
Integration of Metabolite Profiling and Transcriptome Analysis Reveals Genes Related to Volatile Terpenoid Metabolism in Finger Citron (C. medica var. sarcodactylis)

 , and
, and
Abstract
:1. Introduction
2. Results
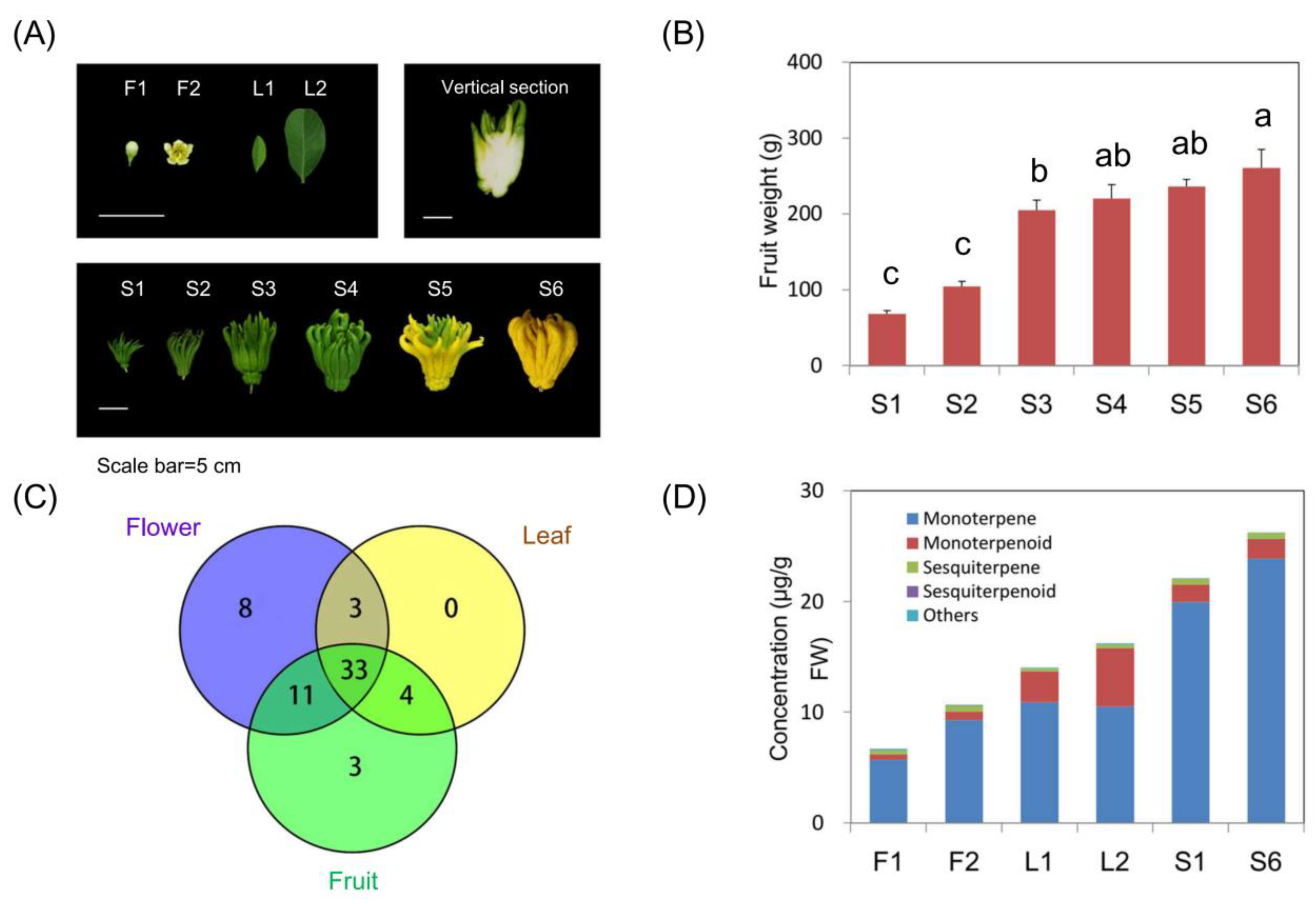
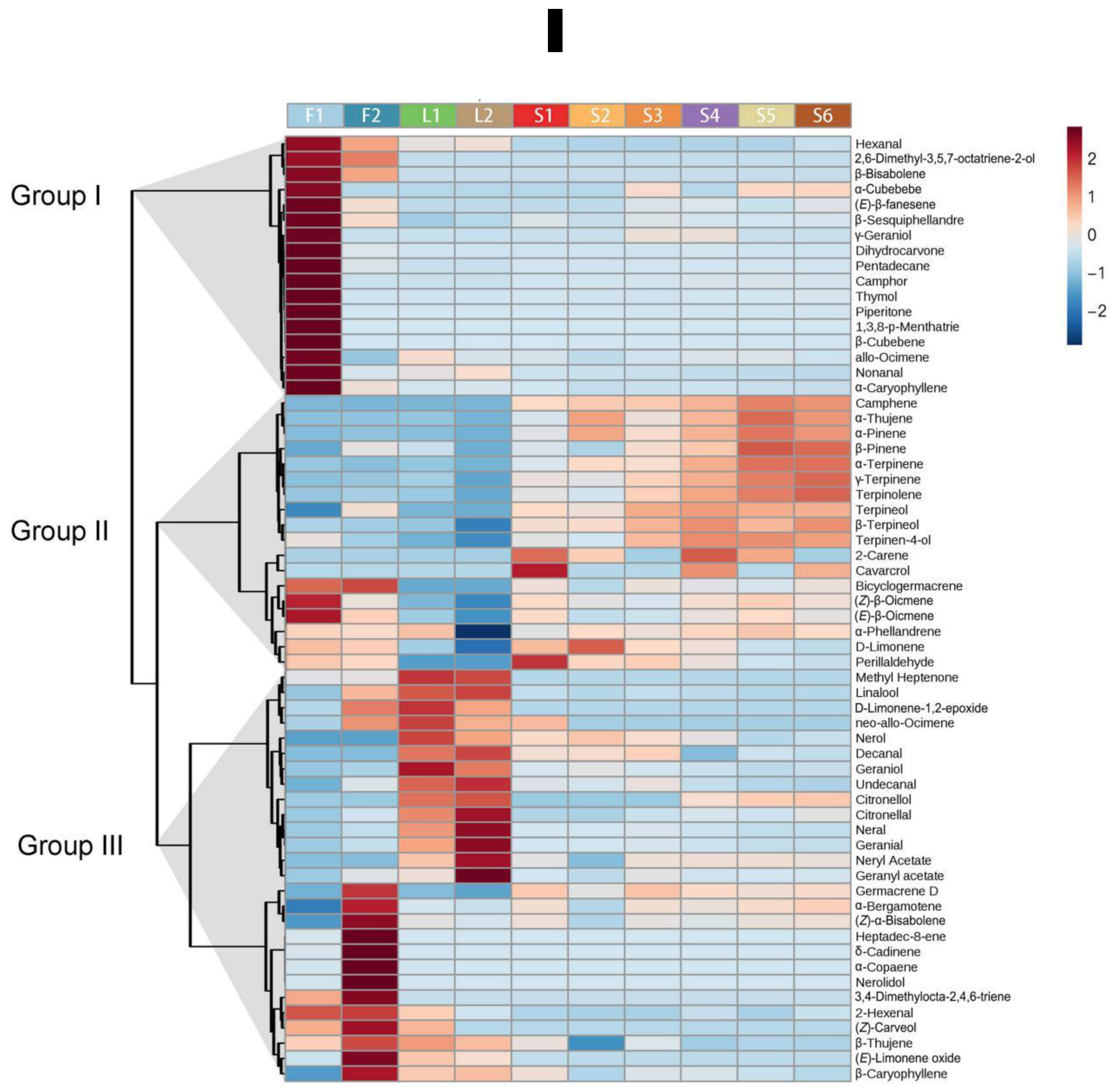
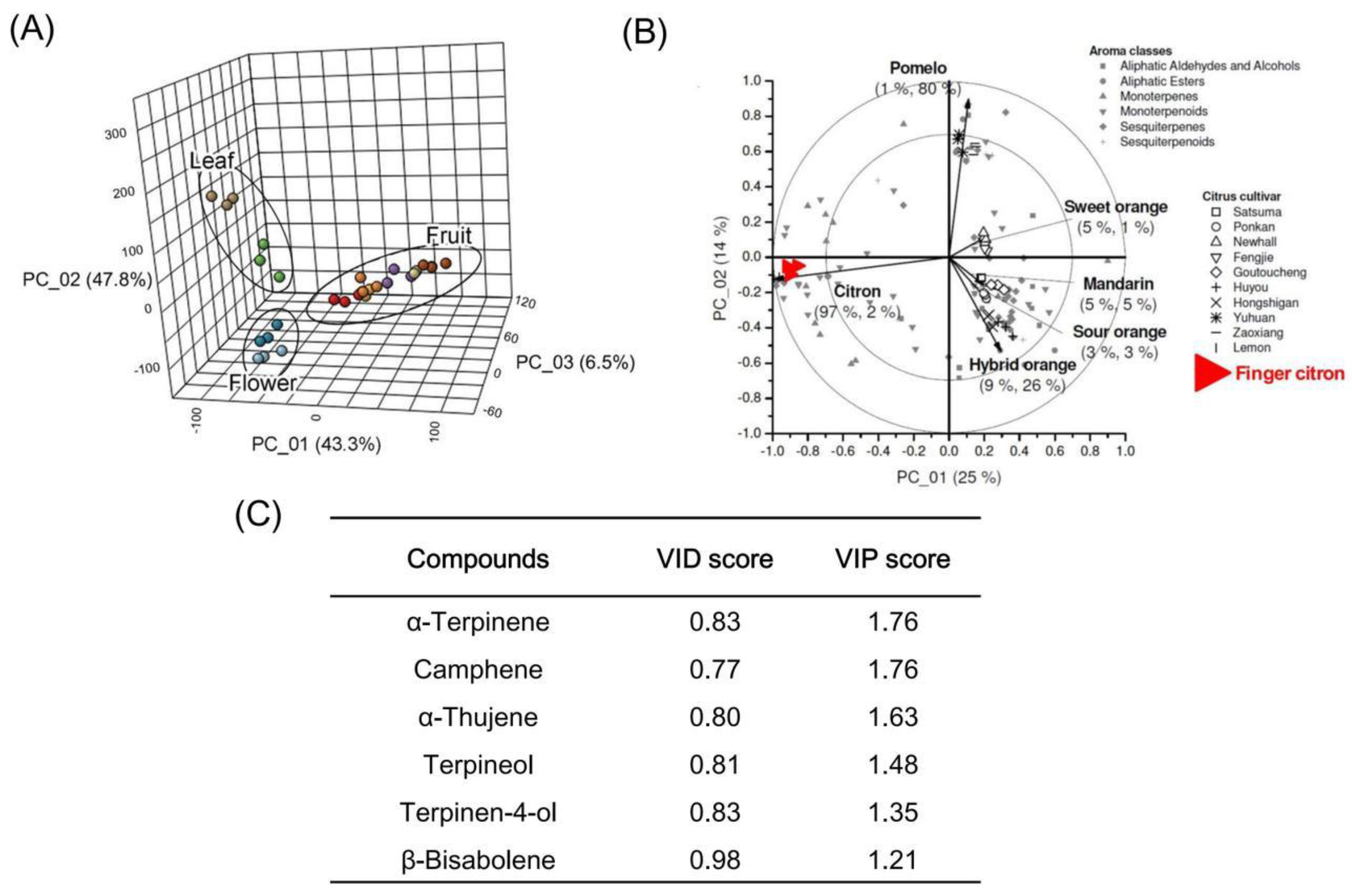
2.1. Volatile Profiling of Finger Citron
2.2. Identification of Key Volatiles
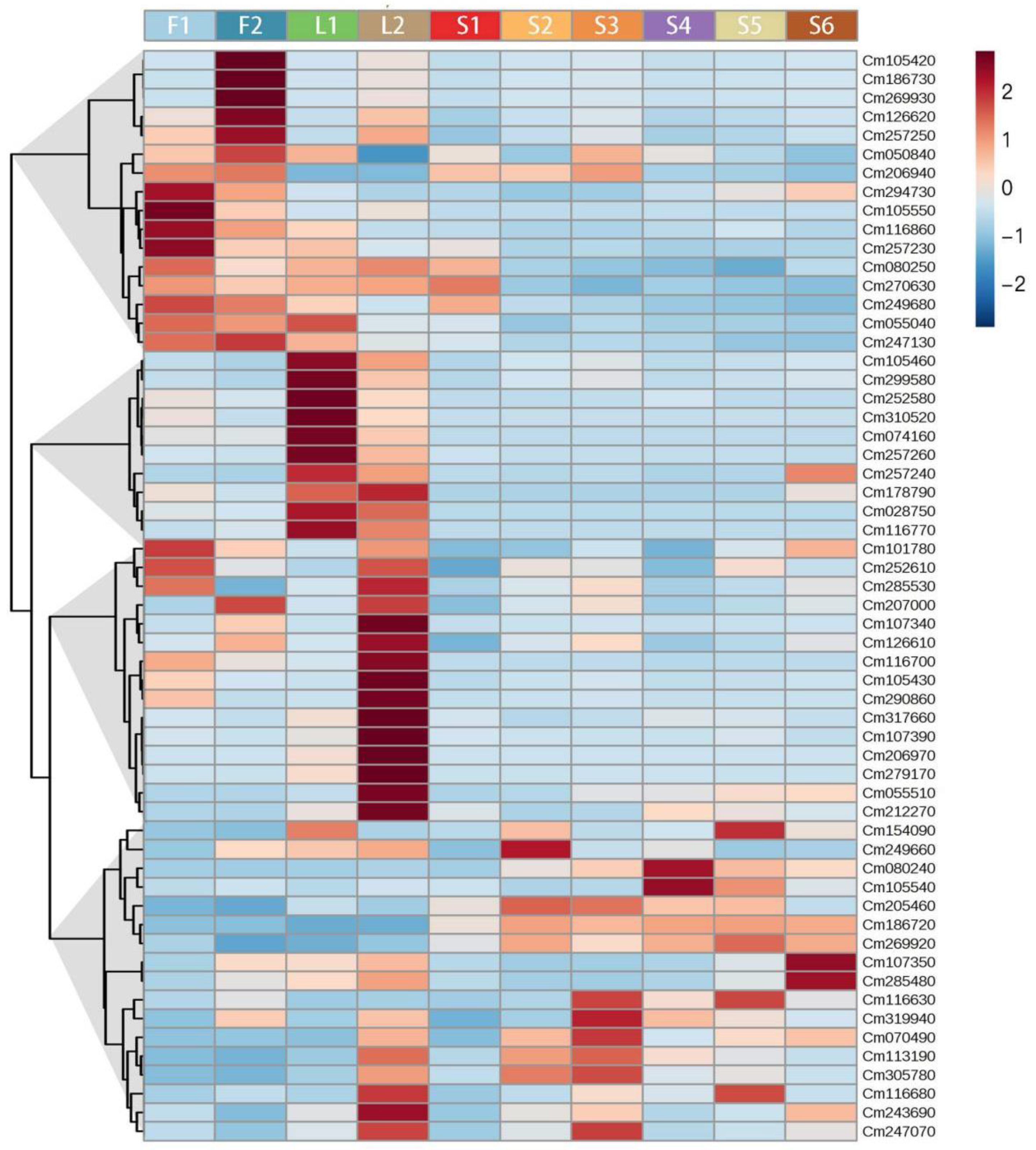
2.3. A Transcriptome Atlas of Finger Citron
2.4. Changes in Transcript Levels of TPS Gene Family
2.5. Changes in Transcript Levels of Transcription Factors
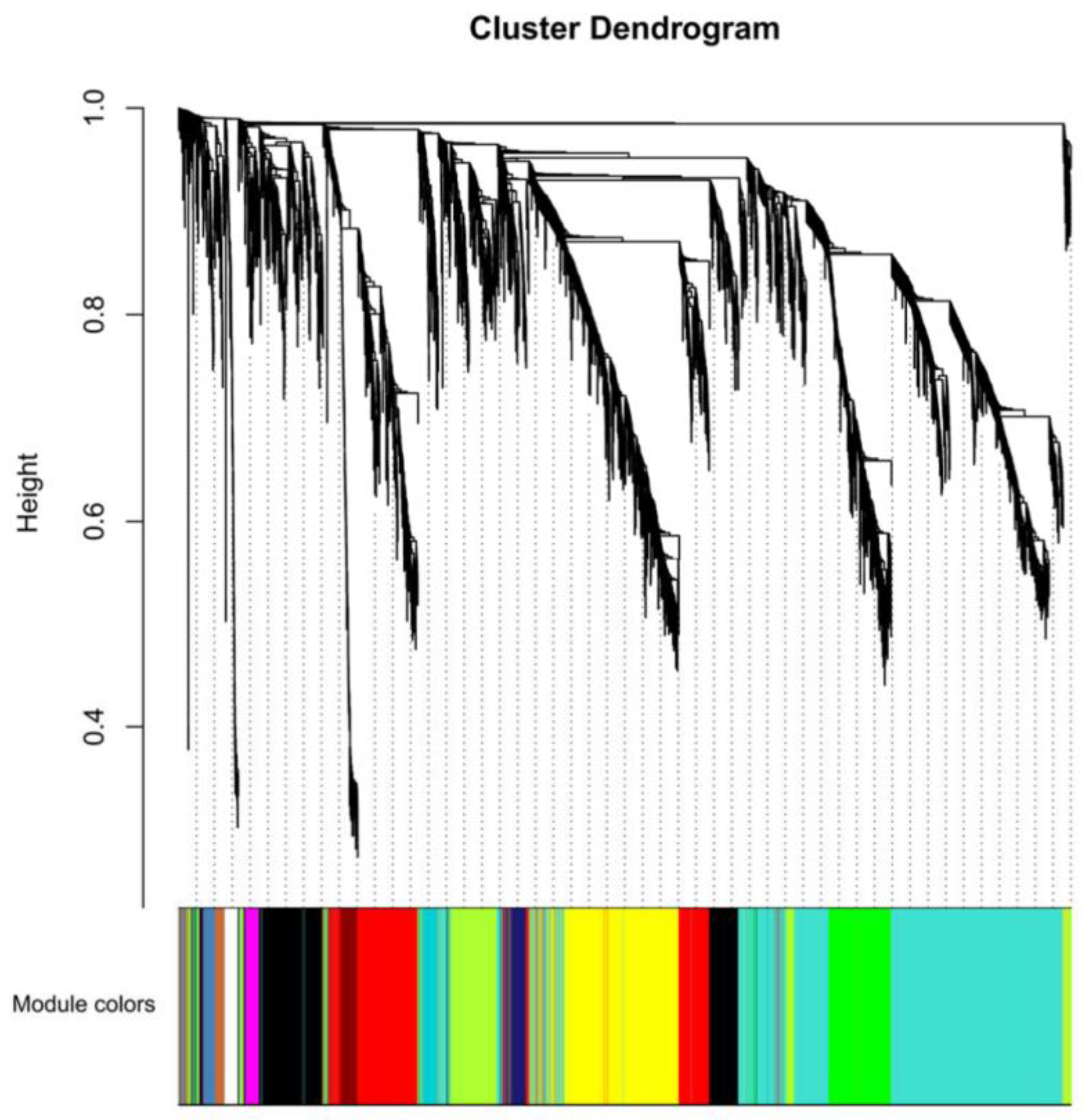
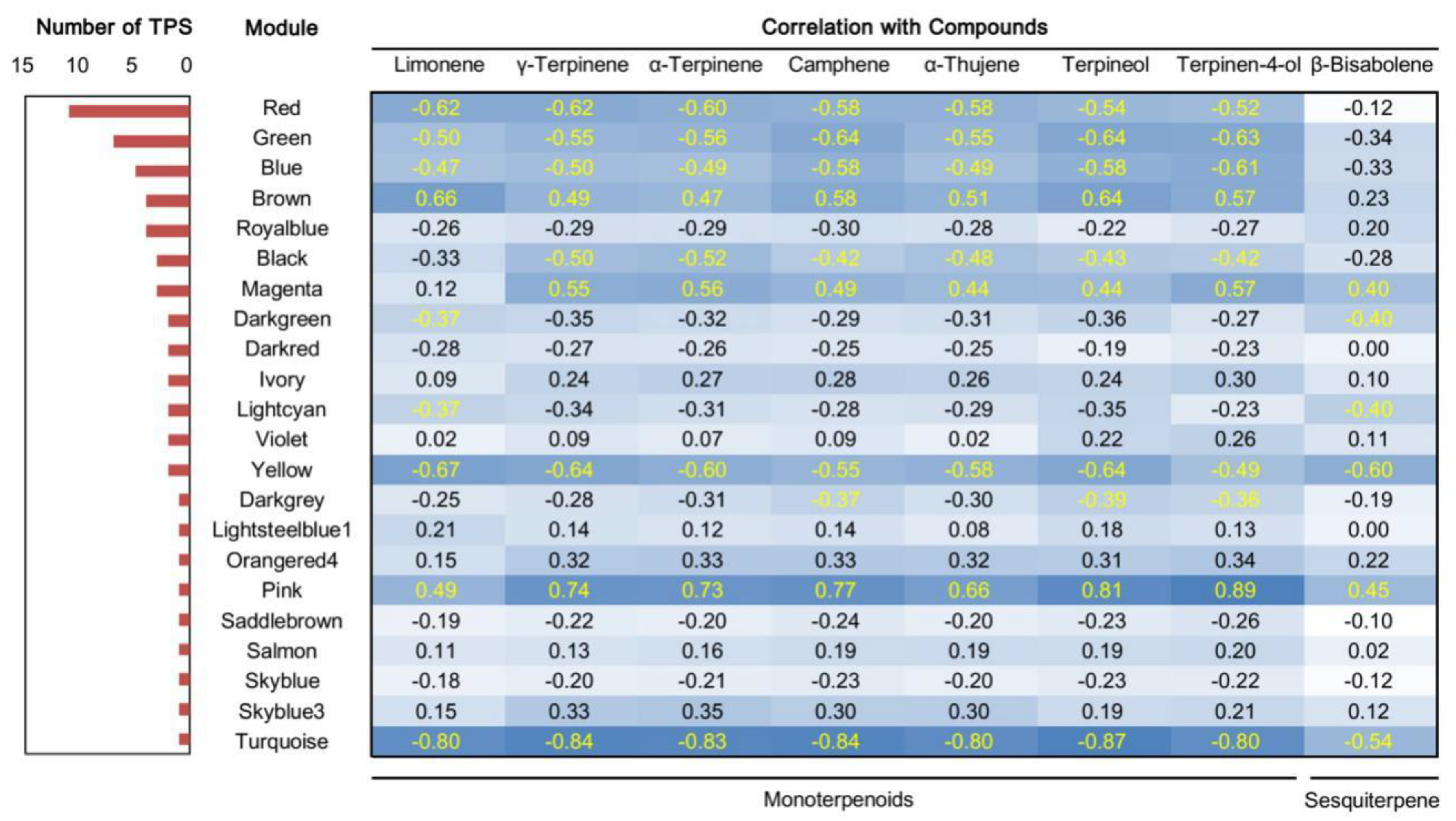
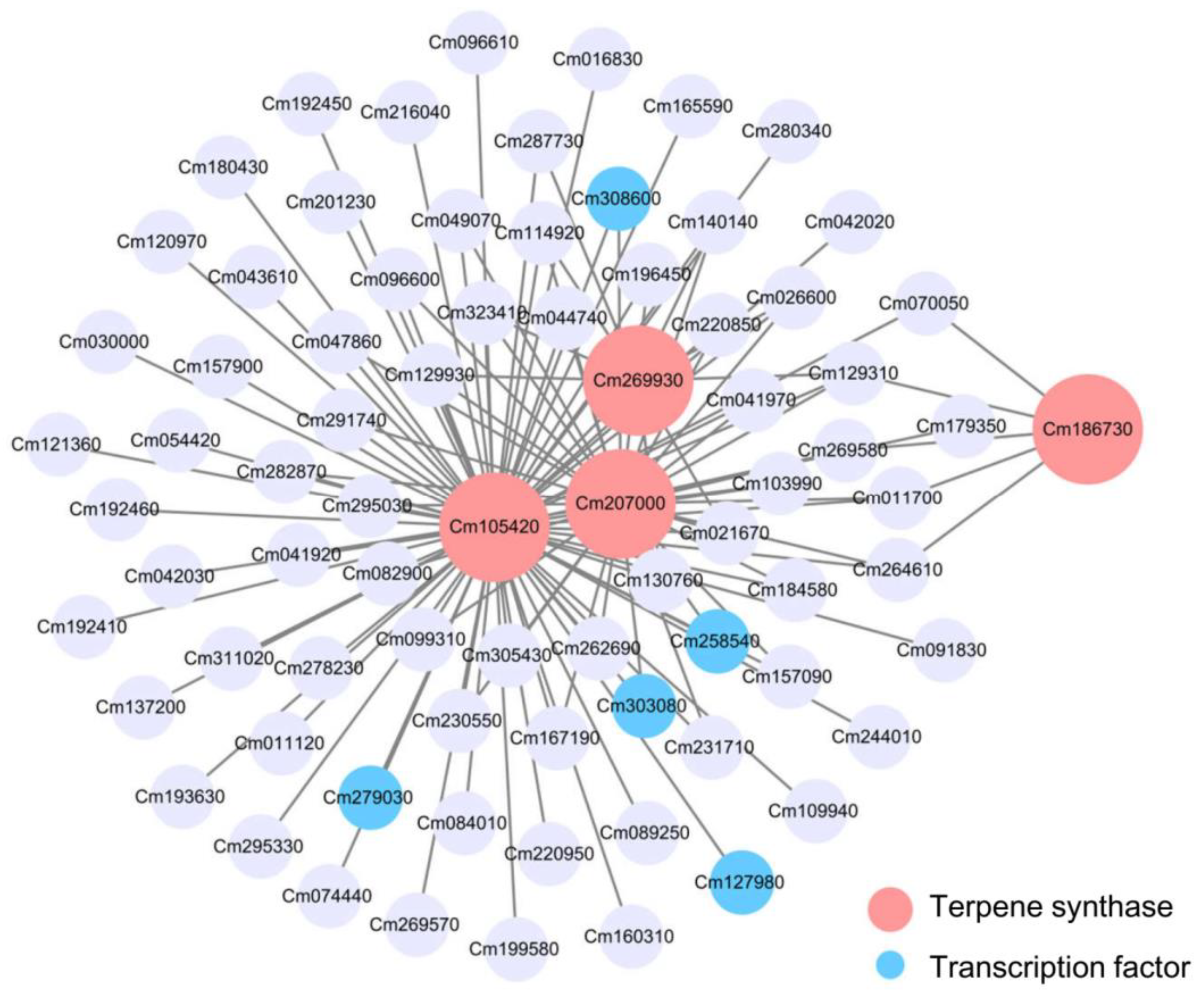
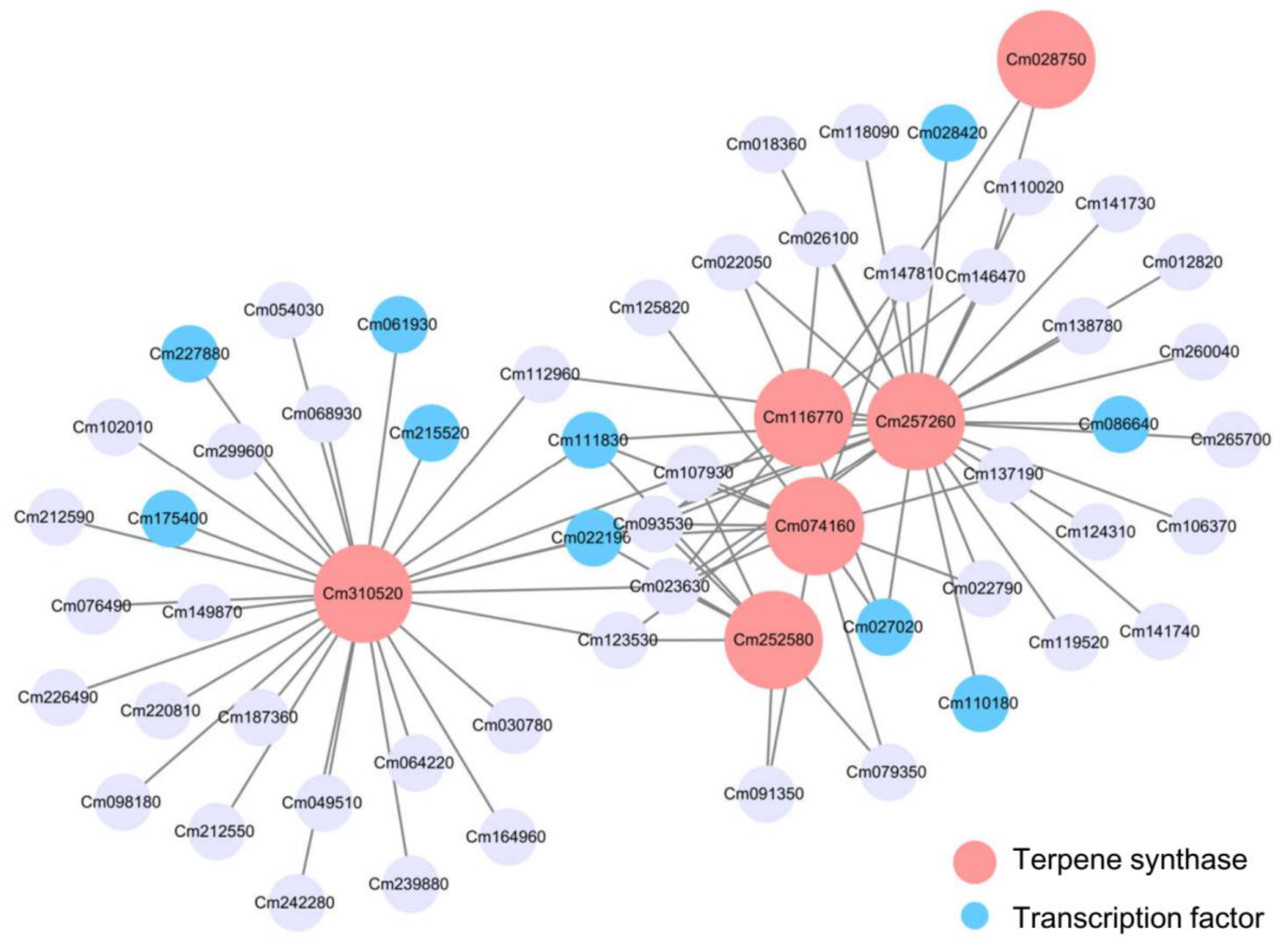
2.6. Analysis of Gene Correlation Network
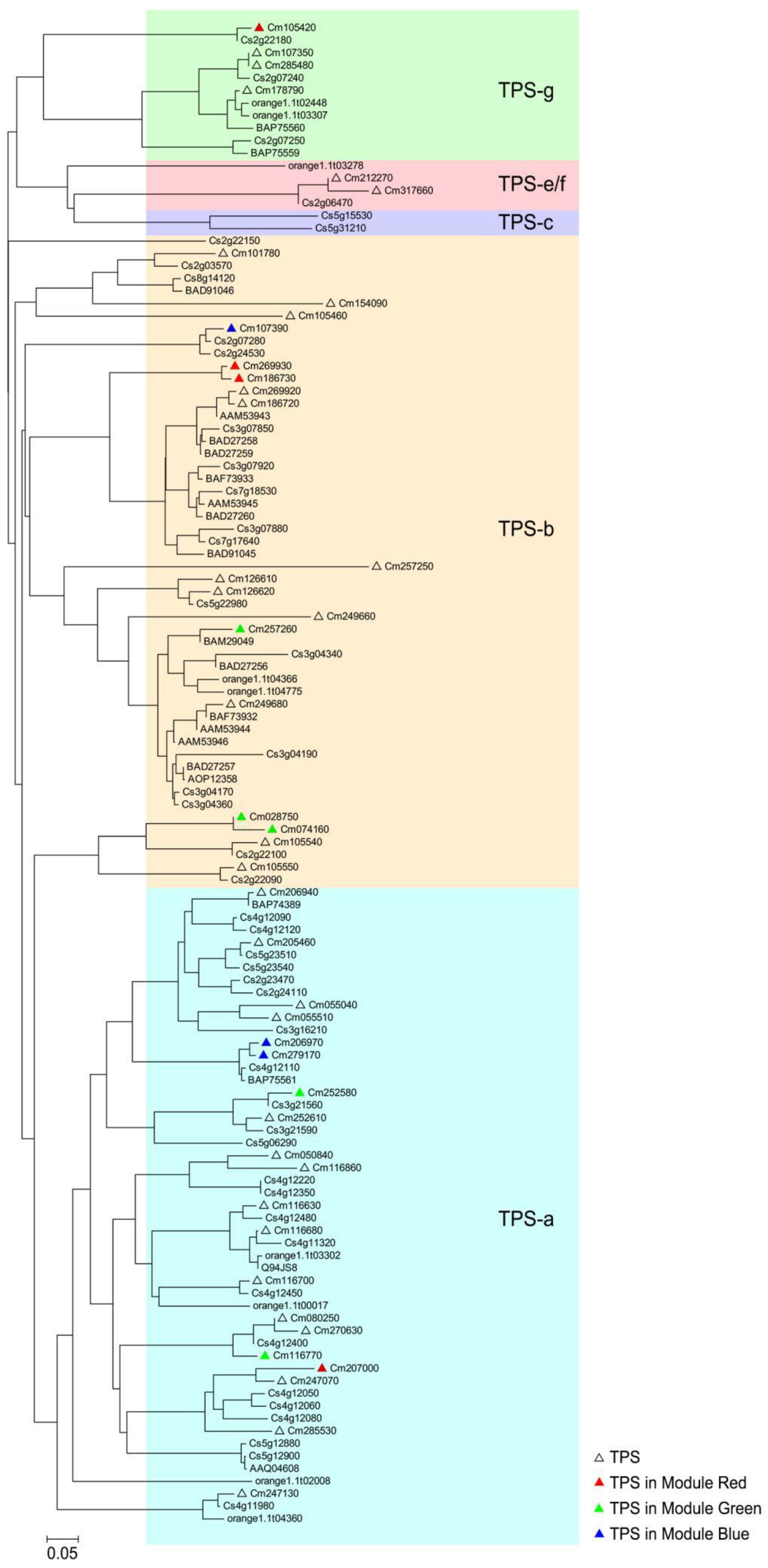
2.7. Phylogenetic Tree Analysis of Finger Citron TPS
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Volatile Collection
4.3. GC-MS Analysis
4.4. Gene Expression Analysis by RNA Sequencing and Real-Time qPCR
4.5. Sequence Analysis and Phylogenetic Tree Construction
4.6. Gene Co-Expression Network Construction
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgements
Conflicts of Interest
References
- Gershenzon, J.; Dudareva, N. The function of terpene natural products in the natural world. Nat. Chem. Biol. 2007, 3, 408–414. [Google Scholar] [CrossRef]
- Vranová, E.; Coman, D.; Gruissem, W. Network analysis of the MVA and MEP pathways for isoprenoid synthesis. Annu. Rev. Plant Biol. 2013, 64, 665–700. [Google Scholar] [CrossRef] [PubMed]
- Kuzuyama, T. Mevalonate and nonmevalonate pathways for the biosynthesis of isoprene units. Biosci. Biotechnol. Biochem. 2002, 66, 1619–1627. [Google Scholar] [CrossRef]
- Degenhardt, J.; Köllner, T.G.; Gershenzon, J. Monoterpene and sesquiterpene synthases and the origin of terpene skeletal diversity in plants. Phytochemistry 2009, 70, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Gershenzon, J. Multiple stress factors and the emission of plant VOCs. Trends Plant Sci. 2010, 15, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Breitmaier, E. Terpenes: Importance, general structure, and biosynthesis. In Terpenes: Flavors, Fragrances, Pharmaca, Pheromones; Breitmaier, E., Ed.; Wiley-VCH: Weinheim, Germany, 2006; Volume 1, pp. 1–3. [Google Scholar]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile compounds in Citrus essential oils: A comprehensive review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef]
