. Acad. Sci. USA 1989, 86, 9871–9875. [Google Scholar]
- Yeh, KW; Lin, MI; Tuan, SJ; Chen, YM; Lin, CJ; Kao, SS. Sweet potato (Ipomoea batatas) trypsin inhibitors expressed in transgenic tobacco plants confer resistance against Spodoptera litura. Plant Cell Rep 1997, 16, 696–699. [Google Scholar]
- Charity, JA; Anderson, MA; Bittisnich, DJ; Whitecross, M; Higgins, TJV. Transgenic tobacco and peas expressing a proteinase inhibitor from Nicotiana alata have increased insect resistance. Mol. Breed 1999, 5, 357–365. [Google Scholar]
- Bown, DP; Wilkinson, HS; Gatehouse, JA. Differentially regulated inhibitor-sensitive and insensitive protease genes from the phytophagous insect pest, Helicoverpa armigera, are members of complex multigene families. Insect Biochem. Mol. Biol 1997, 27, 625–638. [Google Scholar]
- Haq, SK; Atif, SM; Khan, RH. Protein proteinase inhibitor genes in combat against insects, pests, and pathogens: Natural and engineered phytoprotection. Arch. Biochem. Biophys 2004, 431, 145–159. [Google Scholar]
- Leplé, JC; Bonadé-Bottino, M; Augustin, S; Pilate, G; Lê Tan, VD; Delplanque, A; Cornu, D; Jouanin, L. Toxicity tochrysomela tremulae (Coleoptera: Chrysomelidae) of transgenic poplars expressing a cysteine proteinase inhibitor. Mol. Breed 1995, 1, 319–328. [Google Scholar]
- Altpeter, F; Diaz, I; McAuslane, H; Gaddour, K; Carbonero, P; Vasil, IK. Increased insect resistance in transgenic wheat stably expressing trypsin inhibitor CMe. Mol. Breed 1999, 5, 53–63. [Google Scholar]
- Lee, SI; Lee, SH; Koo, JC; Chun, HJ; Lim, CO; Mun, JH; Song, YH; Cho, MJ. Soybean Kunitz trypsin inhibitor (SKTI) confers resistance to the brown planthopper (Nilaparvata lugens Stal) in transgenic rice. Mol. Breed 1999, 5, 1–9. [Google Scholar]
- Vila, L; Quilis, J; Meynard, D; Breitler, JC; Marfa, V; Murillo, I; Vassal, JM; Messeguer, J; Guiderdoni, E; San Segundo, B. Expression of the maize proteinase inhibitor (mpi) gene in rice plants enhances resistance against the striped stem borer (Chilo suppressalis): effects on larval growth and insect gut proteinases. Plant Biotechnol. J 2005, 3, 187–202. [Google Scholar]
- Jongsma, MA; Bolter, C. The adaptation of insects to plant protease inhibitors. J. Insect Physiol 1997, 43, 885–895. [Google Scholar]
- Harsulkar, AM; Giri, AP; Patankar, AG; Gupta, VS; Sainani, MN; Ranjekar, PK; Deshpande, VV. Successive use of non-host plant proteinase inhibitors required for effective inhibition of Helicoverpa armigera gut proteinases and larval growth. Plant Physiol 1999, 121, 497–506. [Google Scholar]
- Jongsma, MA; Bakker, PL; Peters, J; Bosch, D; Stiekema, WJ. Adaptation of Spodoptera exigua larvae to plant proteinase inhibitors by induction of gut proteinase activity insensitive to inhibition. Proc. Natl. Acad. Sci. USA 1995, 92, 8041–8045. [Google Scholar]
- Wu, Y; Llewellyn, D; Mathews, A; Dennis, ES. Adaptation of Helicoverpa armigera (Lepidoptera: Noctuidae) to a proteinase inhibitor expressed in transgenic tobacco. Mol. Breed 1997, 3, 371–380. [Google Scholar]
- Girard, C; Bonade-Bottino, M; Pham-Delegue, MH; Jouanin, L. Two strains of cabbage seed weevil (Coleoptera : Curculionidae) exhibit differential susceptibility to a transgenic oilseed rape expressing oryzacystatin I. J. Insect Physiol 1998, 44, 569–577. [Google Scholar]
