
Antifungal Activity of Essential Oil Compounds (Geraniol and Citral) and Inhibitory Mechanisms on Grain Pathogens (Aspergillus flavus and Aspergillus ochraceus)



Abstract
:1. Introduction
2. Results
2.1. Inhibitory Effects of Geraniol and Citral on Toxigenic Fungi
2.2. Influence of EOC Treatments on Grain Quality Parameters
2.3. Scanning Electron Microscopy (SEM) Analysis
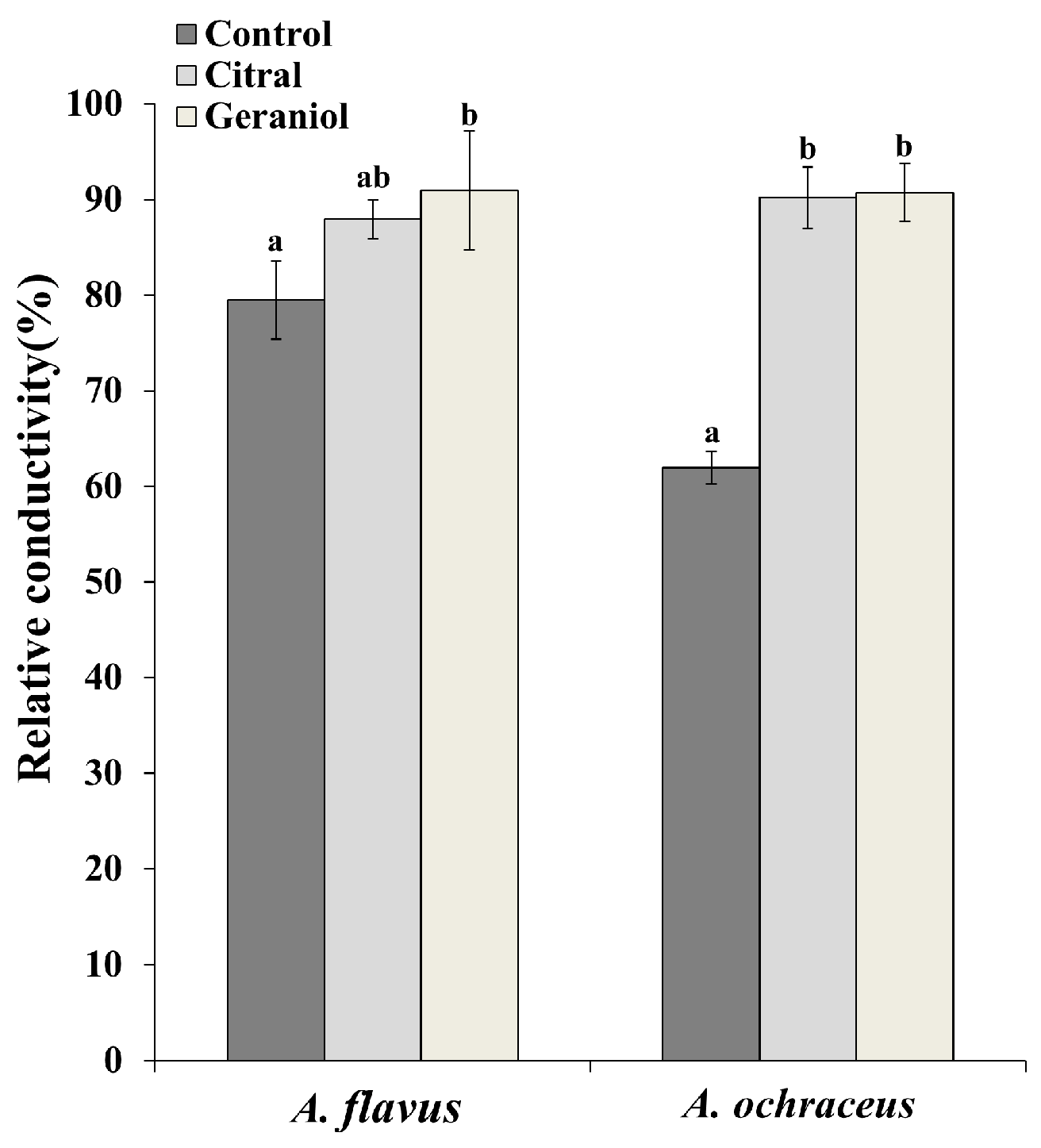
2.4. Determination of Cell Membrane Permeability
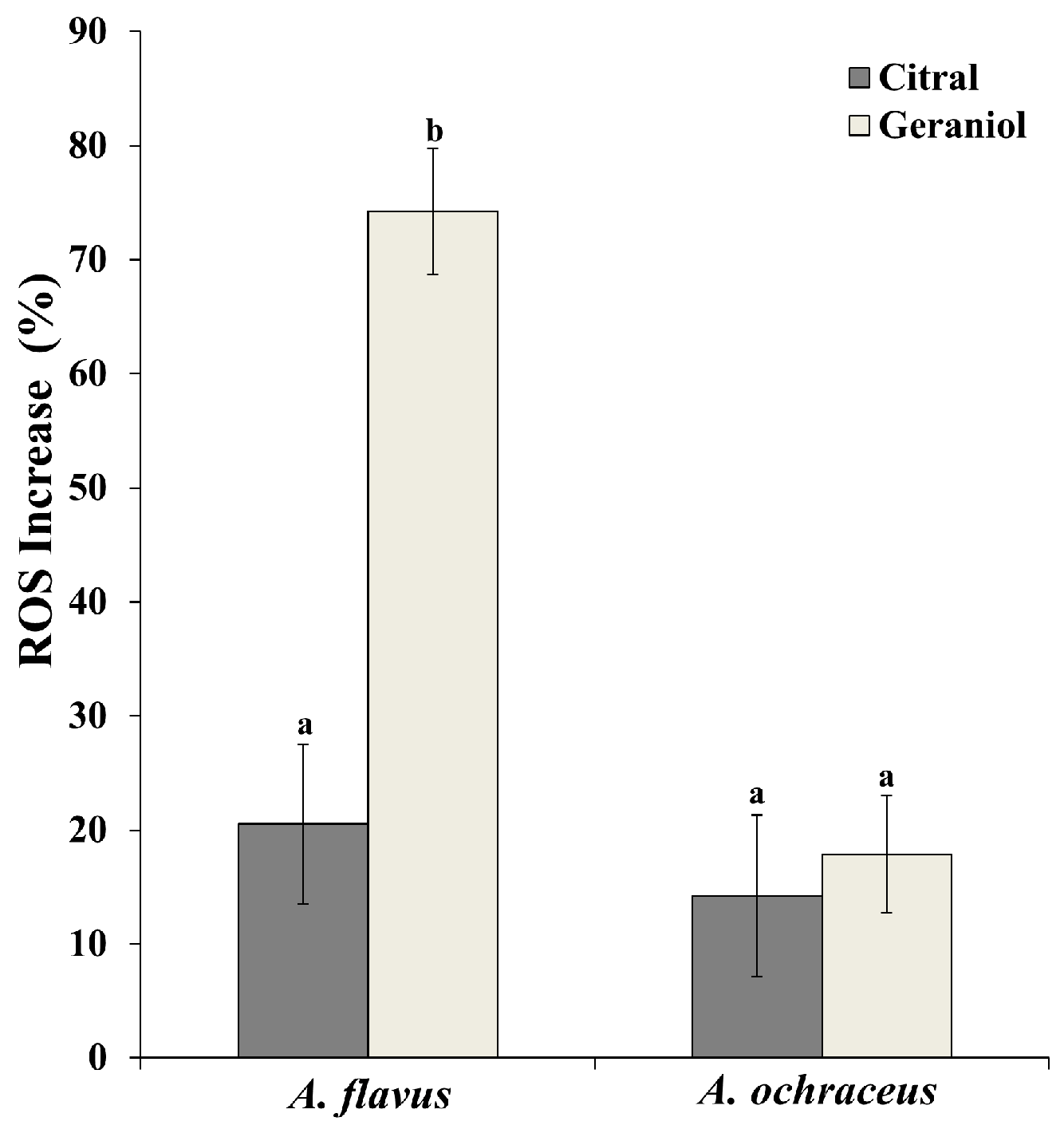
2.5. Effect of EOCs on Cellular ROS Increase
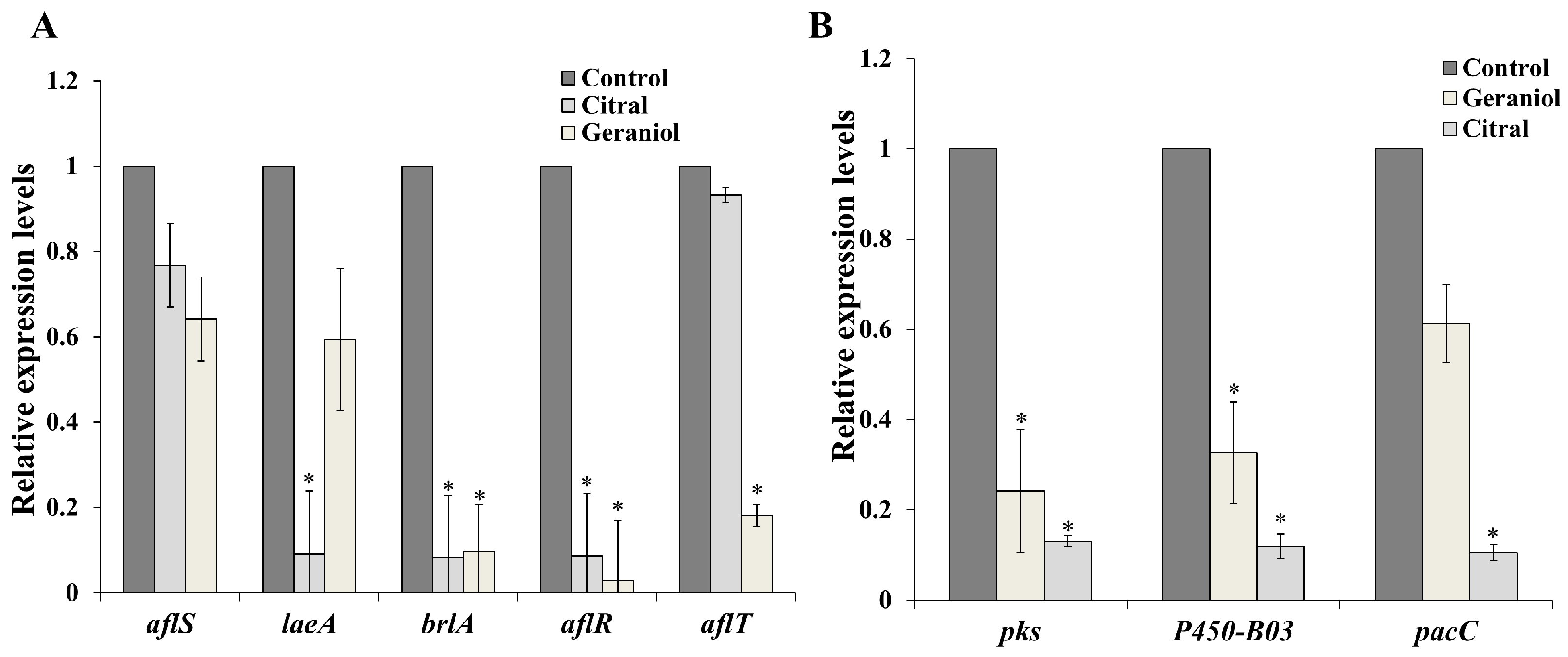
2.6. Gene Expression Analysis
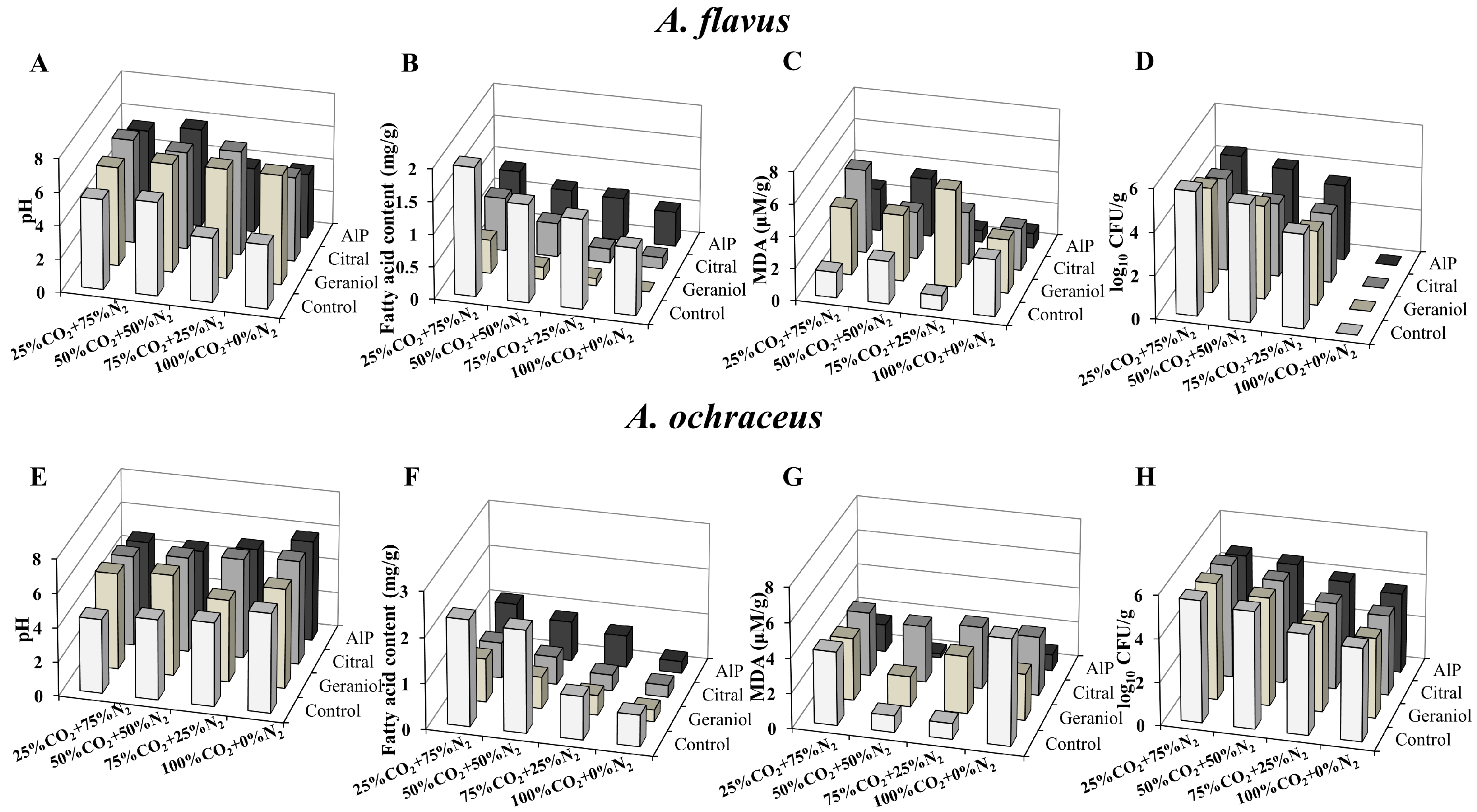
2.7. Cotreatment with EOCs and MAP for Cereal Preservation
3. Discussion
4. Materials and Methods
4.1. Chemical and Reagents
4.2. Strains and Culture Conditions
4.3. In Vitro Antifungal Activity Assay
4.4. In Situ Investigation with EOC Fumigation against Pathogens of Grains
4.5. Determination of pH, Fatty Acid Content, MDA Content, and Colony Number
4.6. SEM Analysis
4.7. Permeability of the Toxigenic Cell Membrane Assay
4.8. Determination of ROS in Pathogens
4.9. Isolation of Total RNA and cDNA Synthesis
4.10. Quantification of Gene Expression by qRT-PCR
4.11. Synergistic Effect of EOCs Combined with MAP against Pathogens on Grains
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Reddy, B.N.; Raghavender, C.R. Outbreaks of aflatoxicoses in India. Afr. J. Food Agric. Nutr. Dev. 2007, 7, 1–15. [Google Scholar]
- Candlish, A.A.G.; Pearson, S.M.; Aidoo, K.E.; Smith, J.E.; Kelly, B.; Irvine, H. A survey of ethnic foods for microbial quality and content aflatoxin. Food Addit. Contam. 2001, 18, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Bassolé, I.H.N.; Juliani, H.R. Essential oils in combination and their antimicrobial properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Carson, C.F.; Mee, B.J.; Riley, T.V. Mechanism of action of Melaleuca alternifolia (tea tree) oil on Staphylococcus aureus determined by time-kill, lysis, leakage, and salt tolerance assays and electron microscopy. Antimicrob. Agents Chemother. 2002, 46, 1914–1920. [Google Scholar] [CrossRef] [PubMed]