- Aubourg, S.; Lecharny, A.; Bohlmann, J. Genomic analysis of the terpenoid synthase (AtTPS) gene family of Arabidopsis thaliana. Mol. Genet. Genom. 2002, 267, 730–745. [Google Scholar]
- Falara, V.; Akhtar, T.A.; Nguyen, T.T.H.; Spytopoulou, E.A.; Bleeker, P.M.; Schauvinhold, I.; Matsuba, Y.; Bonini, M.E.; Schimiller, A.L.; Last, R.L.; et al. The tomato terpene synthase gene family. Plant Physiol. 2011, 157, 770–789. [Google Scholar] [CrossRef]
- Martin, D.M.; Aubourg, S.; Schouwey, M.B.; Daviet, L.; Schalk, M.; Toub, O.; Lund, S.T.; Bohlmann, J. Functional annotation, genome organization and phylogeny of the grapevine (Vitis vinifera) terpene synthase gene family based on genome assembly, FLcDNA cloning, and enzyme assays. BMC Plant Biol. 2010, 10, 226. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuizen, N.J.; Green, S.A.; Chen, X.J.; Bailleul, E.J.D.; Matich, A.J.; Wang, M.Y.; Atkinson, R.G. Functional genomics reveals that a compact terpene synthase gene family can account for terpene volatile production in apple. Plant Physiol. 2013, 161, 787–804. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Lewinsohn, E. Convergent evolution in plant specialized metabolism. Annu. Rev. Plant Biol. 2011, 62, 549–566. [Google Scholar] [CrossRef]
- Dornelas, M.C.; Mazzafera, P. A genomic approach to characterization of the Citrus terpene synthase gene family. Genet. Mol. Biol. 2007, 30, 832–840. [Google Scholar] [CrossRef]
- Alquézar, B.; Rodríguez, A.; Peña, M.D.L.; Peña, L. Genomic analysis of terpene synthase family and functional characterization of seven sesquiterpene synthases from Citrus sinensis. Front. Plant Sci. 2017, 8, 1481. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Endo, T.; Fujii, H.; Hara, M.; Ueda, T.; Kira, M.; Omura, M. Molecular cloning and functional characterization of four monoterpene synthase genes from Citrus unshiu Marc. Plant Sci. 2004, 166, 49–58. [Google Scholar] [CrossRef]
