Cyclodextrins: Assessing the Impact of Cavity Size, Occupancy, and Substitutions on Cytotoxicity and Cholesterol Homeostasis


Abstract
:
1. Introduction
2. Results and Discussion
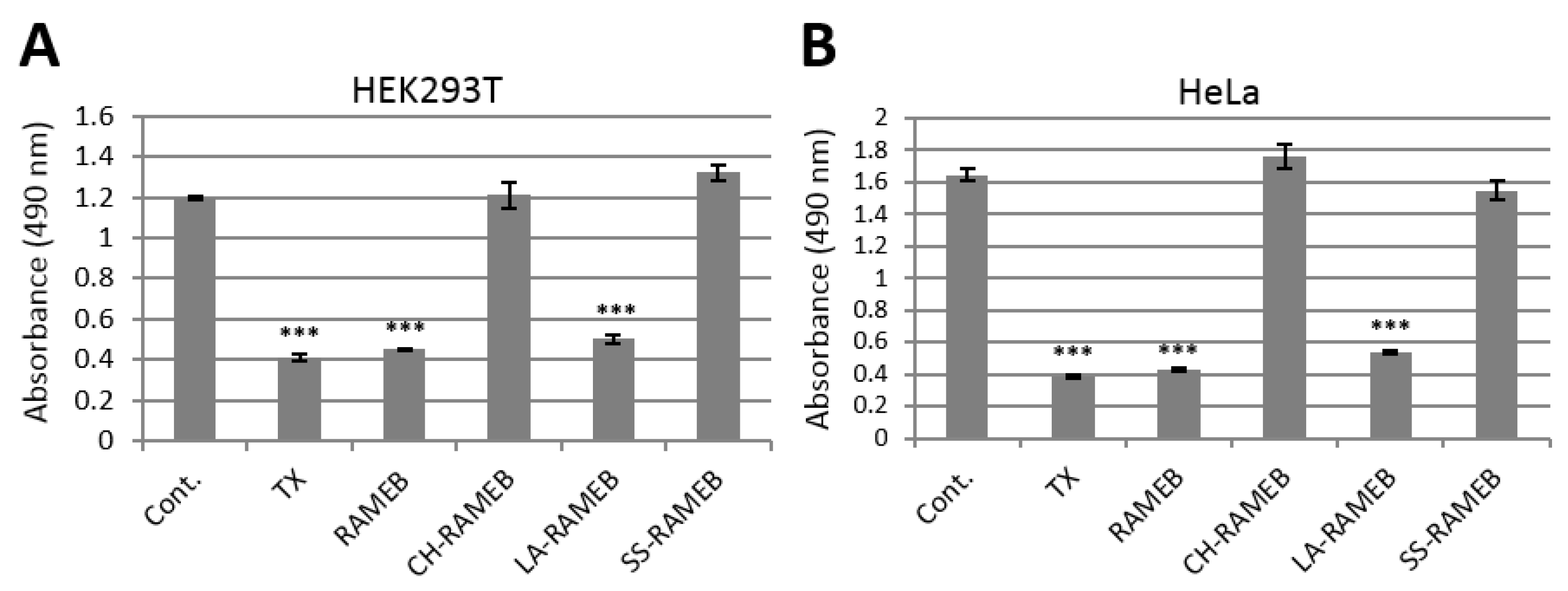
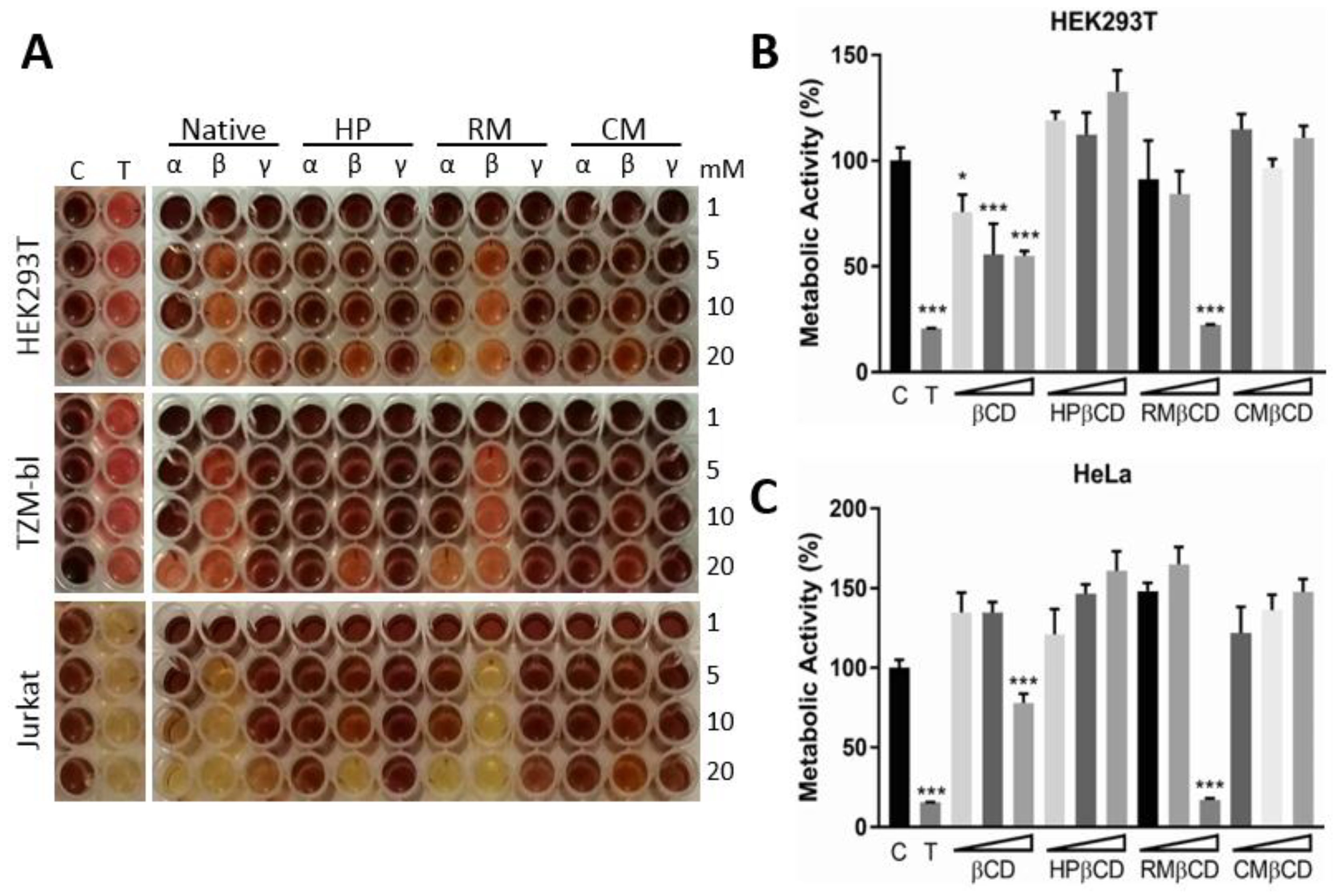
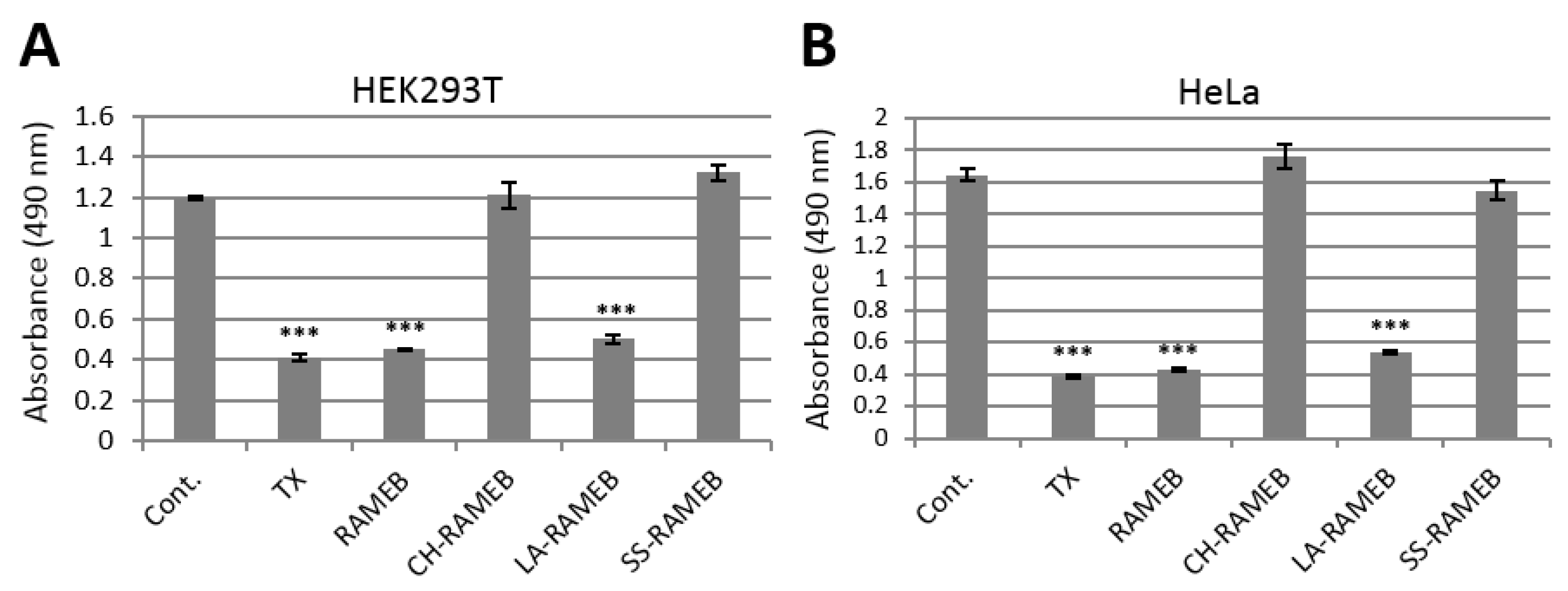
2.1. Cytotoxic Activity of CDs in Human Cell Lines
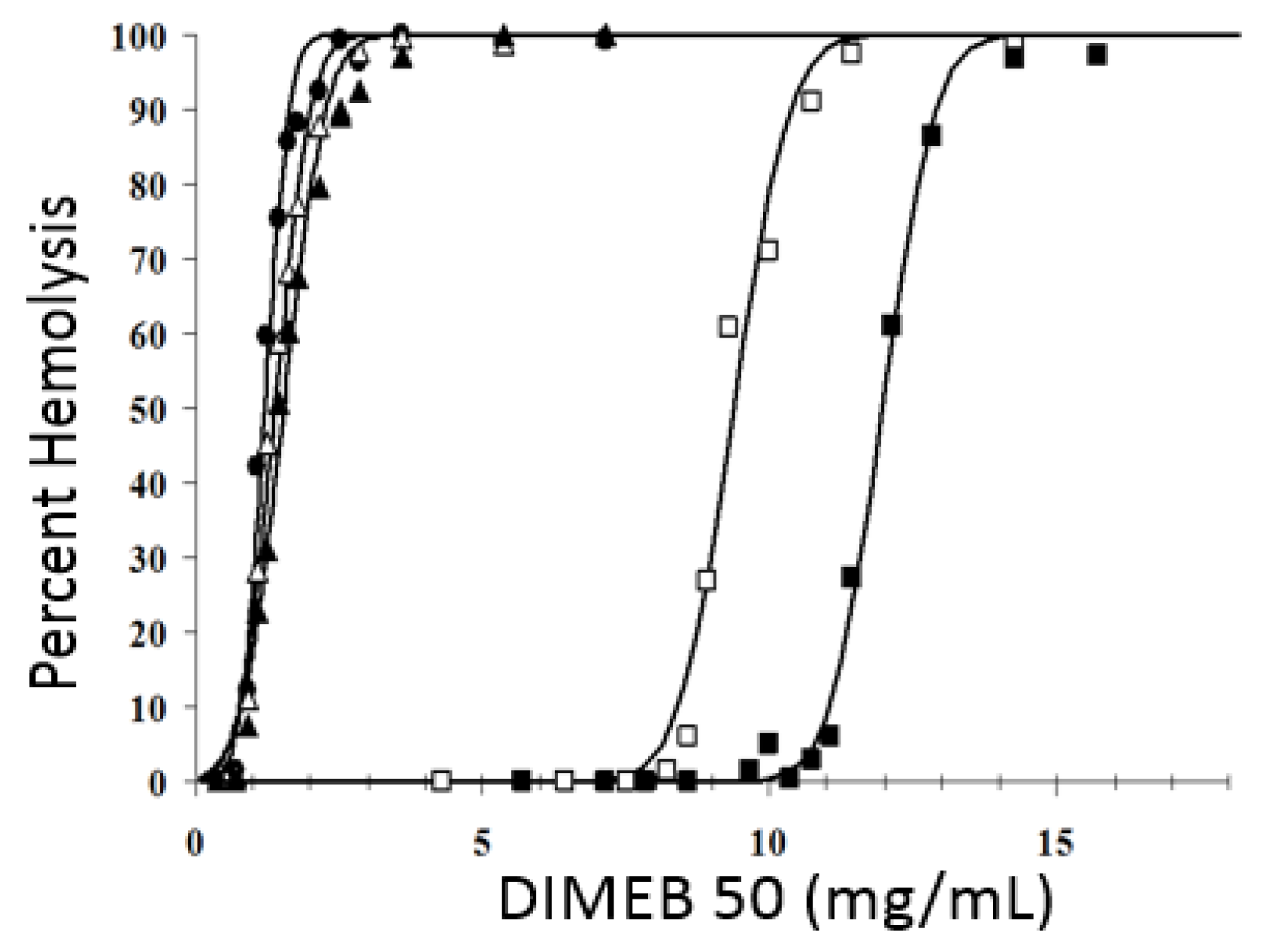
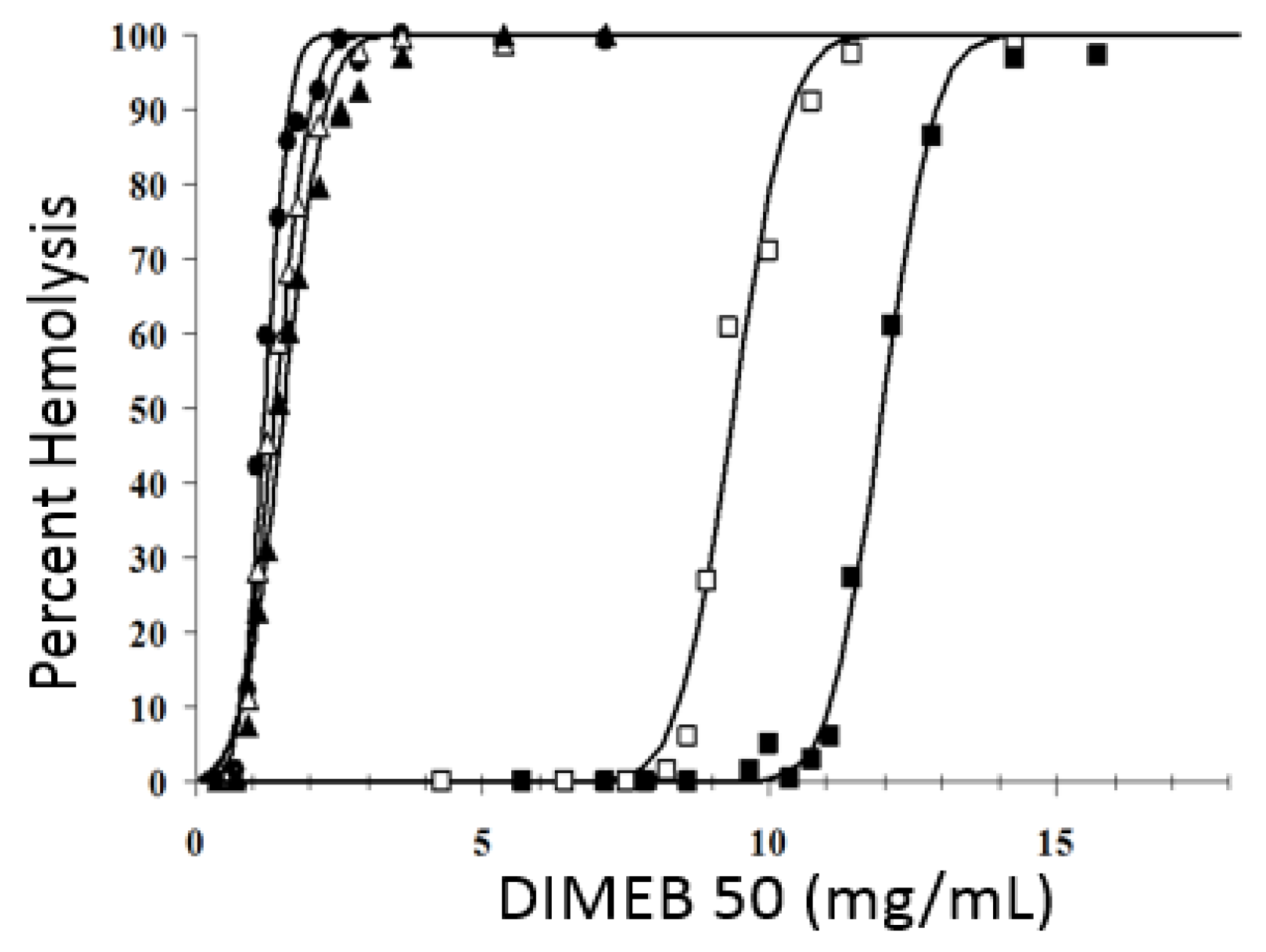
2.2. Hemolytic Activity of Methyl-β-CDs In Vitro
2.3. Impact of Blood Plasma Lipids on the Hemolytic Activity of Methyl-β-CDs
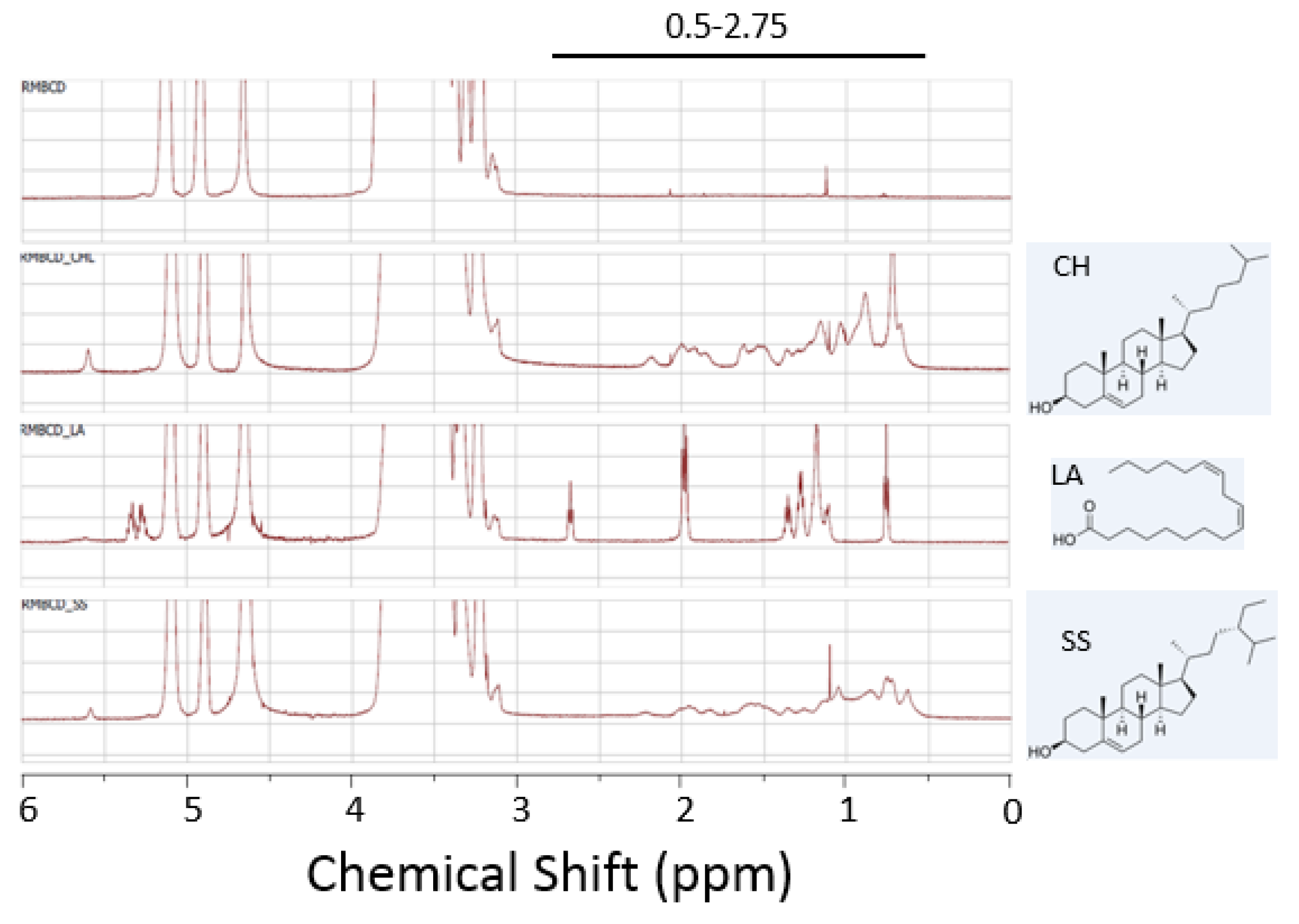
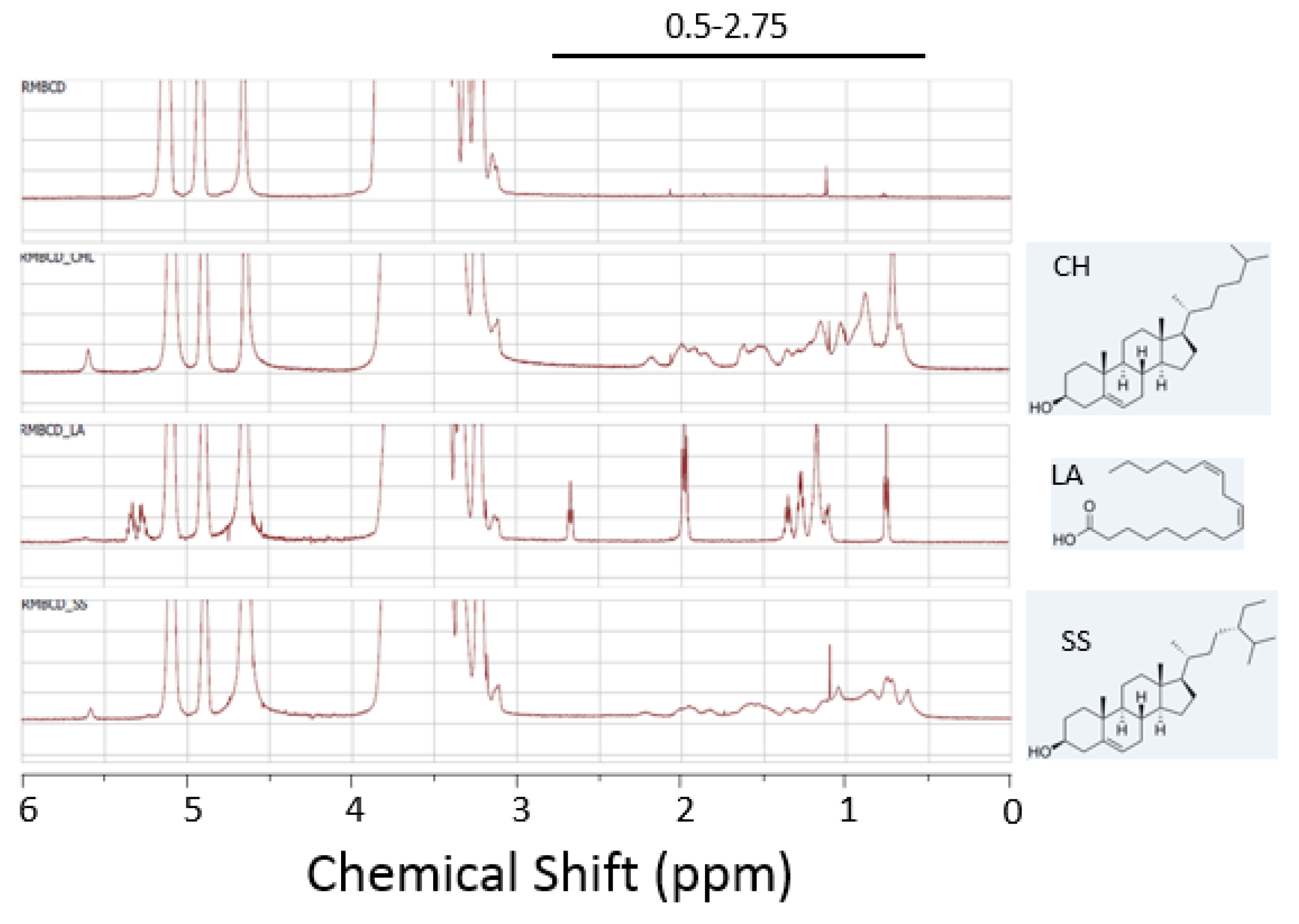
2.4. Abrogation of Cytotoxic Activity by Cholesterol Occupancy in the CD Cavity
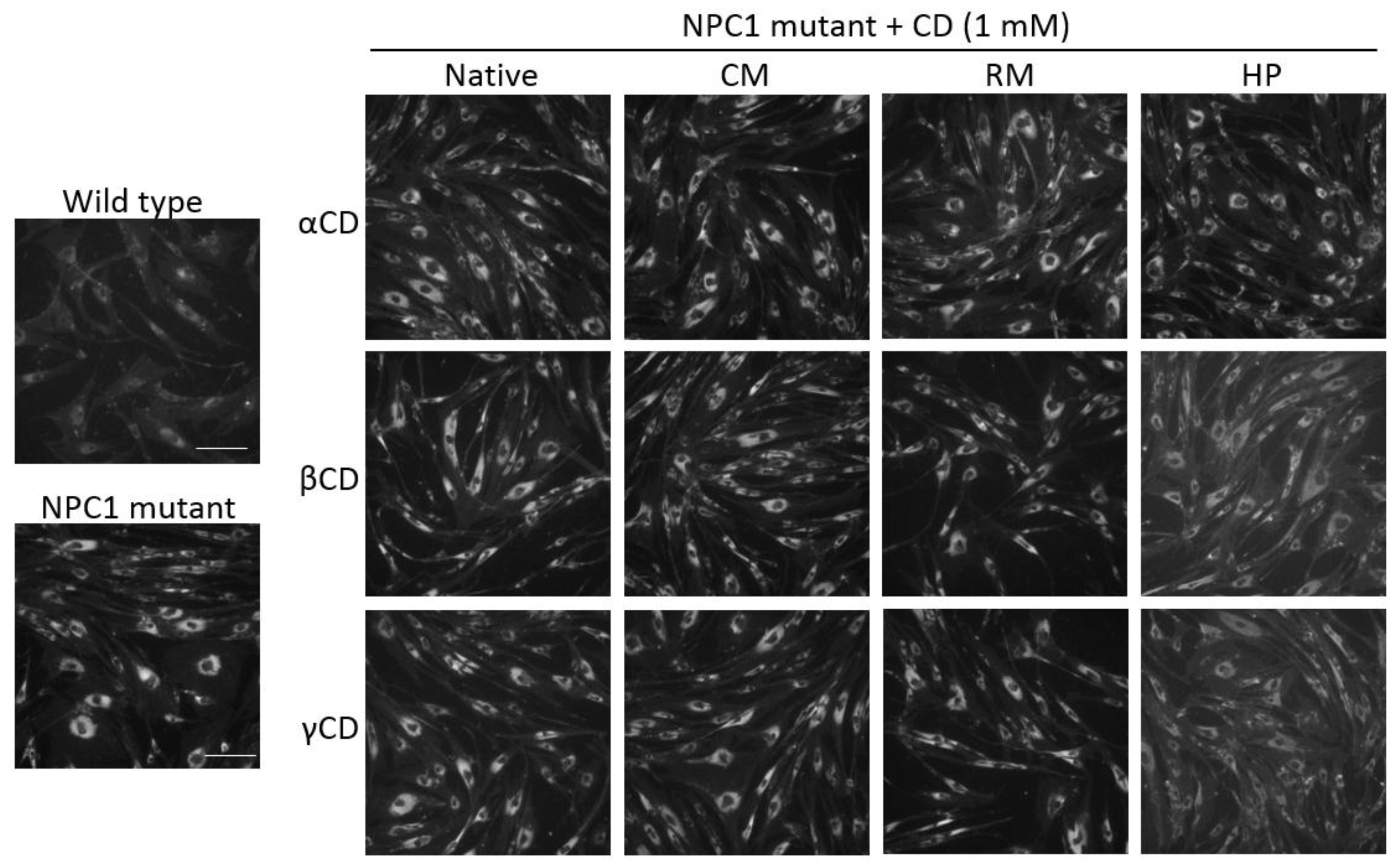
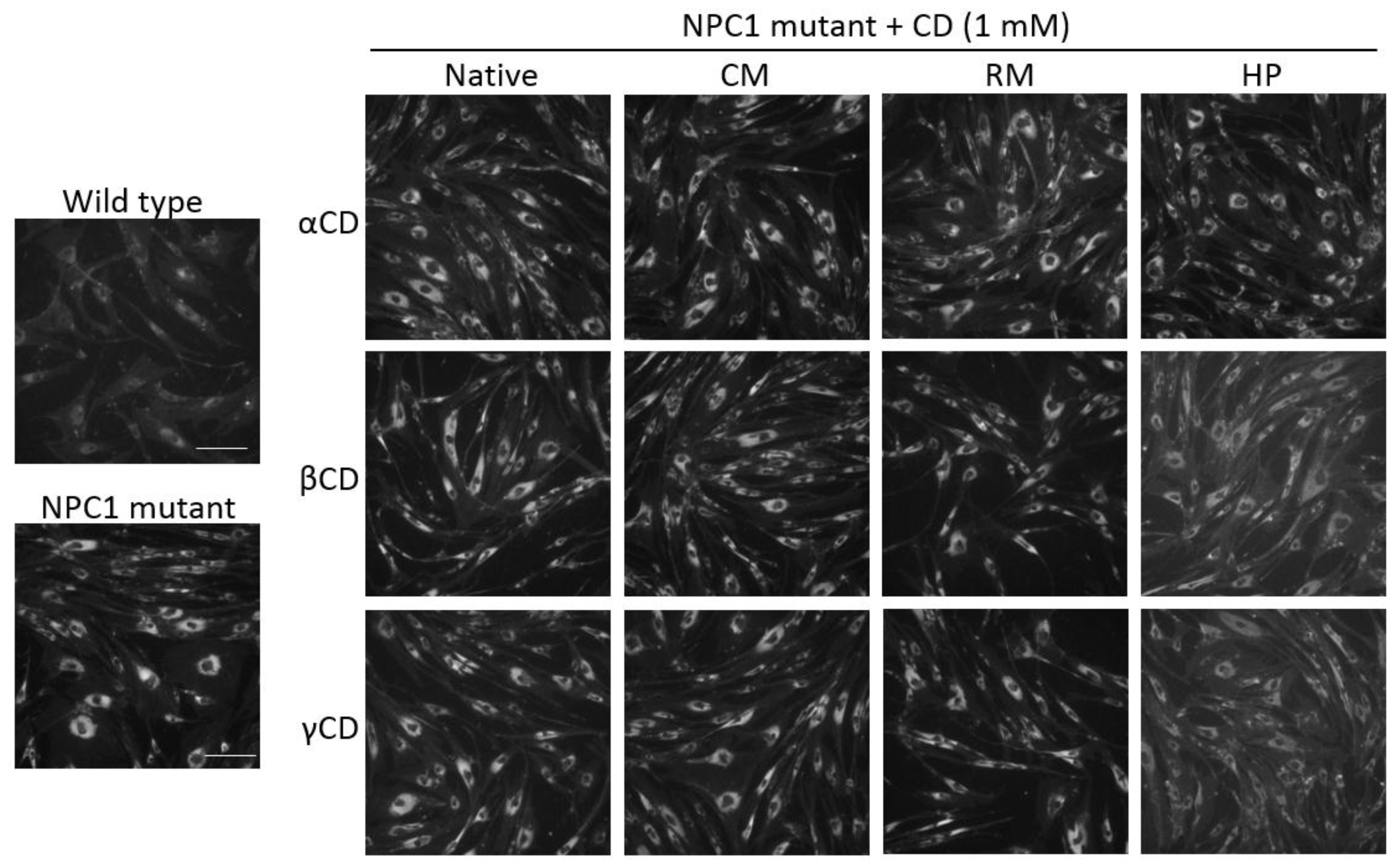
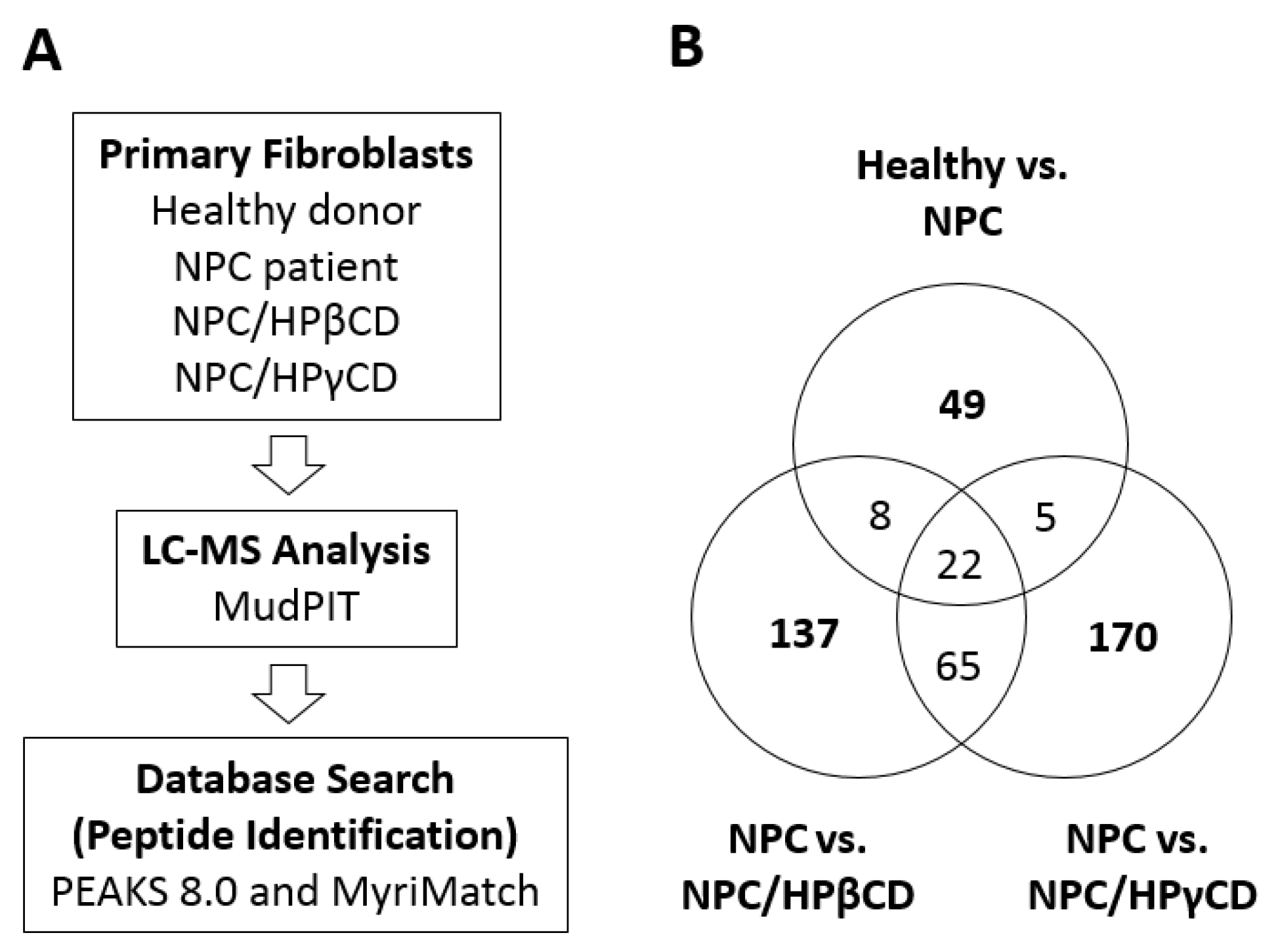
2.5. Effects of CDs on Intracellular Cholesterol Distribution
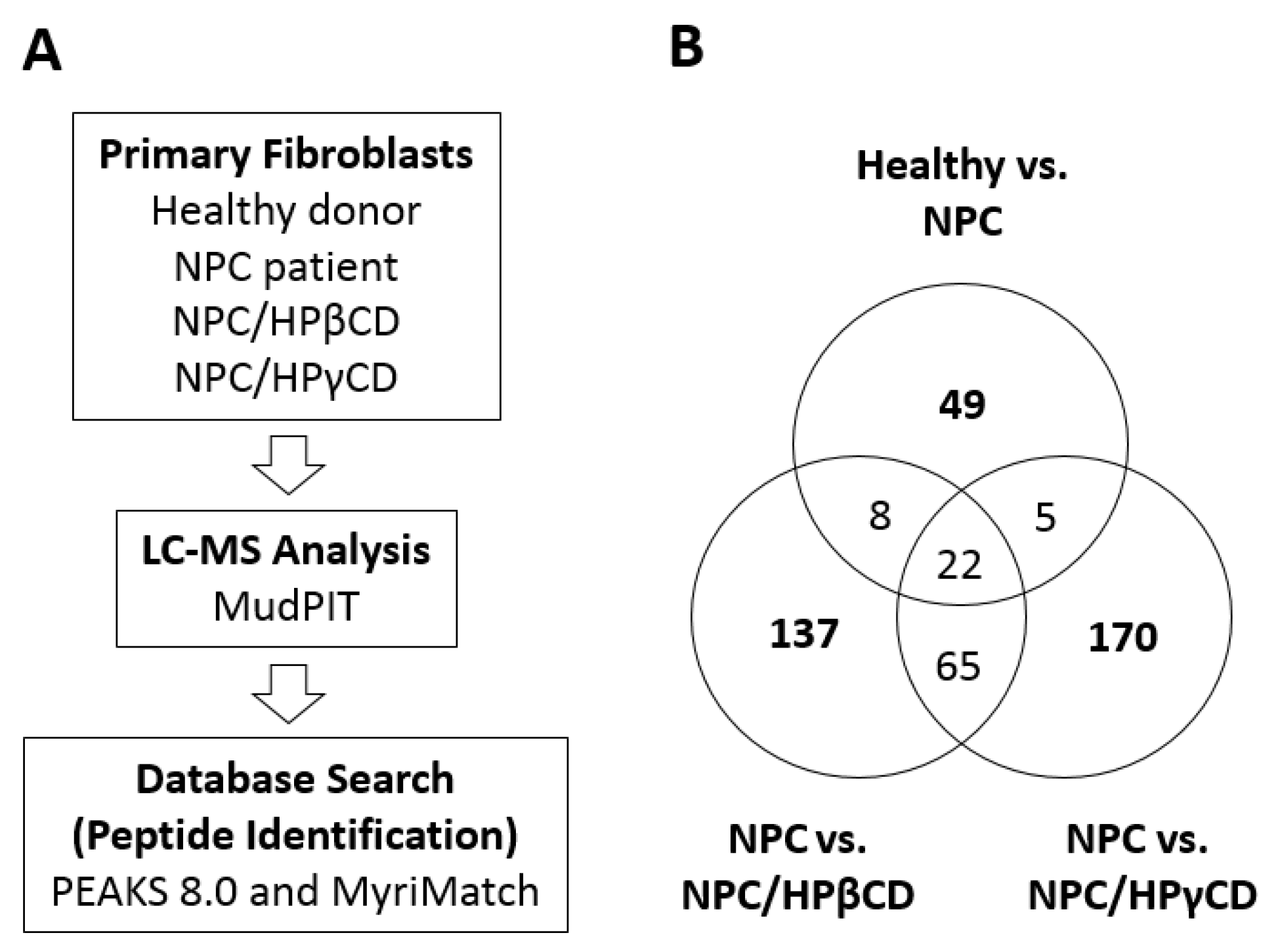
2.6. Effects of HPβCD and HPγCD Treatments on the Expression of Cellular Proteins
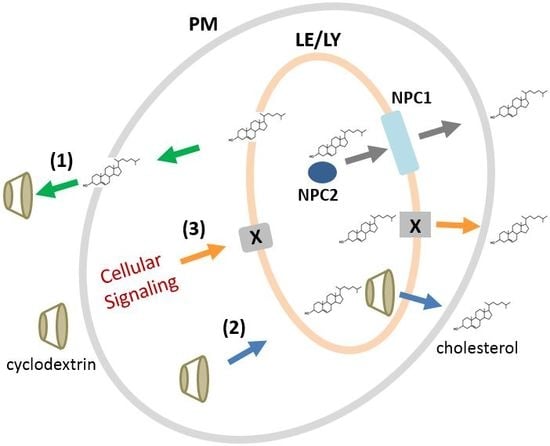
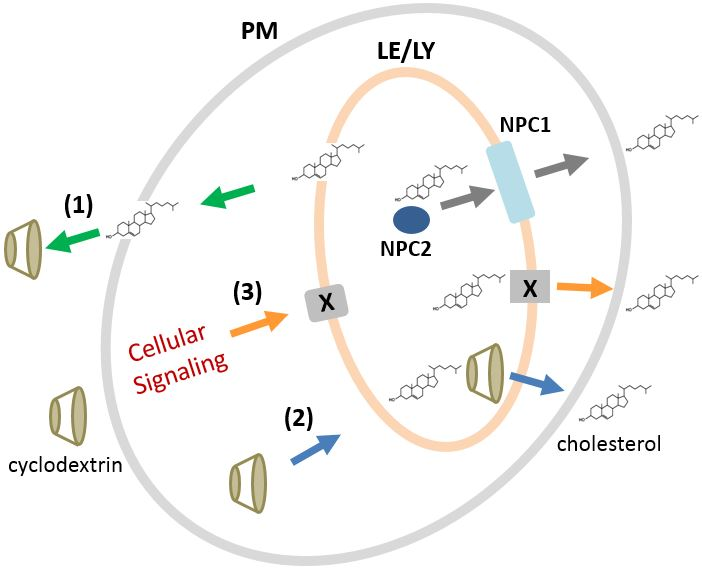
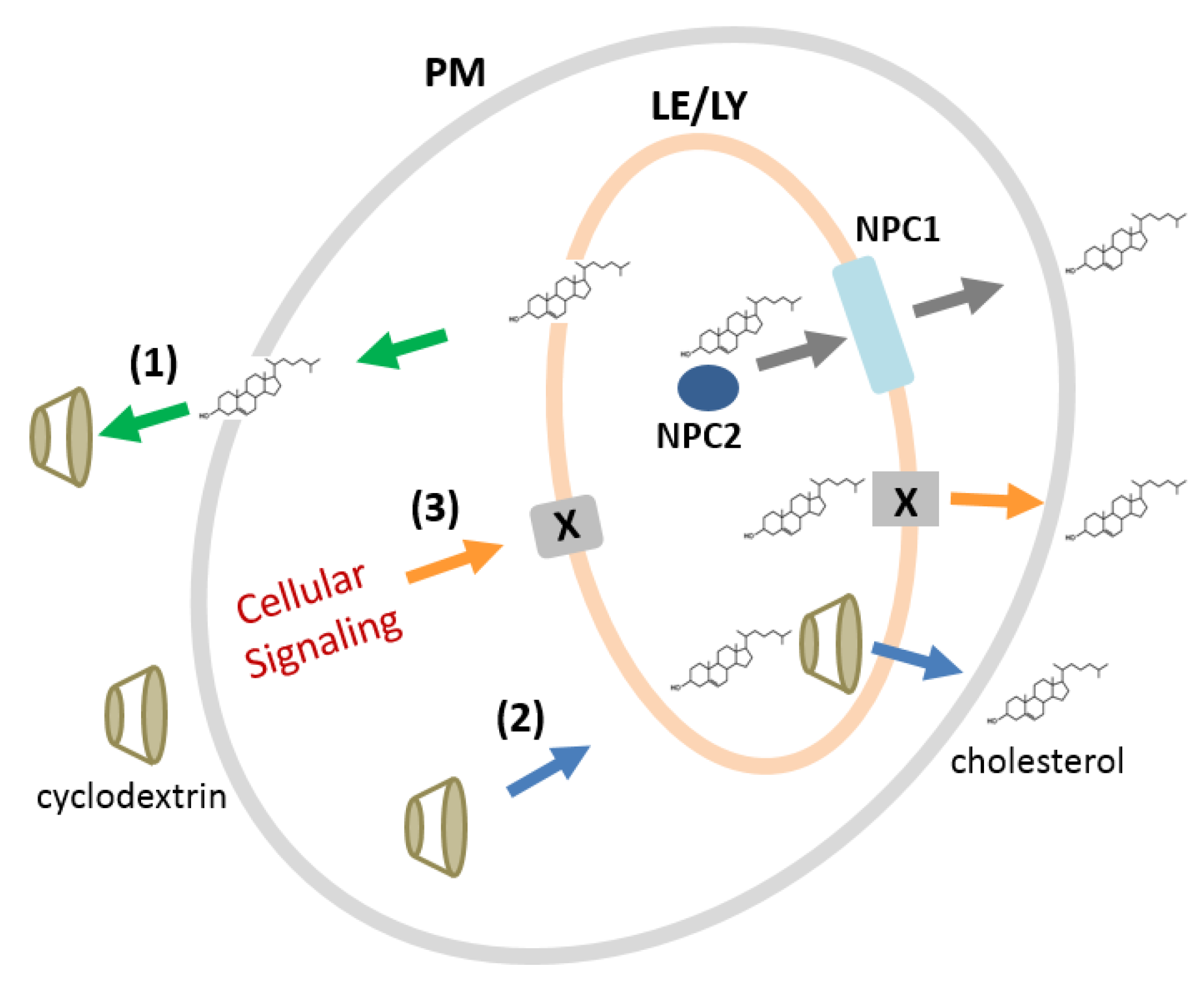
2.7. Mechanisms of CDs for Modulating Intracellular Cholesterol Accumulation
3. Materials and Methods
3.1. Materials
3.2. Cell Culture
3.3. Cell Viability and Cytotoxicity Tests
3.4. Cholesterol Solubilization by CDs
3.5. In Vitro Hemolysis Tests Using Isolated Human Erythrocytes