- Richter, C.; Schlegel, J. Mitochondrial calcium release induced by prooxidants. Toxicol. Lett. 1993, 67, 119–127. [Google Scholar] [CrossRef]
- Yoon, H.S.; Moon, S.C.; Kim, N.D.; Park, B.S.; Jeong, M.H.; Yoo, Y.H. Genistein induces apoptosis of RPE-J cells by opening mitochondrial PTP. Biochem. Biophys. Res. Commun. 2000, 276, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Jayas, D.S.; Jeyamkondan, S. pH-Postharvest technology. Biosyst. Eng. 2002, 82, 235–251. [Google Scholar] [CrossRef]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH oxidases: An overview from structure to innate immunity-associated pathologies. Cell. Mol. Immunol. 2014, 12, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Tang, Z.; Tan, Y.; Zhang, Y.; Luo, B.; Yang, M.; Lian, X.; Shen, Q.; Miller, A.J.; Xu, G. Overexpression of a pH-sensitive nitrate transporter in rice increases crop yields. Proc. Natl. Acad. Sci. USA 2016, 113, 7118–7123. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Juven, B.J.; Kanner, J.; Schved, F.; Weisslowicz, H. Factors that interact with the antibacterial action of thyme essential oil and its active constituents. J. Appl. Bacteriol. 1994, 76, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Fourar-Belaifa, R.; Fleurat-Lessard, F.; Bouznad, Z. A systemic approach to qualitative changes in the stored-wheat ecosystem: Prediction of deterioration risks in unsafe storage conditions in relation to relative humidity level, infestation by Sitophilus oryzae (L.), and wheat variety. J. Stored Prod. Res. 2011, 47, 48–61. [Google Scholar] [CrossRef]