- Shimada, T.; Endo, T.; Fujii, H.; Hara, M.; Omura, M. Isolation and characterization of (E)–β–ocimene and 1,8 cineole synthases in Citrus unshiu Marc. Plant Sci. 2005, 168, 987–995. [Google Scholar] [CrossRef]
- Shimada, T.; Endo, T.; Fujii, H.; Omura, M. Isolation and characterization of a new d-limonene synthase gene with a different expression pattern in Citrus unshiu Marc. Sci. Hortic. 2005, 105, 507–512. [Google Scholar] [CrossRef]
- Shimada, T.; Endo, T.; Rodríguez, A.; Fujii, H.; Nakano, M.; Sugiyama, A.; Shimizu, T.; Peña, L.; Omura, M. Isolation and characterization of germacrene A synthases gene in Citrus unshiu Marc. Sci. Hortic. 2012, 145, 102–108. [Google Scholar] [CrossRef]
- Shimada, T.; Endo, T.; Fujii, H.; Rodríguez, A.; Peña, L.; Omura, M. Characterization of three linalool synthase genes from Citrus unshiu Marc. and analysis of linalool-mediated resistance against Xanthomonas citri subsp. citri and Penicilium italicum in citrus leaves and fruits. Plant Sci. 2014, 229, 154–166. [Google Scholar] [CrossRef]
- Lücker, J.; Tamer, M.K.E.; Schwab, W.; Verstappen, F.W.A.; Plas, L.H.W.V.D.; Bouwmeester, H.J.; Verhoeven, H.A. Monoterpene biosynthesis in lemon (Citrus limon): cDNA isolation and functional analysis of four monoterpene synthases. Eur. J. Biochem. 2010, 269, 3160–3171. [Google Scholar] [CrossRef]
- Sharon–Asa, L.; Shalit, M.; Frydman, A.; Bar, E.; Holland, D.; Or, E.; Lavi, U.; Lewinsohn, E.; Eyal, Y. Citrus fruit flavor and aroma biosynthesis: Isolation functional characterization, and developmental regulation of Cstps1, a key gene in the production of the sesquiterpene aroma compound valencene. Plant J. 2003, 36, 664–674. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Ito, M.; Honda, F. Molecular cloning, functional expression and characterization of (E)-β-farnesene synthase from Citrus junos. Biol. Pharm. Bull. 2001, 24, 1171–1175. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.Y.; Wu, B.P.; Cao, X.M.; Zhang, B.; Chen, K.S. Citrus CmTPS1 is associated with formation of sesquiterpene bicyclogermacrene. Sci. Hortic. 2017, 226, 133–140. [Google Scholar] [CrossRef]