3.6. In Vitro Hemolysis Test in Whole Blood Samples
3.7. Generation of Cyclodextrin/Lipid Complexes
3.8. NMR Spectroscopy
3.9. Intracellular Distribution of Cholesterol
3.10. MudPIT Proteomic Analysis
3.11. Data Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davis, M.E.; Brewster, M.E. Cyclodextrin-based pharmaceutics: Past, present and future. Nat. Rev. Drug Discov. 2004, 3, 1023–1035. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Puskas, I.; Szente, L.; Hildreth, J.E. Hyaluronic Acid-Based Biocompatible Supramolecular Assembly for Sustained Release of Antiretroviral Drug. J. Pharm. Sci. 2016, 105, 2760–2769. [Google Scholar] [CrossRef] [PubMed]
- Jahr, J.S.; Miller, J.E.; Hiruma, J.; Emaus, K.; You, M.; Meistelman, C. Sugammadex: A Scientific Review Including Safety and Efficacy, Update on Regulatory Issues, and Clinical Use in Europe. Am. J. Ther. 2015, 22, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Arima, H.; Motoyama, K.; Higashi, T. Potential Use of Cyclodextrins as Drug Carriers and Active Pharmaceutical Ingredients. Chem. Pharm. Bull. 2017, 65, 341–348. [Google Scholar] [CrossRef] [PubMed]
- Alonso, L.; Fontecha, J.; Cuesta, P. Combined effect of Lactobacillus acidophilus and beta-cyclodextrin on serum cholesterol in pigs. Br. J. Nutr. 2016, 115, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.K.; Ahn, J.; Kwak, H.S. Cholesterol removal from homogenized milk with beta-cyclodextrin. J. Dairy Sci. 1999, 82, 2327–2330. [Google Scholar] [CrossRef]
