Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review





Abstract
:1. Introduction
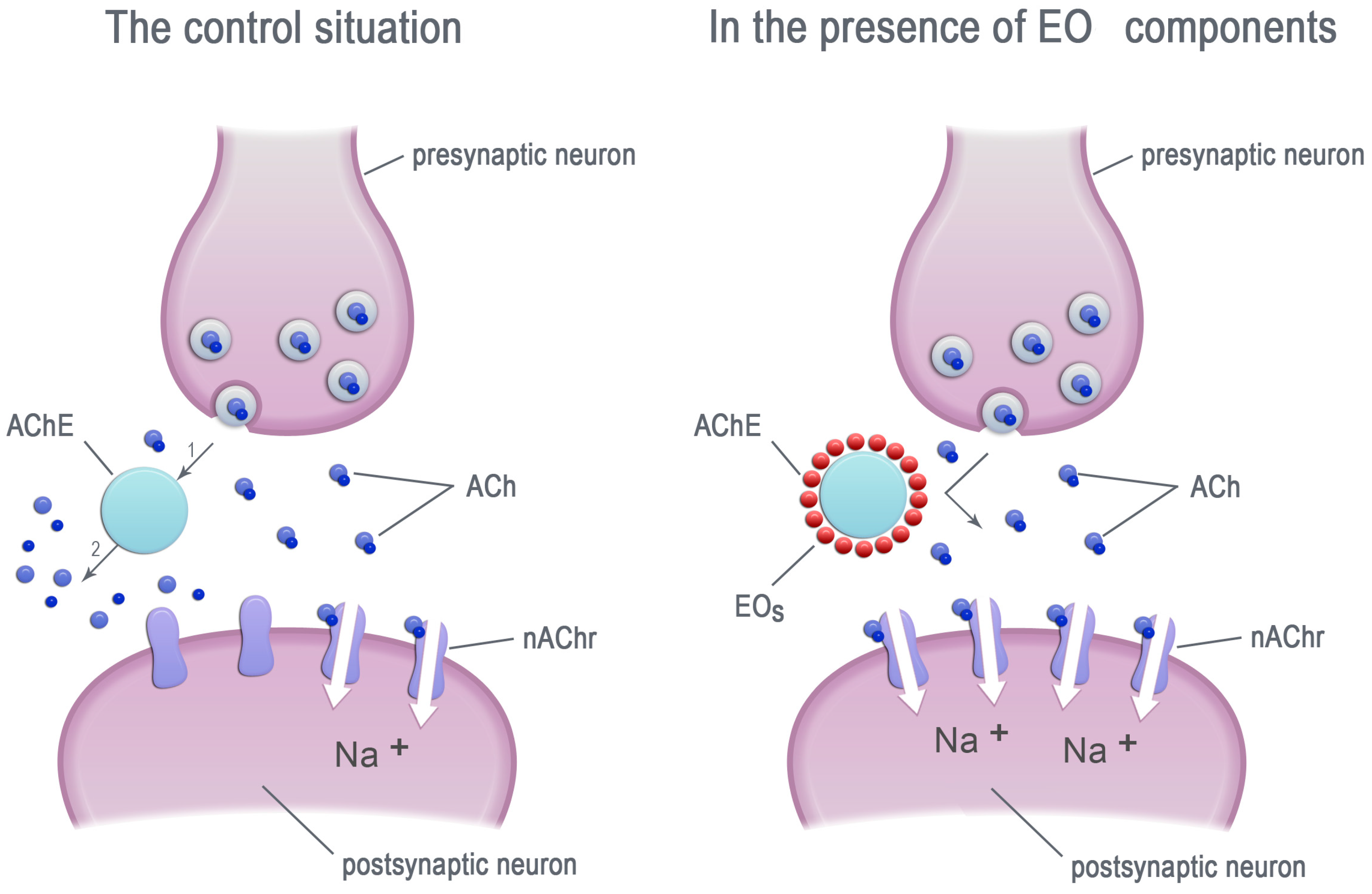
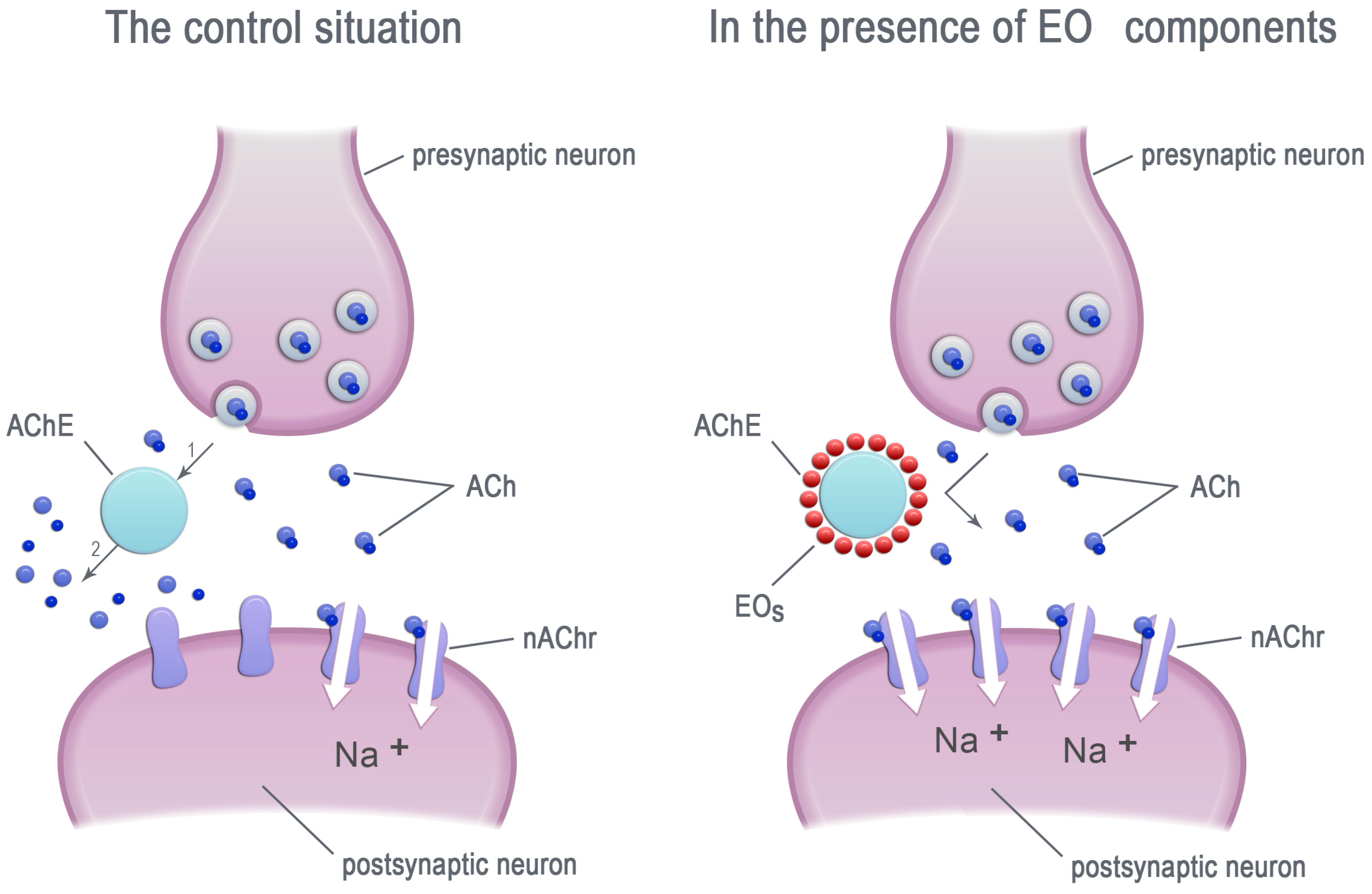