- Shen, S.L.; Yin, X.R.; Zhang, B.; Xie, X.L.; Jiang, Q.; Grierson, D.; Chen, K.S. CitAP2.10 activation of the terpene synthase CsTPS1 is associated with the synthesis of (+)-valencene in ‘Newhall’ orange. J. Exp. Bot. 2016, 67, 4105–4115. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xu, Y.Y.; Shen, S.L.; Yin, X.R.; Klee, H.; Zhang, B.; Chen, K.S. Transcription factor CitERF71 activates the terpene synthase gene CitTPS16 involved in the synthesis of E-geraniol in sweet orange fruit. J. Exp. Bot. 2017, 68, 4929–4938. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.A.; Wang, Q.; Dhar, N.; Kumar, N.; Venkatesh, N.; Rajan, C.; Panicker, D.; Sridhar, V.; Mao, H.Z.; Sarojam, R. Spearmint R2R3-MYB transcription factor MsMYB negatively regulates monoterpene production and suppresses the expression of geranyl diphosphate synthase large subunit (MsGPPS.LSU). Plant Biotechnol. J. 2017, 15, 1105–1119. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.J.; Chen, X.Y.; Wang, M.Y.; Matich, A.J.; Perez, R.L.; Allan, A.C.; Green, S.A.; Atkinson, R.G. Natural variation in monoterpene synthesis in kiwifruit: Transcriptional regulation of terpene synthases by NAC and ETHYLENE-INSENSITIVE3-like transcription factors. Plant Physiol. 2015, 167, 1243–1258. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Wang, J.W.; Wang, S.; Wang, J.Y.; Chen, X.Y. Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-δ-cadinene synthase-A1. Plant Physiol. 2004, 135, 507–515. [Google Scholar] [CrossRef]
- Miyamoto, K.; Nishizawa, Y.; Minami, E.; Nojiri, H.; Yamane, H.; Okada, K. Overexpression of the bZIP transcription factor OsbZIP79 suppresses the production of diterpenoid phytoalexin in rice cells. J. Plant Physiol. 2015, 173, 19–27. [Google Scholar] [CrossRef]
- Hong, G.J.; Xue, X.Y.; Mao, Y.B.; Wang, L.J.; Chen, X.Y. Arabidopsis MYC2 interacts with DELLA proteins in regulating sesquiterpene synthase gene expression. Plant Cell 2012, 24, 2635–2648. [Google Scholar] [CrossRef]
- Yu, Z.X.; Li, J.X.; Yang, C.Q.; Hu, W.L.; Wang, L.J.; Chen, X.Y. The jasmonate-responsive AP2/ERF transcription factors AaERF1 and AaERF2 positively regulate artemisinin biosynthesis in Artemisia annua L. Mol. Plant 2012, 5, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Skinem?Li, L.; Hao, X.L.; Liu, H.; Wang, W.; Fu, X.Q.; Ma, Y.N.; Shen, Q.; Chen, M.H.; Tang, K.X. Jasmonic acid-responsive AabHLH1 positively regulates artemisinin biosynthesis in Artemisia annua. Biotechnol. Appl. Biochem. 2019. [Google Scholar] [CrossRef]
