Selective Immobilization of Fluorescent Proteins for the Fabrication of Photoactive Materials

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
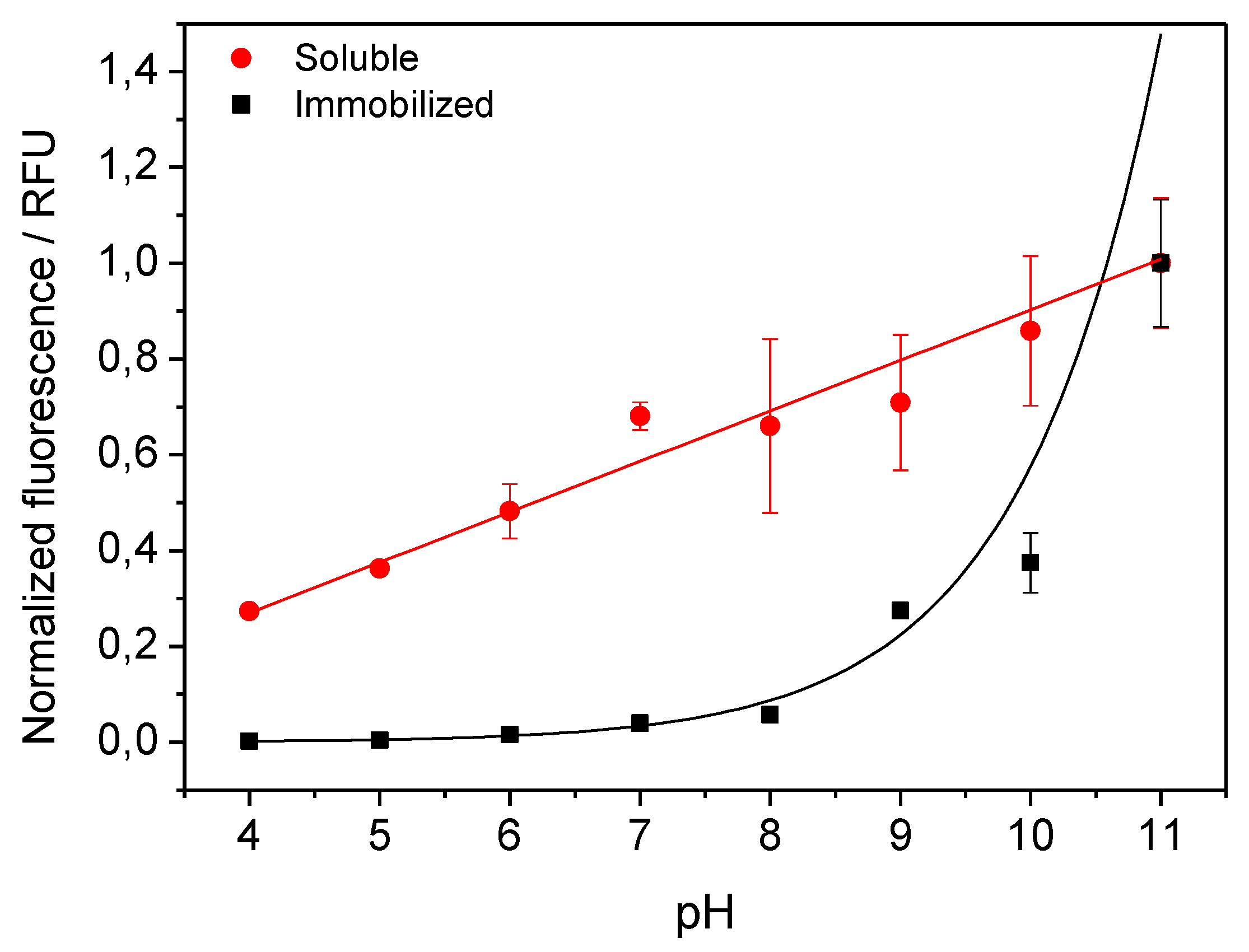
2.1. Selective Immobilization of His-Tagged Fluorescent Proteins on Cobalt-Activated Materials
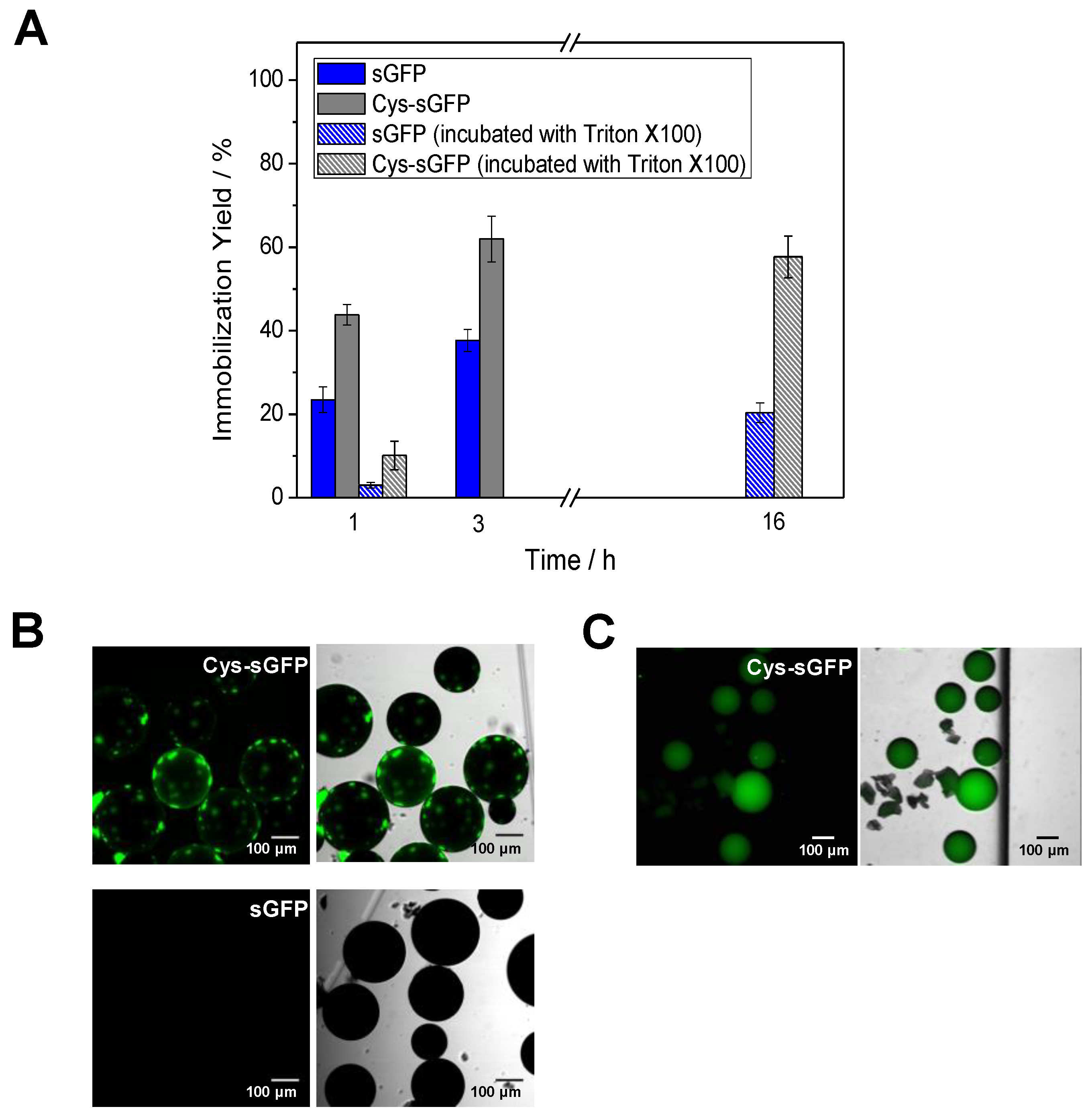
2.2. Direct and Irreversible Immobilization of Cys-Tagged Fluorescent Proteins on Epoxy-Activated Carriers
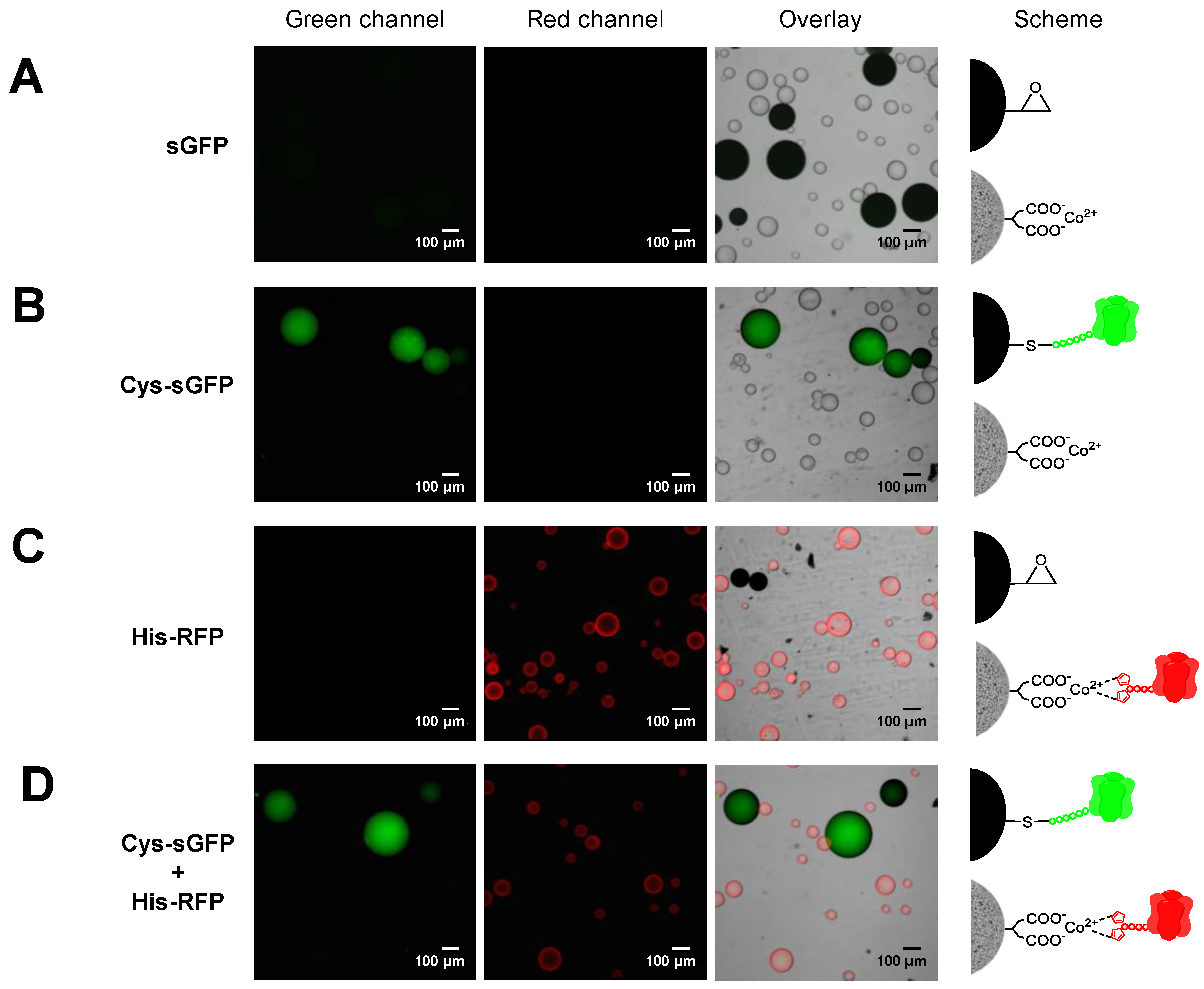
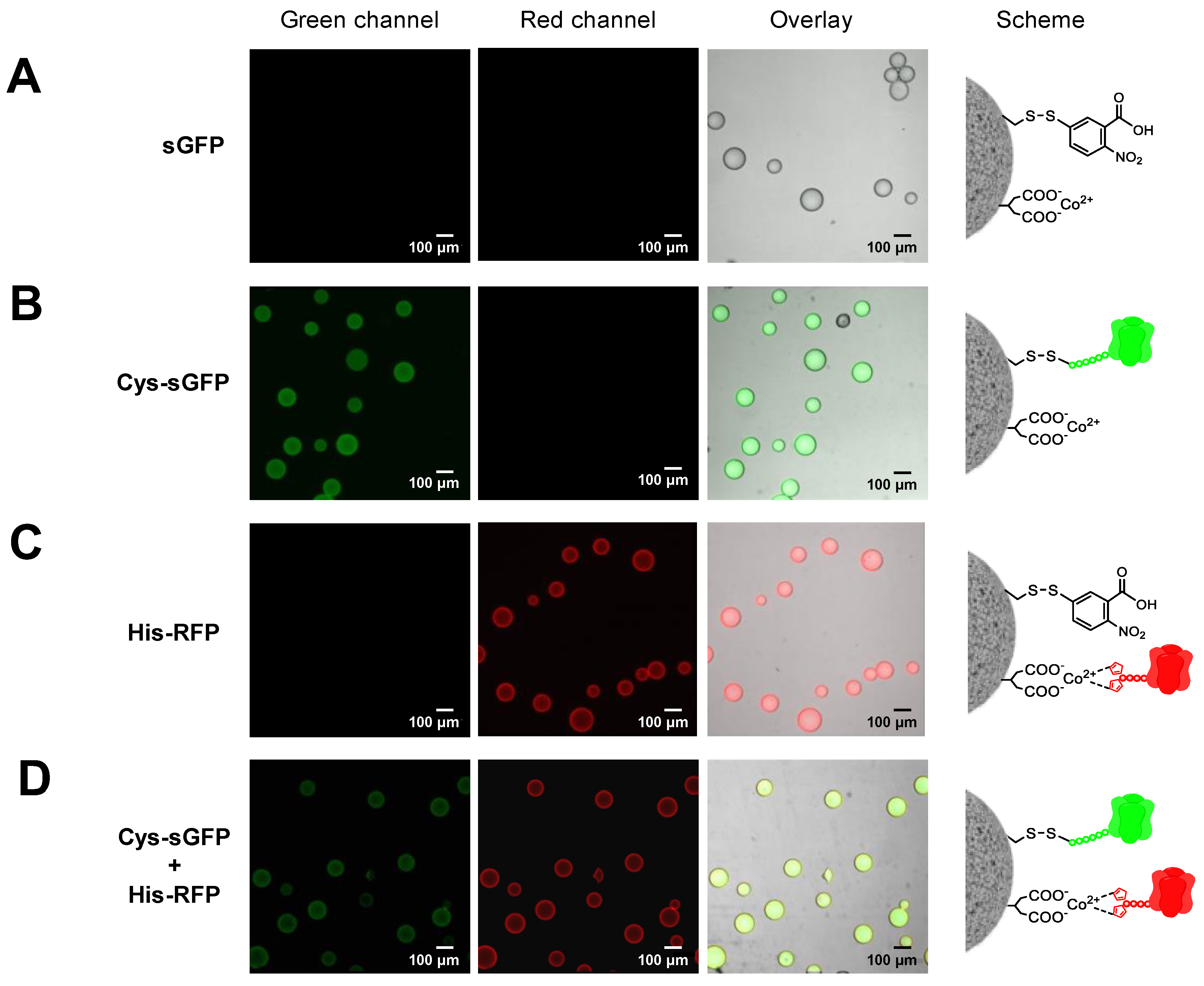
2.3. Genetically Programmed Spatial Distribution of Tagged Proteins. Co-Immobilization of His- and Cys-Tagged Fluorescent Proteins
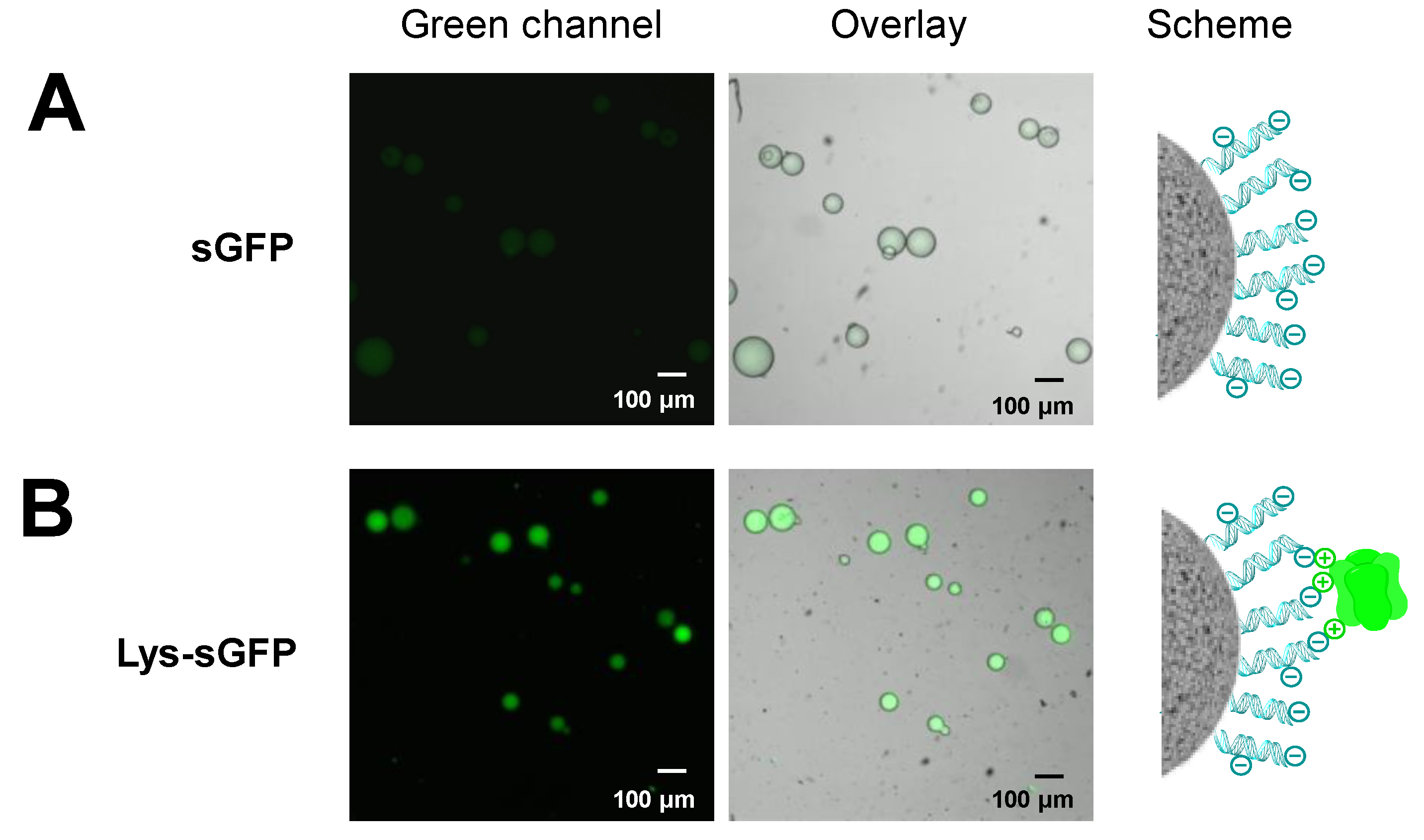
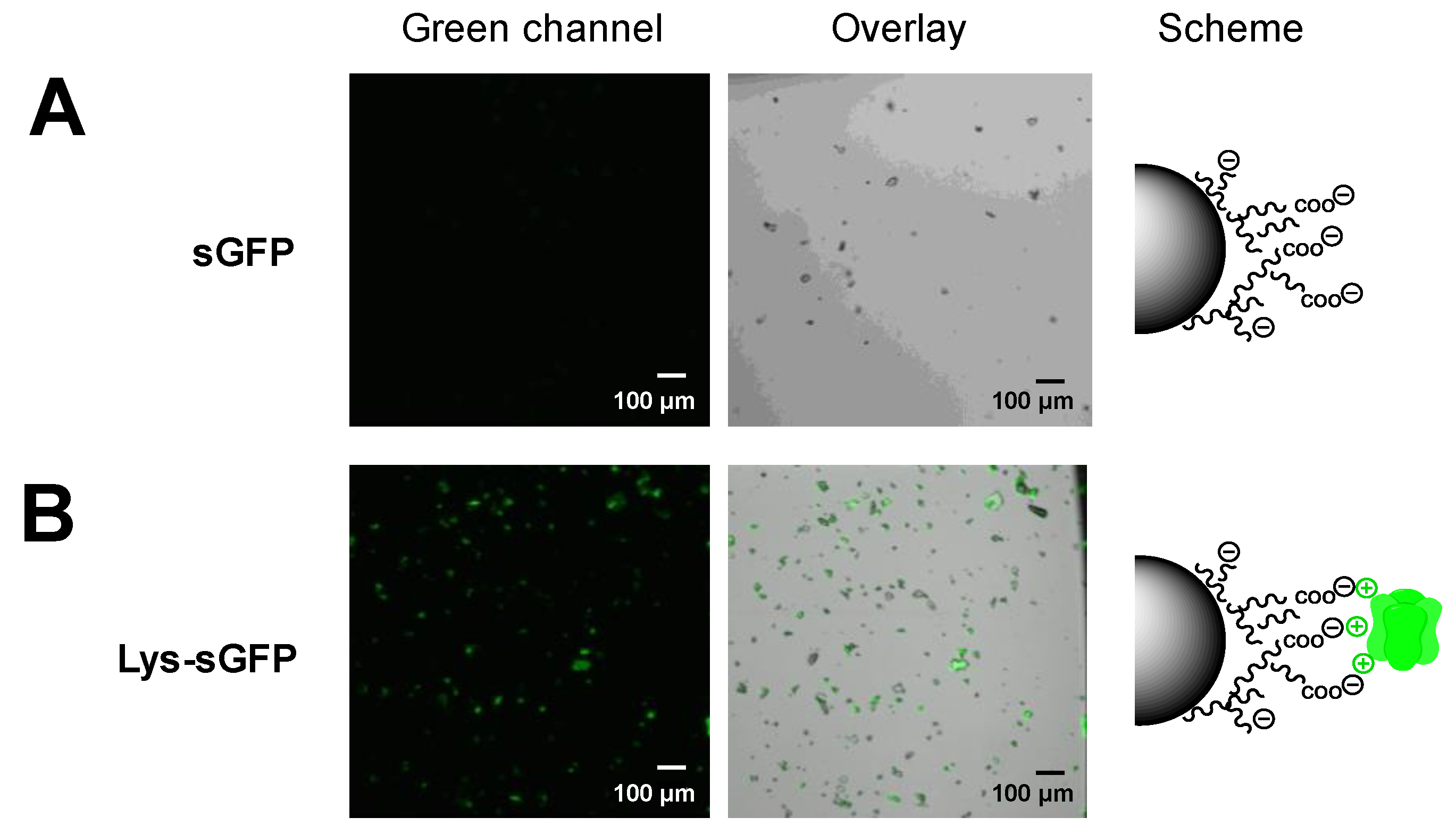
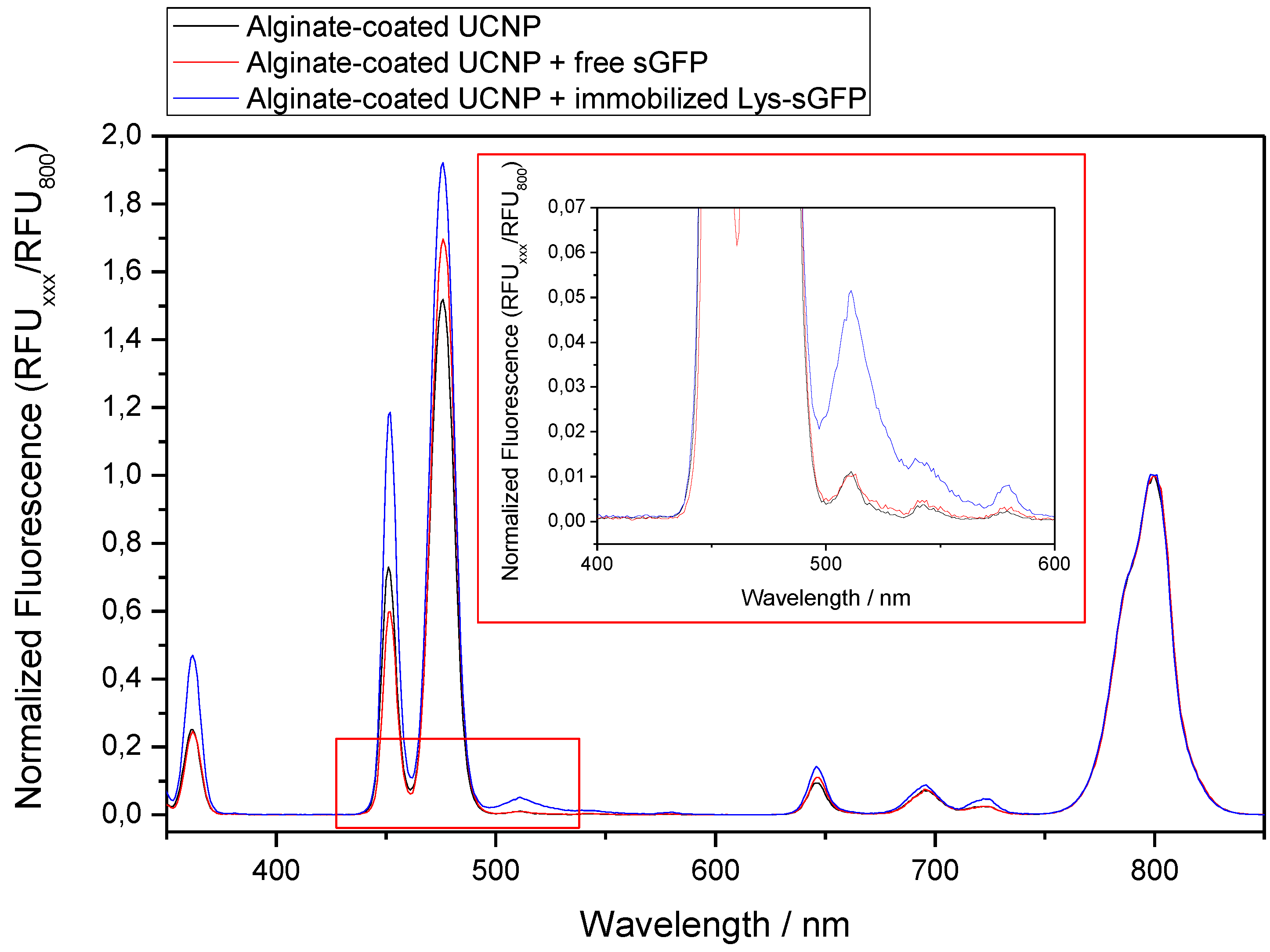
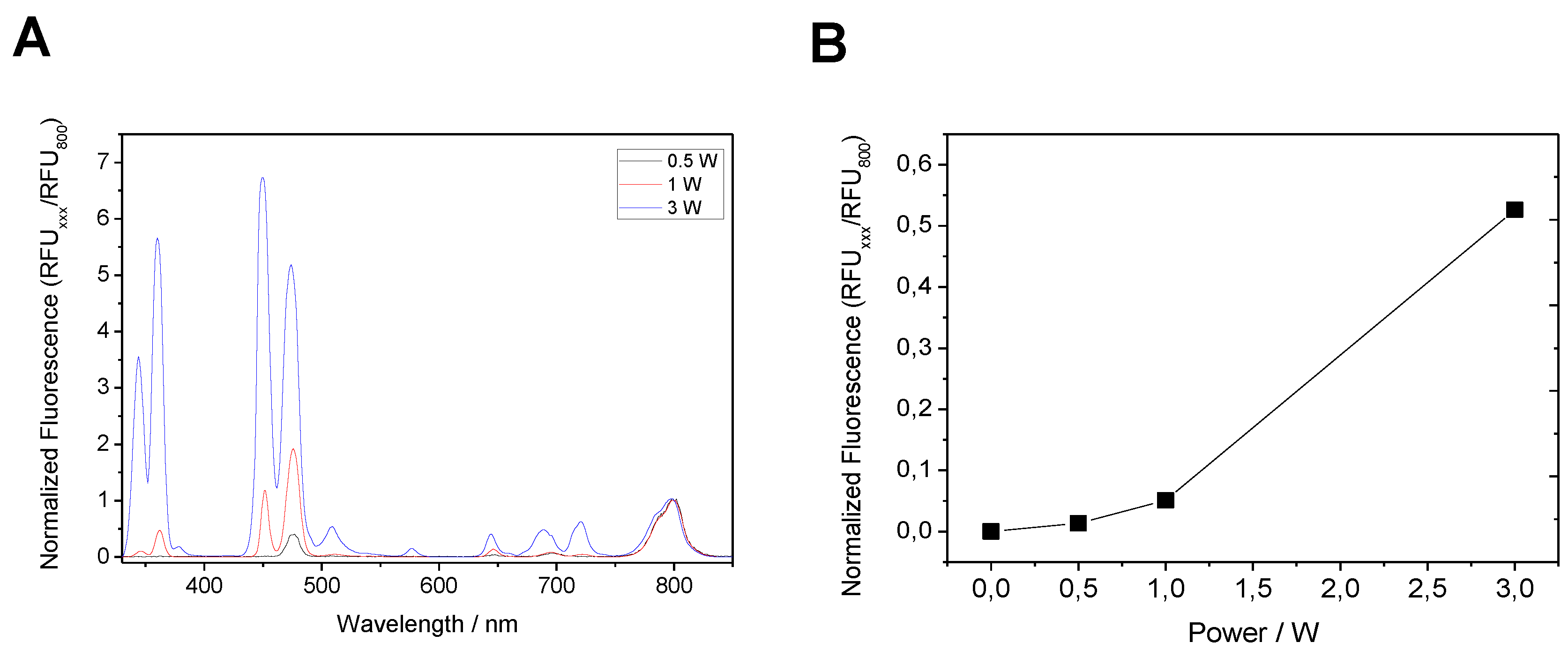
2.4. Selective Immobilization of Lys-Tagged Fluorescent Proteins on Negatively Charged Materials
3. Materials and Methods
3.1. Materials
3.2. Preparation of Supports
3.2.1. Reduced Graphene Composite Structures Functionalized with Epoxy and Cobalt-Chelates (rGOe-Co2+)
3.2.2. Heterofunctional Cobalt- and Thiol-Activated Agarose (AG-Co2+/S)
3.2.3. Agarose Coated with Genomic DNA
3.2.4. Upconverting Nanoparticles Coated with Alginate (UCNP-ALG)
3.3. Protein Expression
3.4. Protein Quantification
3.5. Protein Immobilization
3.6. Silver Staining of Proteins Following Polyacrylamide Gel Electrophoresis