- Cloutier, C; Jean, C; Fournier, M; Yelle, S; Michaud, D. Adult Colorado potato beetles, Leptinotarsa decemlineata compensate for nutritional stress on oryzacystatin I-transgenic potato plants by hypertrophic behavior and over-production of insensitive proteases. Arch. Insect Biochem. Physiol 2000, 44, 69–81. [Google Scholar]
- Xu, ZF; Qi, WQ; Ouyang, XZ; Yeung, E; Chye, ML. A proteinase inhibitor II of Solanum americanum is expressed in phloem. Plant Mol. Biol 2001, 47, 727–738. [Google Scholar]
- Wang, ZY; Ding, LW; Ge, ZJ; Wang, Z; Wang, F; Li, N; Xu, ZF. Purification and characterization of native and recombinant SaPIN2a, a plant sieve element-localized proteinase inhibitor. Plant Physiol. Biochem 2007, 45, 757–766. [Google Scholar]
- Fitt, GP. The ecology of heliothis species in relation to agroecosystems. Annu. Rev. Entomol 1989, 34, 17–52. [Google Scholar]
- Ahmad, M; Saleem, MA; Sayyed, AH. Efficacy of insecticide mixtures against pyrethroid-and organophosphate-resistant populations of Spodoptera litura Lepidoptera: Noctuidae. Pest Manage. Sci 2009, 65, 266–274. [Google Scholar]
- Xu, ZF; Teng, WL; Chye, ML. Inhibition of endogenous trypsin- and chymotrypsin-like activities in transgenic lettuce expressing heterogeneous proteinase inhibitor SaPIN2a. Planta 2004, 218, 623–629. [Google Scholar]
- Xie, J; Ouyang, XZ; Xia, KF; Huang, YF; Pan, WB; Cai, YP; Xu, XP; Li, BJ; Xu, ZF. Chloroplast-like organelles were found in enucleate sieve elements of transgenic plants overexpressing a proteinase inhibitor. Biosci. Biotech. Biochem 2007, 71, 2759–2765. [Google Scholar]
- Satoh, J; Kato, K; Shinmyo, A. The 5′-untranslated region of the tobacco alcohol dehydrogenase gene functions as an effective translational enhancer in plant. J. Biosci. Bioeng 2004, 98, 1–8. [Google Scholar]
- Sugio, T; Satoh, J; Matsuura, H; Shinmyo, A; Kato, K. The 5′-untranslated region of the Oryza sativa alcohol dehydrogenase gene functions as a translational enhancer in monocotyledonous plant cells. J. Biosci. Bioeng 2008, 105, 300–302. [Google Scholar]
- Dansako, T; Kato, K; Satoh, J; Sekine, M; Yoshida, K; Shinmyo, A. 5′ Untranslated region of the HSP18.2 gene contributes to efficient translation in plant cells. J. Biosci. Bioeng 2003, 95, 52–58. [Google Scholar]
- Abdeen, A; Virgos, A; Olivella, E; Villanueva, J; Aviles, X; Gabarra, R; Prat, S. Multiple insect resistance in transgenic tomato plants over-expressing two families of plant proteinase inhibitors. Plant Mol. Biol 2005, 57, 189–202. [Google Scholar]
- Chye, ML; Sin, SF; Xu, ZF; Yeung, EC. Serine proteinase inhibitor proteins: Exogenous and endogenous functions. In Vitro Cell. Dev. Biol. Plant 2006, 42, 100–108. [Google Scholar]
- Telang, M; Srinivasan, A; Patankar, A; Harsulkar, A; Joshi, V; Damle, A; Deshpande, V; Sainani, M; Ranjekar, P; Gupta, G; Birah, A; Rani, S; Kachole, M; Giri, A; Gupta, V. Bitter gourd proteinase inhibitors: potential growth inhibitors of Helicoverpa armigera and Spodoptera litura. Phytochemistry 2003, 63, 643–652. [Google Scholar]
- Harsulkar, AM; Giri, AP; Gupta, VS; Sainani, MN; Deshpande, VV; Patankar, AG; Ranjekar, PK. Characterization of Helicoverpa armigera gut proteinases and their interaction with proteinase inhibitors using gel X-ray film contact print technique. Electrophoresis 1998, 19, 1397–1402. [Google Scholar]