- Wang, X.; Xu, Y.T.; Zhang, S.Q.; Cao, Y.; Huang, Y.; Cheng, J.F.; Wu, G.Z.; Tian, S.L.; Chen, C.L.; Liu, Y.; et al. Genomic analyses of primitive, wild and cultivated citrus provide insights into asexual reproduction. Nature Genet. 2017, 49, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.L.; Wang, Y.J.; Chen, M.G.; Chen, W.R.; Zong, Y.; Li, Y.Q.; Yang, L.; Han, X.X.; Guo, W.D. Low transcription of CmsIAA9 in the basal pistil is related to parthenocarpic fruiting of fingered citron (Foshou). Mol. Breed. 2017, 37, 101. [Google Scholar] [CrossRef]
- Brigand, J.P.; Nahon, P. Gastronomy and the citron tree (Citrus medica L.). Int. J. Gastro. Food Sci. 2016, 3, 12–16. [Google Scholar] [CrossRef]
- Zhang, W.Q. Chapter 37 Citrus medica L. var. sarcodactylis Swingle (foshou, finger citron). In Dietary Chinese Herbs: Chemistry, Pharmacology and Clinical Evidence; Liu, Y.Z., Wang, Z.M., Zhang, J.Z., Eds.; Springer: Friesach, Austria, 2015; pp. 327–331. [Google Scholar]
- Lou, Z.X.; Chen, J.; Yu, F.H.; Wang, H.X.; Kou, X.R.; Ma, C.Y.; Zhu, S. The antioxidant, antibacterial, antibiofilm activity of essential oil from Citrus medica L. var. sarcodactylis and its nanoemulsion. LWT-Food Sci. Technol. 2017, 80, 371–377. [Google Scholar] [CrossRef]
- Peng, C.H.; Ker, Y.B.; Weng, C.F.; Peng, C.C.; Huang, C.N.; Lin, L.Y.; Peng, R.Y. Insulin secretagogue bioactivity of finger citron fruit (Citrus medica L. var. Sarcodactylis Hort, Rutaceae). J. Agric. Food Chem. 2009, 57, 8812–8819. [Google Scholar] [CrossRef]
- Wu, Z.; Li, H.; Yang, Y.; Zhan, Y.; Tu, D.W. Variation in the components and antioxidant activity of Citrus medica L. var. sarcodactylis essential oils at different stages of maturity. Ind. Crop. Prod. 2013, 46, 311–315. [Google Scholar] [CrossRef]
- He, Y.F.; Jin, X.L. Review of research progress in Citrus medica with reference to prediction of development prospects. Econ. For. Res. 2001, 4, 41–43. (In Chinese) [Google Scholar]
- Jing, L.; Lei, Z.T.; Zhang, G.W.; Pilon, A.C.; Huhman, D.V.; Xie, R.J.; Xi, W.P.; Zhou, Z.Q. Metabolite profiles of essential oils in citrus peels and their taxonomic implications. Metabolomics 2015, 11, 952–963. [Google Scholar] [CrossRef]