3.7. CLSM Microscopy Imaging
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shimomura, O.; Johnson, F.H.; Saiga, Y. Extraction, purification and properties of aequorin, a bioluminescent protein from the Luminous hydromedusan, Aequorea. J. Cell. Comp. Physiol. 1962, 59, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Day, R.N.; Davidson, M.W. The fluorescent protein palette: Tools for cellular imaging. Chem. Soc. Rev. 2009, 38, 2887–2921. [Google Scholar] [CrossRef] [PubMed]
- Kremers, G.-J.; Gilbert, S.G.; Cranfill, P.J.; Davidson, M.W.; Piston, D.W. Fluorescent proteins at a glance. J. Cell Sci. 2011, 124, 2676. [Google Scholar] [CrossRef]
- Lambert, T.J. FPbase: A community-editable fluorescent protein database. Nat. Methods 2019, 16, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Kneen, M.; Farinas, J.; Li, Y.; Verkman, A.S. Green fluorescent protein as a noninvasive intracellular pH indicator. Biophys. J. 1998, 74, 1591–1599. [Google Scholar] [CrossRef]
- Tantama, M.; Hung, Y.P.; Yellen, G. Imaging intracellular pH in live cells with a genetically encoded red fluorescent protein sensor. J. Am. Chem. Soc. 2011, 133, 10034–10037. [Google Scholar] [CrossRef] [PubMed]
- Pakhomov, A.A.; Martynov, V.I.; Orsa, A.N.; Bondarenko, A.A.; Chertkova, R.V.; Lukyanov, K.A.; Petrenko, A.G.; Deyev, I.E. Fluorescent protein Dendra2 as a ratiometric genetically encoded pH-sensor. Biochem. Biophys. Res. Commun. 2017, 493, 1518–1521. [Google Scholar] [CrossRef]