- Rodrigues, S.M.; Andrade, M.O.; Gomes, A.P.; Damatta, F.M.; Baracat-Pereira, M.C.; Fontes, E.P. Arabidopsis and tobacco plants ectopically expressing the soybean antiquitin-like ALDH7 gene display enhanced tolerance to drought, salinity, and oxidative stress. J. Exp. Bot. 2006, 57, 1909–1918. [Google Scholar] [CrossRef] [PubMed]
- Doran, P.J.; Briggs, D.E. Microbes and grain germination. J. Inst. Brew. 1993, 99, 165–170. [Google Scholar] [CrossRef]
- Lahouar, A.; Marin, S.; Crespo-Sempere, A.; Saïd, S.; Sanchis, V. Influence of temperature, water activity and incubation time on fungal growth and production of ochratoxin A and zearalenone by toxigenic Aspergillus tubingensis and Fusarium incarnatum isolates in sorghum seeds. Int. J. Food Microbiol. 2017, 242, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Slump, R.A.; Steging, G.; Smid, E.J. Antimicrobial activity of carvacrol toward Bacillus cereus on rice. J. Food Prot. 2000, 63, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.; Chen, X.G.; Liu, C.S.; Liu, C.G.; Meng, X.H.; Yu, L.J. Antibacterial mechanism of chitosan microspheres in a solid dispersing system against E. coli. Colloids Surf. B Biointerfaces 2008, 65, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Ginsburg, I.; van Heerden, P.V.; Koren, E. From amino acids polymers, antimicrobial peptides, and histones, to their possible role in the pathogenesis of septic shock: A historical perspective. J. Inflamm. Res. 2017, 10, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef] [PubMed]
- Chueca, B.; Pagán, R.; García-Gonzalo, D. Oxygenated monoterpenes citral and carvacrol cause oxidative damage in Escherichia coli without the involvement of tricarboxylic acid cycle and fenton reaction. Int. J. Food Microbiol. 