- Zhang, H.P.; Xie, Y.X.; Liu, C.H.; Chen, S.L.; Hu, S.S.; Xie, Z.Z.; Deng, X.X.; Xu, J. Comprehensive comparative analysis of volatile compounds in citrus fruits of different species. Food Chem. 2017, 230, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Hopper, D.W.; Ghan, R.; Schlauch, K.A.; Cramer, G.R. Transcriptomic network analyses of leaf dehydration responses identify highly connected ABA and ethylene signaling hubs in three grapevine species differing in drought tolerance. BMC Plant Biol. 2016, 16, 118. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; Hotta, C.T.; Poelking, V.G.D.C.; Leite, D.C.C.; Buckeridge, M.S.; Loureiro, M.E.; Barbosa, M.H.P.; Carneiro, M.S.; Souza, G.M. Co-expression network analysis reveals transcription factors associated to cell wall biosynthesis in sugarcane. Plant Mol. Biol. 2016, 91, 15–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, Y.L.; Liu, C.; Yu, S.W.; Yang, H.; Sun, J.M.; Guo, C.X.; Huang, B.; Liu, Z.Y.; Yuan, Y.; Xia, E.H.; et al. Gene co-expression network analysis reveals coordinated regulation of three characteristic secondary biosynthetic pathways in tea plant (Camellia sinensis). BMC Genom. 2018, 19, 616. [Google Scholar] [CrossRef] [PubMed]
- Steingass, C.B.; Jutzi, M.; Müller, J.; Carle, R.; Schmarr, H.G. Ripening-dependent metabolic changes in the volatiles of pineapple (Ananas comosus (L.) Merr.) fruit: II. Multivariate statistical profiling of pineapple aroma compounds based on comprehensive two-dimensional gas chromatography-mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 2609–2624. [Google Scholar] [CrossRef] [PubMed]
- Vandendriessche, T.; Keulemans, J.; Geeraerd, A.; Nicolai, B.M.; Hertog, M.L.A.T.M. Evaluation of fast volatile analysis for detection of Botrytis cinerea infections in strawberry. Food Microbiol. 2012, 32, 406–414. [Google Scholar] [CrossRef]
- Song, A.A.L.; Abdullah, J.O.; Abdullah, M.P.; Shafee, N.; Othman, R.; Tan, E.F.; Noor, N.M.; Raha, A.R. Overexpressing 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGR) in the lactococcal mevalonate pathway for heterologous plant sesquiterpene production. PloS ONE 2012, 7, e52444. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; D’Auria, J.C.; Farooq, A.; Pichersky, E.; Gershenzo, J. Biosynthesis and emission of terpenoid volatiles from Arabidopsis flowers. Plant Cell 2003, 15, 481–494. [Google Scholar] [CrossRef]