- Belousov, V.V.; Fradkov, A.F.; Lukyanov, K.A.; Staroverov, D.B.; Shakhbazov, K.S.; Terskikh, A.V.; Lukyanov, S. Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nat. Methods 2006, 3, 281–286. [Google Scholar] [CrossRef]
- Ortega-Villasante, C.; Burén, S.; Blázquez-Castro, A.; Barón-Sola, Á.; Hernández, L.E. Fluorescent in vivo imaging of reactive oxygen species and redox potential in plants. Free Radic. Biol. Med. 2018, 122, 202–220. [Google Scholar] [CrossRef] [Green Version]
- Schwarzländer, M.; Dick, T.P.; Meyer, A.J.; Morgan, B. Dissecting redox biology using fluorescent protein sensors. Antioxid. Redox Signal. 2015, 24, 680–712. [Google Scholar] [CrossRef]
- Deepankumar, K.; Nadarajan, S.P.; Bae, D.-H.; Baek, K.-H.; Choi, K.-Y.; Yun, H. Temperature sensing using red fluorescent protein. Biotechnol. Bioprocess Eng. 2015, 20, 67–72. [Google Scholar] [CrossRef]
- Donner, J.S.; Thompson, S.A.; Kreuzer, M.P.; Baffou, G.; Quidant, R. Mapping intracellular temperature using green fluorescent protein. Nano Lett. 2012, 12, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Veselov, A.A.; Abraham, B.G.; Lemmetyinen, H.; Karp, M.T.; Tkachenko, N.V. Photochemical properties and sensor applications of modified yellow fluorescent protein (YFP) covalently attached to the surfaces of etched optical fibers (EOFs). Anal. Bioanal. Chem. 2012, 402, 1149–1158. [Google Scholar] [CrossRef]