2014, 189, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Hajyehia, A.; Levischaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Chueca, B.; Pagán, R.; García-Gonzalo, D. Differential mechanism of Escherichia coli inactivation by (+)-limonene as a function of cell physiological state and drug’s concentration. PLoS ONE 2014, 9, e94072. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Shang, B.; Wang, L.; Lu, Z.; Liu, Y. Cinnamaldehyde inhibits fungal growth and aflatoxin b1 biosynthesis by modulating the oxidative stress response of Aspergillus flavus. Appl. Microbiol. Biotechnol. 2015, 100, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Navarro, J.; Greve, R.A.; Adams, T.H. Translational repression of brlA expression prevents premature development in Aspergillus. EMBO J. 1993, 12, 2449. [Google Scholar] [PubMed]
- Chang, P.K. The Aspergillus parasiticus protein AFLJ interacts with the aflatoxin pathway-specific regulator AFLR. Mol. Genet. Genom. 2003, 268, 711–719. [Google Scholar]
- Kong, Q.; Chi, C.; Yu, J.; Shan, S.; Li, Q.; Li, Q.; Guan, B.; Nierman, W.C.; Bennett, J.W. The inhibitory effect of Bacillus megaterium on aflatoxin and cyclopiazonic acid biosynthetic pathway gene expression in Aspergillus flavus. Appl. Microbiol. Biotechnol. 2014, 98, 5161–5172. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Xing, F.; Selvaraj, J.N.; Liu, X.; Wang, L.; Hua, H.; Zhou, L.; Zhao, Y.; Wang, Y.; Liu, Y. Inhibitory effect of cinnamaldehyde, citral, and eugenol on aflatoxin biosynthetic gene expression and aflatoxin b1 biosynthesis in Aspergillus flavus. J. Food Sci. 2015, 80, M2917–M2924. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, F.; Wang, L.; Wang, Q.; Selvaraj, J.N.; Zhao, Y.; Wang, Y.; Xing, F.; Liu, Y. pH-Signaling transcription factor AopacC regulates ochratoxin A biosynthesis in Aspergillus ochraceus. J. Agric. Food Chem. 2018, 66, 4394–4401. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, M. IARC monographs on the evaluation of carcinogenic risks to humans: Some traditional herbal medicines, some mycotoxins, naphthalene and styrene. J. Ethnopharmacol. 2003, 88, 299–300. [Google Scholar] [CrossRef]