- Bhattacharyya, A; Mazumdar Leighton, S; Babu, CR. Bioinsecticidal activity of Archidendron ellipticum trypsin inhibitor on growth and serine digestive enzymes during larval development of Spodoptera litura. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol 2007, 145, 669–677. [Google Scholar]
- Tamhane, VA; Chougule, NP; Giri, AP; Dixit, AR; Sainani, MN; Gupta, VS. In vivo and in vitro effect of Capsicum annum proteinase inhibitors on Helicoverpa armigera gut proteinases. Biochim. Biophys. Acta, Gen. Subj 2005, 1722, 156–167. [Google Scholar]
- Sin, SF; Chye, ML. Expression of proteinase inhibitor II proteins during floral development in Solanum americanum. Planta 2004, 219, 1010–1022. [Google Scholar]
- Sin, SF; Yeung, EC; Chye, ML. Downregulation of Solanum americanum genes encoding proteinase inhibitor II causes defective seed development. Plant J 2006, 45, 58–70. [Google Scholar]
- Liu, J; Xia, KF; Zhu, JC; Deng, YG; Huang, XL; Hu, BL; Xu, X; Xu, ZF. The nightshade proteinase inhibitor IIb gene is constitutively expressed in glandular trichomes. Plant Cell Physiol 2006, 47, 1274–1284. [Google Scholar]
- Berger, D; Altmann, T. A subtilisin-like serine protease involved in the regulation of stomatal density and distribution in Arabidopsis thaliana. Genes Dev 2000, 14, 1119–1131. [Google Scholar]
- Tanaka, H; Onouchi, H; Kondo, M; Hara-Nishimura, I; Nishimura, M; Machida, C; Machida, Y. A subtilisin-like serine protease is required for epidermal surface formation in Arabidopsis embryos and juvenile plants. Development 2001, 128, 4681–4689. [Google Scholar]
- Von Groll, U; Berger, D; Altmann, T. The subtilisin-like serine protease SDD1 mediates cell-tocell signaling during arabidopsis stomatal development. Plant Cell 2002, 14, 1527–1539. [Google Scholar]
- Wagner, GJ. Secreting glandular trichomes: more than just hairs. Plant physiol 1991, 96, 675–679. [Google Scholar]
- Amme, S; Rutten, T; Melzer, M; Sonsmann, G; Vissers, JP; Schlesier, B; Mock, HP. A proteome approach defines protective functions of tobacco leaf trichomes. Proteomics 2005, 5, 2508–2518. [Google Scholar]
- Peter, AJ; Shanower, TG; Romeis, J. The role of plant trichomes in insect resistance: a selective review. Phytophaga (Madras) 1995, 7, 41–64. [Google Scholar]
- Neal, JJ; Steffens, JC; Tingey, WM. Glandular trichomes of Solanum berthaultii and resistance to the Colorado potato beetle. Entomol. Exp. Appl 1989, 51, 133–140. [Google Scholar]
- Lai, A; Cianciolo, V; Chiavarini, S; Sonnino, A. Effects of glandular trichomes on the development of Phytophthora infestans infection in potato (S. tuberosum). Euphytica 2000, 114, 165–174. [Google Scholar]
- Ascensao, L; Marques, N; Pais, MS. Glandular trichomes on vegetative and reproductive organs of Leonotis leonurus (Lamiaceae). Ann. Bot 1995, 75, 619–626. [Google Scholar]
- Fernandez-Munoz, R; Salinas, M; Alvarez, M; Cuartero, J. Inheritance of resistance to two-spotted spider mite and glandular leaf trichomes in wild tomato Lycopersicon pimpinellifolium (Jusl.) Mill. J. Am. Soc. Hortic. Sci 2003, 128, 188–195. [Google Scholar]
- Simmons, AT; Gurr, GM; McGrath, D; Nicol, HI; Martin, PM. Trichomes of Lycopersicon spp. and their effect on Myzus persicae (Sulzer) (Hemiptera : Aphididae). Aust. J. Entomol 2003, 42, 373–378. [Google Scholar]