- Makyła, K.; Müller, C.; Lörcher, S.; Winkler, T.; Nussbaumer, M.G.; Eder, M.; Bruns, N. Fluorescent protein senses and reports mechanical damage in glass-fiber-reinforced polymer composites. Adv. Mater. 2013, 25, 2701–2706. [Google Scholar] [CrossRef] [PubMed]
- Futra, D.; Heng, Y.L.; Ahmad, A.; Surif, S.; Ling, L.T. An optical biosensor from green fluorescent Escherichia coli for the evaluation of single and combined heavy metal toxicities. Sensors 2015, 15, 12668–12681. [Google Scholar] [CrossRef] [PubMed]
- Ooi, L.; Heng, L.Y.; Ahmad, A. Toxicity biosensor for sodium dodecyl sulfate using immobilized green fluorescent protein expressing Escherichia coli. J. Sens. 2015, 2015, 9. [Google Scholar] [CrossRef]
- Consolati, T.; Bolivar, J.M.; Petrasek, Z.; Berenguer, J.; Hidalgo, A.; Guisán, J.M.; Nidetzky, B. Biobased, internally pH-sensitive materials: Immobilized yellow fluorescent protein as an optical sensor for spatiotemporal mapping of pH inside porous matrices. ACS Appl. Mater. Interfaces 2018, 10, 6858–6868. [Google Scholar] [CrossRef]
- Fernández-Luna, V.; Coto, P.B.; Costa, R.D. When fluorescent proteins meet white light-emitting diodes. Angew. Chem. Int. Ed. 2018, 57, 8826–8836. [Google Scholar] [CrossRef]