- Flamini, G.; Tebano, M.; Cioni, P.L. Volatiles emission patterns of different plant organs and pollen of Citrus limon. Anal. Chim. Acta 2007, 589, 120–124. [Google Scholar] [CrossRef]
- Azam, M.; Jiang, Q.; Zhang, B.; Xu, C.J.; Chen, K.S. Citrus leaf volatiles as affected by developmental stage and genetic type. Int. J. Mol. Sci. 2013, 14, 17744–17766. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.; Zhang, B.; Xu, Y.Y.; Xu, C.J.; Chen, K.S. Comparative analysis of flower volatiles from nine Citrus at three blooming stages. Int. J. Mol. Sci. 2013, 14, 22346–22367. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wu, B.P.; Gao, L.X.; Liu, H.R.; Zhang, B.; Sun, C.D.; Chen, K.S. Glycosidically bound volatiles as affected by ripening stages of Satsuma mandarin fruit. Food Chem. 2018, 240, 1097–1105. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Tian, P.; Zhang, F.X.; Qin, H.; Miao, H.; Chen, Q.W.; Hu, Z.Y.; Cao, L.; Wang, M.J.; Gu, X.F.; et al. Integrative analyses of non-targeted volatile profiling and transcriptome data provide molecular insight into VOC diversity in cucumber plants (Cucumis sativus). Plant Physiol. 2016, 172, 603–618. [Google Scholar] [CrossRef] [PubMed]
- Goossens, J.; Mertens, J.; Goossens, A. Role and functioning of bHLH transcription factors in jasmonate signalling. J. Exp. Bot. 2017, 68, 1333–1347. [Google Scholar] [CrossRef] [PubMed]
- Matías-Hernández, L.; Jiang, W.M.; Yang, K.; Tang, K.X.; Brodelius, P.E.; Pelaz, S. AaMYB1, and its orthologue AtMYB61, affect terpene metabolism and trichome development in Artemisia annua and Arabidopsis thaliana. Plant J. 2017, 90, 520–534. [Google Scholar] [CrossRef] [PubMed]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.X. WRKY transcription factors. Trends Plant Sci. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Tholl, D. Terpene synthases and the regulation, diversity and biological roles of terpene metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 22, 1870–1874. [Google Scholar] [CrossRef]
- Yip, A.M.; Horvath, S. Gene network interconnectedness and the generalized topological overlap measure. BMC Bioinform. 2007, 8, 22. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | FC (F2/F1) | DEG | FC (L2/L1) | DEG | FC (S6/S1) | DEG |
|---|---|---|---|---|---|---|---|