- Gallo, A.; Perrone, G.; Solfrizzo, M.; Epifani, F.; Abbas, A.; Dobson, A.; Mule, G. Characterisation of a pks gene which is expressed during ochratoxin a production by Aspergillus carbonarius. Int. J. Food Microbiol. 2009, 129, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Patiño, B.; Cortés, L.; González-Jaén, M.T.; Vázquez, C. Mechanisms involved in reduction of ochratoxin a produced by Aspergillus westerdijkiae using Debaryomyces hansenii cyc 1244. Int. J. Food Microbiol. 2011, 151, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Lappa, I.K.; Simini, E.; Nychas, G.-J.E.; Panagou, E.Z. In vitro evaluation of essential oils against Aspergillus carbonarius isolates and their effects on ochratoxin a related gene expression in synthetic grape medium. Food Control 2017, 73, 71–80. [Google Scholar] [CrossRef]
- Espina, L.; Somolinos, M.; Lorán, S.; Conchello, P.; García, D.; Pagán, R. Chemical composition of commercial citrus fruit essential oils and evaluation of their antimicrobial activity acting alone or in combined processes. Food Control 2011, 22, 896–902. [Google Scholar] [CrossRef]
- Garcia, D.; Garcia, C.E.; Ramos, A.J.; Sanchis, V.; Marín, S. Mould growth and mycotoxin production as affected by Equisetum arvense and Stevia rebaudiana extracts. Food Control 2011, 22, 1378–1384. [Google Scholar] [CrossRef]
- Bard, M.; Albrecht, M.R.; Gupta, N.; Guynn, C.J.; Stillwell, W. Geraniol interferes with membrane functions in strains of Candida and Saccharomyces. Lipids 1988, 23, 534–538. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Fatima, Z.; Hameed, S. Insights into the mode of action of anticandidal herbal monoterpenoid geraniol reveal disruption of multiple MDR mechanisms and virulence attributes in Candida albicans. Arch. Microbiol. 