- Simmons, AT; Gurr, GM; McGrath, D; Martin, PM; Nicol, HI. Entrapment of Helicoverpa armigera (Hubner) (Lepidoptera : Noctuidae) on glandular trichomes of Lycopersicon species. Aust. J. Entomol 2004, 43, 196–200. [Google Scholar]
- Saeidi, Z; Mallik, B; Kulkarni, RS. Inheritance of glandular trichomes and two-spotted spider mite resistance in cross Lycopersicon esculentum “Nandi” and L pennellii “LA2963”. Euphytica 2007, 154, 231–238. [Google Scholar]
- Aruna, R; Rao, DM; Reddy, LJ; Upadhyaya, HD; Sharma, HC. Inheritance of trichomes and resistance to pod borer (Helicoverpa armigera) and their association in interspecific crosses between cultivated pigeonpea (Cajanus cajan) and its wild relative C-scarabaeoides. Euphytica 2005, 145, 247–257. [Google Scholar]
- Gregory, P; Tingey, WM; Ave, DA; Bouthyette, PY. Potato glandular trichomes: a physic chemical defense mechanism against insects. ACS Symp. Ser 1986, 296, 160–167. [Google Scholar]
- Wagner, GJ; Wang, E; Shepherd, RW. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot 2004, 93, 3–11. [Google Scholar]
- Mcmanus, MT; White, DWR; Mcgregor, PG. Accumulation of a Chymotrypsin Inhibitor in Transgenic Tobacco Can Affect the Growth of Insect Pests. Transgenic Res 1994, 3, 50–58. [Google Scholar]
- Jefferson, RA; Kavanagh, TA; Bevan, MW. GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 1987, 6, 3901–3907. [Google Scholar]
- Hofgen, R; Willmitzer, L. Storage of competent cells for Agrobacterium transformation. Nucleic Acids Res 1988, 16, 9877. [Google Scholar]
- Horsch, RB; Fry, JE; Hoffmann, NL; Eichholtz, D; Rogers, SG; Fraley, RT. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Doyle, JJ; Doyle, JL. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mortality (%) | Pupation rate (%) | |||
|---|---|---|---|---|
| H. armigera | S. litura | H. armigera | S. litura | |
| WT | 16.7 ± 3.3 | 43.3 ± 3.3 | 83.8 ± 4.4 | 82.2 ± 1.1 |
| P51 | 36.7 ± 3.3 * | 56.7 ± 8.8 * | 47.6 ± 2.4** | 63.9 ± 7.3 * |
| S14 | 50.0 ± 5.8 ** | 66.7 ± 6.7 * | 26.1 ± 3.9** | 50.0 ± 0.0 * |
| F161 | 40.0 ± 5.8 ** | 86.7 ± 6.7 ** | 32.1 ± 6.6** | 33.3 ± 16.7 ** |
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Luo, M.; Wang, Z.; Li, H.; Xia, K.-F.; Cai, Y.; Xu, Z.-F. Overexpression of a Weed (Solanum americanum) Proteinase Inhibitor in Transgenic Tobacco Results in Increased Glandular Trichome Density and Enhanced Resistance to Helicoverpa armigera and Spodoptera litura. Int. J. Mol. Sci. 2009, 10, 1896-1910. https://doi.org/10.3390/ijms10041896
Luo M, Wang Z, Li H, Xia K-F, Cai Y, Xu Z-F. Overexpression of a Weed (Solanum americanum) Proteinase Inhibitor in Transgenic Tobacco Results in Increased Glandular Trichome Density and Enhanced Resistance to Helicoverpa armigera and Spodoptera litura. International Journal of Molecular Sciences. 2009; 10(4):1896-1910. https://doi.org/10.3390/ijms10041896
Chicago/Turabian StyleLuo, Ming, Zhaoyu Wang, Huapeng Li, Kuai-Fei Xia, Yinpeng Cai, and Zeng-Fu Xu. 2009. "Overexpression of a Weed (Solanum americanum) Proteinase Inhibitor in Transgenic Tobacco Results in Increased Glandular Trichome Density and Enhanced Resistance to Helicoverpa armigera and Spodoptera litura" International Journal of Molecular Sciences 10, no. 4: 1896-1910. https://doi.org/10.3390/ijms10041896