- Alonso, L.; Cuesta, P.; Fontecha, J.; Juarez, M.; Gilliland, S.E. Use of beta-cyclodextrin to decrease the level of cholesterol in milk fat. J. Dairy Sci. 2009, 92, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Maskooki, A.M.; Beheshti, S.H.; Valibeigi, S.; Feizi, J. Effect of Cholesterol Removal Processing Using beta -Cyclodextrin on Main Components of Milk. Int. J. Food Sci. 2013, 2013, 215305. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, Y.; Irie, T.; Uekama, K.; Fukunaga, K.; Pitha, J. Differential effects of alpha-, beta- and gamma-cyclodextrins on human erythrocytes. Eur. J. Biochem. 1989, 186, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Irie, T.; Fukunaga, K.; Pitha, J. Hydroxypropylcyclodextrins in parenteral use. I: Lipid dissolution and effects on lipid transfers in vitro. J. Pharm. Sci. 1992, 81, 521–523. [Google Scholar] [CrossRef] [PubMed]
- Ohvo, H.; Slotte, J.P. Cyclodextrin-mediated removal of sterols from monolayers: Effects of sterol structure and phospholipids on desorption rate. Biochemistry 1996, 35, 8018–8024. [Google Scholar] [CrossRef] [PubMed]
- Irie, T.; Otagiri, M.; Sunada, M.; Uekama, K.; Ohtani, Y.; Yamada, Y.; Sugiyama, Y. Cyclodextrin-induced hemolysis and shape changes of human erythrocytes in vitro. J. Pharmacobio-Dyn. 1982, 5, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Ilangumaran, S.; Hoessli, D.C. Effects of cholesterol depletion by cyclodextrin on the sphingolipid microdomains of the plasma membrane. Biochem. J. 1998, 335 Pt 2, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Kilsdonk, E.P.; Yancey, P.G.; Stoudt, G.W.; Bangerter, F.W.; Johnson, W.J.; Phillips, M.C.; Rothblat, G.H. Cellular cholesterol efflux mediated by cyclodextrins. J. Biol. Chem. 1995, 270, 17250–17256. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J.; Casey, L. Cholesterol levels modulate EGF receptor-mediated signaling by altering receptor function and trafficking. Biochemistry 2002, 41, 10315–10322. [Google Scholar] [CrossRef] [PubMed]
- Rothblat, G.H.; de la Llera-Moya, M.; Atger, V.; Kellner-Weibel, G.; Williams, D.L.; Phillips, M.C. Cell cholesterol efflux: Integration of old and new observations provides new insights. J. Lipid Res. 1999, 40, 781–796. [Google Scholar] [PubMed]
- Szejtli, J. Dimethyl-beta-cyclodextrin as a parenterally applicable drug carrier. J. Incl. Phenom. 1983, 1, 135–138. [Google Scholar] [CrossRef]
- EPAN, 88309597. Cyclodextrin Derivatives with Low Hemolytic Activity; Chinoin Pharmaceutical and Chemical Works: Budapest, Hungary, 1988. [Google Scholar]