- Weber, M.D.; Niklaus, L.; Pröschel, M.; Coto, P.B.; Sonnewald, U.; Costa, R.D. Bioinspired Hybrid White Light-Emitting Diodes. Adv. Mater. 2015, 27, 5493–5498. [Google Scholar] [CrossRef]
- Couleaud, P.; Adan-Bermudez, S.; Aires, A.; Mejías, S.H.; Sot, B.; Somoza, A.; Cortajarena, A.L. Designed modular proteins as scaffolds to stabilize fluorescent nanoclusters. Biomacromolecules 2015, 16, 3836–3844. [Google Scholar] [CrossRef]
- Mejías, S.H.; López-Andarias, J.; Sakurai, T.; Yoneda, S.; Erazo, K.P.; Seki, S.; Atienza, C.; Martín, N.; Cortajarena, A.L. Repeat protein scaffolds: Ordering photo- and electroactive molecules in solution and solid state. Chem. Sci. 2016, 7, 4842–4847. [Google Scholar] [CrossRef] [PubMed]
- Bolivar, J.M.; Hidalgo, A.; Sánchez-Ruiloba, L.; Berenguer, J.; Guisán, J.M.; López-Gallego, F. Modulation of the distribution of small proteins within porous matrixes by smart-control of the immobilization rate. J. Biotechnol. 2011, 155, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Mateos, A.I.; Llarena, I.; Sánchez-Iglesias, A.; López-Gallego, F. Expanding one-pot cell-free protein synthesis and immobilization for on-demand manufacturing of biomaterials. ACS Synth. Biol. 2018, 7, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.G.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef] [PubMed]