2016, 198, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Giorni, P.; Battilani, P.; Pietri, A.; Magan, N. Effect of aw and CO2 level on Aspergillus flavus growth and aflatoxin production in high moisture maize post-harvest. Int. J. Food Microbiol. 2008, 122, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Alfonzo, A.; Martorana, A.; Guarrasi, V.; Barbera, M.; Gaglio, R.; Santulli, A.; Settanni, L.; Galati, A.; Moschetti, G.; Francesca, N. Effect of the lemon essential oils on the safety and sensory quality of salted sardines (Sardina pilchardus Walbaum 1792). Food Control 2017, 73, 1265–1274. [Google Scholar] [CrossRef]
- Chaves-López, C.; Serio, A.; Gianotti, A.; Sacchetti, G.; Ndagijimana, M.; Ciccarone, C.; Stellarini, A.; Corsetti, A.; Paparella, A. Diversity of food-borne Bacillus volatile compounds and influence on fungal growth. J. Appl. Microbiol. 2015, 119, 487–499. [Google Scholar] [CrossRef] [PubMed]
- AACC. Approved Methods of American Association of Cereal Chemists, 10th ed.; American Association of Cereal Chemists Inc.: St. Paul, MN, USA, 2000. [Google Scholar]
- Demiral, T.; Turkan, I. Comparative lipid peroxidation, antioxidant defense systems and proline content in roots of two rice cultivars differing in salt tolerance. Environ. Exp. Bot. 2005, 53, 247–257. [Google Scholar] [CrossRef]
- Patra, J.K.; Baek, K.H. Antibacterial activity and action mechanism of the essential oil from Enteromorpha linza L. against foodborne pathogenic bacteria. Molecules 2016, 21, 388. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the essential oil compounds are available from the authors. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EOC Concentration | A. flavus | A. ochraceus | |||
|---|---|---|---|---|---|
| (μL/mL) | Growth (mm) 1 | Inhibition Rate (%) 1 | Growth (mm) 1 | Inhibition Rate (%) 1 | |