- Irie, T.; Uekama, K. Pharmaceutical applications of cyclodextrins. III. Toxicological issues and safety evaluation. J. Pharm. Sci. 1997, 86, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, A.I.; Maxfield, F.R. Niemann-Pick type C disease: Molecular mechanisms and potential therapeutic approaches. J. Neurochem. 2011, 116, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Carstea, E.D.; Morris, J.A.; Coleman, K.G.; Loftus, S.K.; Zhang, D.; Cummings, C.; Gu, J.; Rosenfeld, M.A.; Pavan, W.J.; Krizman, D.B.; et al. Niemann-Pick C1 disease gene: Homology to mediators of cholesterol homeostasis. Science 1997, 277, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Naureckiene, S.; Sleat, D.E.; Lackland, H.; Fensom, A.; Vanier, M.T.; Wattiaux, R.; Jadot, M.; Lobel, P. Identification of HE1 as the second gene of Niemann-Pick C disease. Science 2000, 290, 2298–2301. [Google Scholar] [CrossRef] [PubMed]
- Camargo, F.; Erickson, R.P.; Garver, W.S.; Hossain, G.S.; Carbone, P.N.; Heidenreich, R.A.; Blanchard, J. Cyclodextrins in the treatment of a mouse model of Niemann-Pick C disease. Life Sci. 2001, 70, 131–142. [Google Scholar] [CrossRef]
- Ottinger, E.A.; Kao, M.L.; Carrillo-Carrasco, N.; Yanjanin, N.; Shankar, R.K.; Janssen, M.; Brewster, M.; Scott, I.; Xu, X.; Cradock, J.; et al. Collaborative development of 2-hydroxypropyl-beta-cyclodextrin for the treatment of Niemann-Pick type C1 disease. Curr. Top. Med. Chem. 2014, 14, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, M.; Togawa, M.; Hirabaru, K.; Mochinaga, S.; Narita, A.; Adachi, M.; Egashira, M.; Irie, T.; Ohno, K. Effects of cyclodextrin in two patients with Niemann-Pick Type C disease. Mol. Genet. Metab. 