- Block, H.; Maertens, B.; Spriestersbach, A.; Brinker, N.; Kubicek, J.; Fabis, R.; Labahn, J.; Schäfer, F. Immobilized-metal affinity chromatography (IMAC): A review. In Methods in Enzymology; Burgess, R.R., Deutscher, M.P., Eds.; Elsevier: Amsterdam, NL, Holland, 2009; pp. 439–473. [Google Scholar]
- Mateo, C.; Fernández-Lorente, G.; Cortés, E.; Garcia, J.L.; Fernández-Lafuente, R.; Guisan, J.M. One-step purification, covalent immobilization, and additional stabilization of poly-His-tagged proteins using novel heterofunctional chelate-epoxy supports. Biotechnol. Bioeng. 2001, 76, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Grazu, V.; Palomo, J.M.; Lopez-Gallego, F.; Fernandez-Lafuente, R.; Guisan, J.M. Immobilization of enzymes on heterofunctional epoxy supports. Nat. Protoc. 2007, 2, 1022. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, H.; Shannon, M.; Nagai, T. Fluorescent proteins for investigating biological events in acidic environments. Int. J. Mol. Sci. 2018, 19, 1548. [Google Scholar] [CrossRef] [PubMed]
- Benítez-Mateos, A.I.; Contente, M.L.; Velasco-Lozano, S.; Paradisi, F.; López-Gallego, F. Self-sufficient flow-biocatalysis by coimmobilization of pyridoxal 5′-phosphate and ω-transaminases onto porous carriers. ACS Sustain. Chem. Eng. 2018, 6, 13151–13159. [Google Scholar] [CrossRef]