| Citral | 0 | 65.67 ± 3.01a | 0.00 ± 0.00a | 66.83 ± 1.26a | 0.00 ± 0.00a |
| 0.1 | 7.03 ± 0.58b | 96.64 ± 0.26b | 6.68 ± 0.29b | 97.28 ± 0.96b | |
| 0.2 | 6.33 ± 0.22b | 97.80 ± 0.37b | 6.00 ± 0.09bc | 98.38 ± 0.17c | |
| 0.3 | 6.00 ± 0.16b | 98.35 ± 0.18c | 5.28 ± 0.95cd | 99.54 ± 0.35c | |
| 0.4 | 5.52 ± 0.10b | 99.14 ± 0.48d | 5.00 ± 0.00d | 100 ± 0.00d | |
| 0.5 | 5.00 ± 0.00b | 100 ± 0.00b | 5.00 ± 0.00d | 100 ± 0.00e | |
| 0.6 | 5.00 ± 0.00b | 100 ± 0.00b | 5.00 ± 0.00d | 100 ± 0.00e | |
| Geraniol | 0 | 65.67 ± 3.01a | 0.00 ± 0.00a | 66.83 ± 1.26a | 0.00 ± 0.00a |
| 0.1 | 8.87 ± 0.12b | 93.62 ± 0.35b | 8.47 ± 0.58b | 94.39 ± 0.16b | |
| 0.2 | 7.03 ± 0.59bc | 96.64 ± 0.26c | 6.37 ± 0.12c | 97.79 ± 0.23c | |
| 0.3 | 6.00 ± 0.13c | 98.35 ± 0.38d | 6.00 ± 0.16c | 98.38 ± 0.13d | |
| 0.4 | 6.00 ± 0.03c | 98.35 ± 0.08d | 6.00 ± 0.32c | 98.38 ± 0.25d | |
| 0.5 | 5.95 ± 0.70c | 98.44 ± 0.52d | 6.00 ± 0.24c | 98.38 ± 0.06d | |
| 0.6 | 5.95 ± 0.35c | 98.44 ± 0.05d | 6.00 ± 0.15c | 98.38 ± 0.11d | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, X.; Shao, Y.-L.; Tang, Y.-J.; Zhou, W.-W. Antifungal Activity of Essential Oil Compounds (Geraniol and Citral) and Inhibitory Mechanisms on Grain Pathogens (Aspergillus flavus and Aspergillus ochraceus). Molecules 2018, 23, 2108. https://doi.org/10.3390/molecules23092108
Tang X, Shao Y-L, Tang Y-J, Zhou W-W. Antifungal Activity of Essential Oil Compounds (Geraniol and Citral) and Inhibitory Mechanisms on Grain Pathogens (Aspergillus flavus and Aspergillus ochraceus). Molecules. 2018; 23(9):2108. https://doi.org/10.3390/molecules23092108
Chicago/Turabian StyleTang, Xi, Ye-Lin Shao, Ya-Jie Tang, and Wen-Wen Zhou. 2018. "Antifungal Activity of Essential Oil Compounds (Geraniol and Citral) and Inhibitory Mechanisms on Grain Pathogens (Aspergillus flavus and Aspergillus ochraceus)" Molecules 23, no. 9: 2108. https://doi.org/10.3390/molecules23092108