2013, 108, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Soga, M.; Ishitsuka, Y.; Hamasaki, M.; Yoneda, K.; Furuya, H.; Matsuo, M.; Ihn, H.; Fusaki, N.; Nakamura, K.; Nakagata, N.; et al. HPGCD outperforms HPBCD as a potential treatment for Niemann-Pick disease type C during disease modeling with iPS cells. Stem Cells 2015, 33, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Vecsernyes, M.; Fenyvesi, F.; Bacskay, I.; Deli, M.A.; Szente, L.; Fenyvesi, E. Cyclodextrins, Blood-Brain Barrier, and Treatment of Neurological Diseases. Arch. Med. Res. 2014, 45, 711–729. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, E.; Szeman, J.; Csabai, K.; Malanga, M.; Szente, L. Methyl-beta-cyclodextrins: The role of number and types of substituents in solubilizing power. J. Pharm. Sci. 2014, 103, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Nishijo, J.; Moriyama, S.; Shiota, S. Interactions of cholesterol with cyclodextrins in aqueous solution. Chem. Pharm. Bull. 2003, 51, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.A.; de Vries, A.H.; Marrink, S.J. Molecular mechanism of cyclodextrin mediated cholesterol extraction. PLoS Comput. Biol. 2011, 7, e1002020. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Anderson, K.; Baker, J.W. Flocculation of serum lipoproteins with cyclodextrins: Application to assay of hyperlipidemic serum. Clin. Chem. 1990, 36, 529–532. [Google Scholar] [PubMed]