- Velasco-Lozano, S.; Benítez-Mateos, A.I.; López-Gallego, F. Co-immobilized phosphorylated cofactors and enzymes as self-sufficient heterogeneous biocatalysts for chemical processes. Angew. Chem. Int. Ed. 2017, 56, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Velasco-Lozano, S.; da Silva, E.S.; Llop, J.; López-Gallego, F. Sustainable and continuous synthesis of enantiopure L-amino acids by using a versatile immobilised multienzyme system. ChemBioChem 2018, 19, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Zhao, H.; Chang, Y.; Wang, Q.; Yu, H.; Shen, Z. Oriented immobilization and characterization of a poly-lysine-tagged cephalosporin c acylase on glyoxyl agarose support. Appl. Biochem. Biotechnol. 2015, 175, 2114–2123. [Google Scholar] [CrossRef] [PubMed]
- Back, S.H.; Park, J.H.; Cui, C.; Ahn, D.J. Bio-recognitive photonics of a DNA-guided organic semiconductor. Nat. Commun. 2016, 7, 10234. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, E.; Garino, C.; Mareque-Rivas, J.C.; Habtemariam, A.; Salassa, L. Upconverting nanoparticles prompt remote near-infrared photoactivation of Ru(II)–arene complexes. Chem. Eur. J. 2016, 22, 2801–2811. [Google Scholar] [CrossRef] [PubMed]
- Bogdan, N.; Vetrone, F.; Ozin, G.A.; Capobianco, J.A. Synthesis of Ligand-Free Colloidally Stable Water Dispersible Brightly Luminescent Lanthanide-Doped Upconverting Nanoparticles. Nano Lett. 2011, 11, 835–840. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Group | FP | Epoxy Density | Immobilization Yield (%) | Offered (mgFP/gcarrier) | Immobilized (mgFP/gcarrier) |