| MEP Pathway | |||||||
| Cm152440 | DXS | 1.21 | 0.49 | yes, down | 0.20 | yes, down | |
| Cm165980 | DXS | 0.97 | 1.24 | 0.12 | yes, down | ||
| Cm165990 | DXS | 1.42 | 2.03 | 1.34 | |||
| Cm219000 | DXS | 1.09 | 1.72 | 1.32 | |||
| Cm104830 | DXS | 0.88 | 2.08 | yes, up | 1.49 | ||
| Cm018790 | DXR | 0.95 | 1.25 | 0.93 | |||
| Cm018780 | DXR | 1.43 | 0.86 | 1.22 | |||
| Cm191510 | MCT | 1.39 | 0.61 | 0.90 | |||
| Cm162030 | CMK | 1.34 | 0.88 | 0.91 | |||
| Cm051360 | MDS | 0.88 | 1.60 | 2.53 | yes, up | ||
| Cm131750 | HDS | 1.20 | 2.67 | yes, up | 1.52 | ||
| Cm315300 | HDR | 0.31 | - | - | |||
| Cm315660 | HDR | 0.24 | - | - | |||
| Cm219660 | HDR | 0.99 | 0.80 | 0.42 | yes, down | ||
| Cm276630 | HDR | 1.41 | 2.10 | yes, up | 0.93 | ||
| Cm318240 | HDR | 1.05 | - | 0.61 | |||
| MVA Pathway | |||||||
| Cm183050 | AACT | 0.92 | 1.03 | 1.28 | |||
| Cm042040 | AACT | 1.04 | 0.60 | 2.90 | yes, up | ||
| Cm014300 | HMGS | 1.17 | 0.82 | 1.24 | |||
| Cm248930 | HMGS | 0.98 | 0.38 | yes, down | 1.47 | ||
| Cm197870 | HMGR | 0.93 | 0.91 | 1.42 | |||
| Cm212470 | HMGR | 0.72 | 0.86 | 0.66 | |||
| Cm122170 | HMGR | 1.56 | 0.33 | yes, down | 5.51 | yes, up | |
| Cm268880 | MVK | 1.23 | 0.97 | 0.55 | |||
| Cm034040 | PMK | 1.10 | 0.52 | 1.08 | |||
| Cm235330 | MVD | 1.13 | 0.55 | 0.65 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Zhu, C.; Xu, C.; Sun, J.; Grierson, D.; Zhang, B.; Chen, K. Integration of Metabolite Profiling and Transcriptome Analysis Reveals Genes Related to Volatile Terpenoid Metabolism in Finger Citron (C. medica var. sarcodactylis). Molecules 2019, 24, 2564. https://doi.org/10.3390/molecules24142564
Xu Y, Zhu C, Xu C, Sun J, Grierson D, Zhang B, Chen K. Integration of Metabolite Profiling and Transcriptome Analysis Reveals Genes Related to Volatile Terpenoid Metabolism in Finger Citron (C. medica var. sarcodactylis). Molecules. 2019; 24(14):2564. https://doi.org/10.3390/molecules24142564
Chicago/Turabian StyleXu, Yaying, Changqing Zhu, Changjie Xu, Jun Sun, Donald Grierson, Bo Zhang, and Kunsong Chen. 2019. "Integration of Metabolite Profiling and Transcriptome Analysis Reveals Genes Related to Volatile Terpenoid Metabolism in Finger Citron (C. medica var. sarcodactylis)" Molecules 24, no. 14: 2564. https://doi.org/10.3390/molecules24142564