- Kiss, T.; Fenyvesi, F.; Bacskay, I.; Varadi, J.; Fenyvesi, E.; Ivanyi, R.; Szente, L.; Tosaki, A.; Vecsernyes, M. Evaluation of the cytotoxicity of beta-cyclodextrin derivatives: Evidence for the role of cholesterol extraction. Eur. J. Pharm. Sci. 2010, 40, 376–380. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.V.; Tan, A.S. Characterization of the cellular reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT): Subcellular localization, substrate dependence, and involvement of mitochondrial electron transport in MTT reduction. Arch. Biochem. Biophys. 1993, 303, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Creer, M.H.; Ladenson, J. Analytical errors due to lipaemia. Lab. Med. 1983, 14, 351–355. [Google Scholar] [CrossRef]
- Link, A.J.; Eng, J.; Schieltz, D.M.; Carmack, E.; Mize, G.J.; Morris, D.R.; Garvik, B.M.; Yates, J.R., III. Direct analysis of protein complexes using mass spectrometry. Nat. Biotechnol. 1999, 17, 676–682. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DIMEB-50 a | DIMEB-95 b | RAMEB c | HPβCD d | HPγCD e | SBEβCD f | |
|---|---|---|---|---|---|---|
| Cholesterol (mg/mL) | 5.68 ± 0.09 | 4.56 ± 0.04 | 4.97 ± 0.05 | 0.75 ± 0.003 | 0 | 0.08 ± 0.001 |
| Test System | DIMEB-50 a | DIMEB-95 b | RAMEB c | TRIMEB d |
|---|---|---|---|---|
| No plasma | 1.2 ± 0.2 | 1.2 ± 0.3 | 2.3 ± 0.3 | 7.3 ± 0.2 |