|---|---|---|---|---|---|
| None | His-RFP | High | 7 | 2 | 0.14 |
| Epoxy | sGFP | High | 97 | 2 | 1.9 |
| His-RFP | Low | 30 | 2 | 0.6 | |
| Medium | 60 | 2 | 1.2 | ||
| High | 95 | 2 | 1.9 | ||
| 59 | 22 | 13 | |||
| 43 | 42 | 18 | |||
| Epoxy and Co2+-chelates | sGFP | High | 98 | 2 | 1.9 |
| His-RFP | High | 99 ± 1 | 2 | 1.9 | |
| 95 | 22 | 21 | |||
| 59 | 42 | 25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benítez-Mateos, A.I.; Mehravar, E.; Velasco-Lozano, S.; Tomovska, R.; Salassa, L.; López-Gallego, F. Selective Immobilization of Fluorescent Proteins for the Fabrication of Photoactive Materials. Molecules 2019, 24, 2775. https://doi.org/10.3390/molecules24152775
Benítez-Mateos AI, Mehravar E, Velasco-Lozano S, Tomovska R, Salassa L, López-Gallego F. Selective Immobilization of Fluorescent Proteins for the Fabrication of Photoactive Materials. Molecules. 2019; 24(15):2775. https://doi.org/10.3390/molecules24152775
Chicago/Turabian StyleBenítez-Mateos, Ana I., Ehsan Mehravar, Susana Velasco-Lozano, Radmila Tomovska, Luca Salassa, and Fernando López-Gallego. 2019. "Selective Immobilization of Fluorescent Proteins for the Fabrication of Photoactive Materials" Molecules 24, no. 15: 2775. https://doi.org/10.3390/molecules24152775