| Plasma 50 μL | 8.5 ± 0.2 | 4.7 ± 0.4 | 5.2 ± 0.3 | 11.3 ± 0.3 |
| Plasma 100 μL | 9.4 ± 0.4 | 6.5 ± 0.6 | 7.0 ± 0.4 | 14.7 ± 0.4 |
| Plasma 200 μL | 11.9 ± 0.3 | 8.8 ± 0.5 | 9.7 ± 0.5 | 19.0 ± 0.2 |
| Test System | Plasma Added (μL) | CHO a in Plasma (mM) | HDL b in Plasma (mM) | TG c in Plasma (mM) | Hemolytic EC50 (mg/mL) |
|---|---|---|---|---|---|
| RBCs-washed d | - | - | - | - | 1.32 ± 0.1 |
| RBCs-washed d | 200 | 0.76 | 0.25 | 0.13 | 12.6 ± 0.2 |
| Whole blood e | - | 2.22 | 1.13 | 0.48 | 18.0 ± 0.2 |
| Whole blood e | - | 4.38 | 0.58 | 1.08 | 22.4 ± 0.4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szente, L.; Singhal, A.; Domokos, A.; Song, B. Cyclodextrins: Assessing the Impact of Cavity Size, Occupancy, and Substitutions on Cytotoxicity and Cholesterol Homeostasis. Molecules 2018, 23, 1228. https://doi.org/10.3390/molecules23051228
Szente L, Singhal A, Domokos A, Song B. Cyclodextrins: Assessing the Impact of Cavity Size, Occupancy, and Substitutions on Cytotoxicity and Cholesterol Homeostasis. Molecules. 2018; 23(5):1228. https://doi.org/10.3390/molecules23051228
Chicago/Turabian StyleSzente, Lajos, Ashutosh Singhal, Andras Domokos, and Byeongwoon Song. 2018. "Cyclodextrins: Assessing the Impact of Cavity Size, Occupancy, and Substitutions on Cytotoxicity and Cholesterol Homeostasis" Molecules 23, no. 5: 1228. https://doi.org/10.3390/molecules23051228