2. Essential Oils—Inhibitors of Acetylcholinesterase
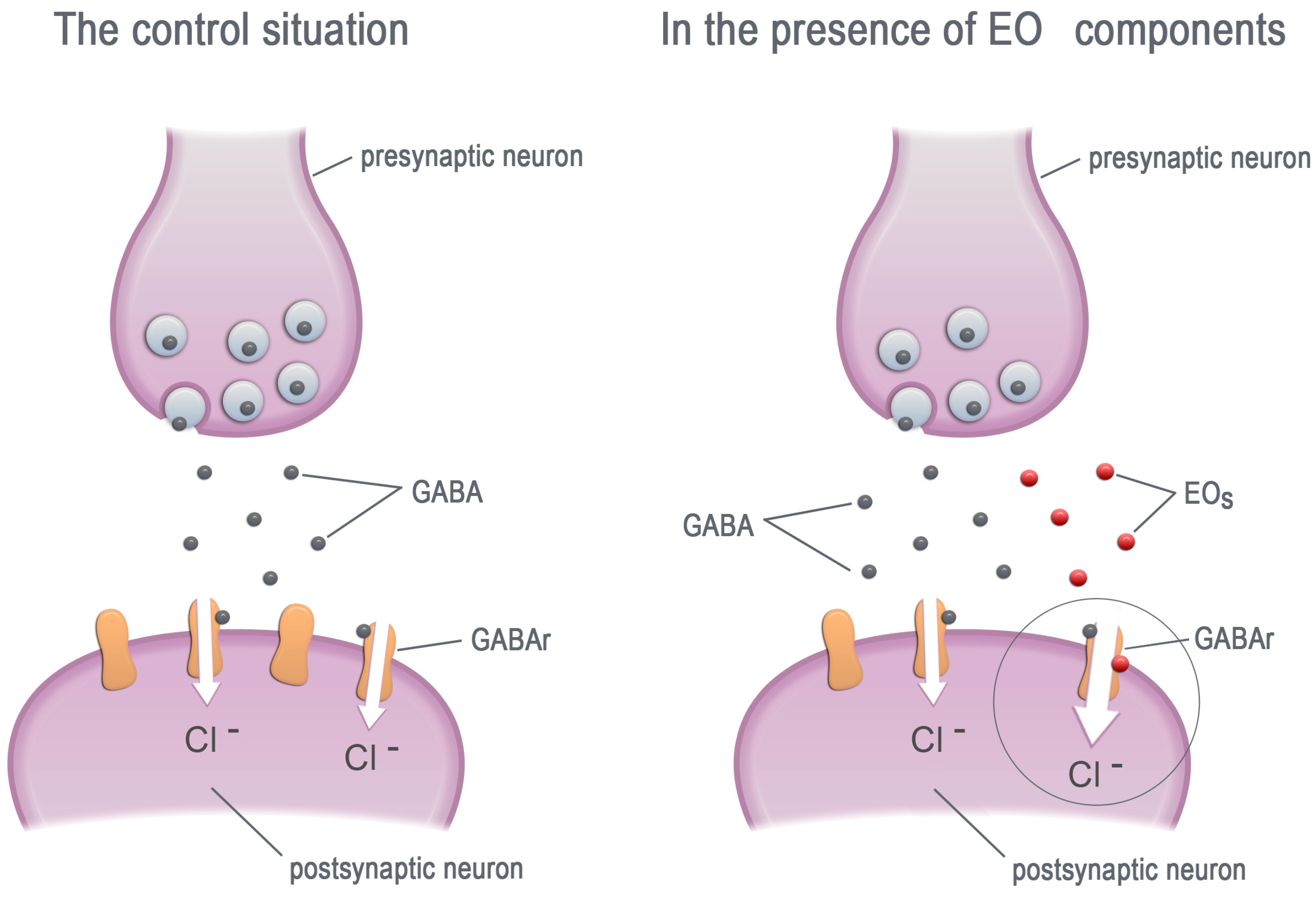
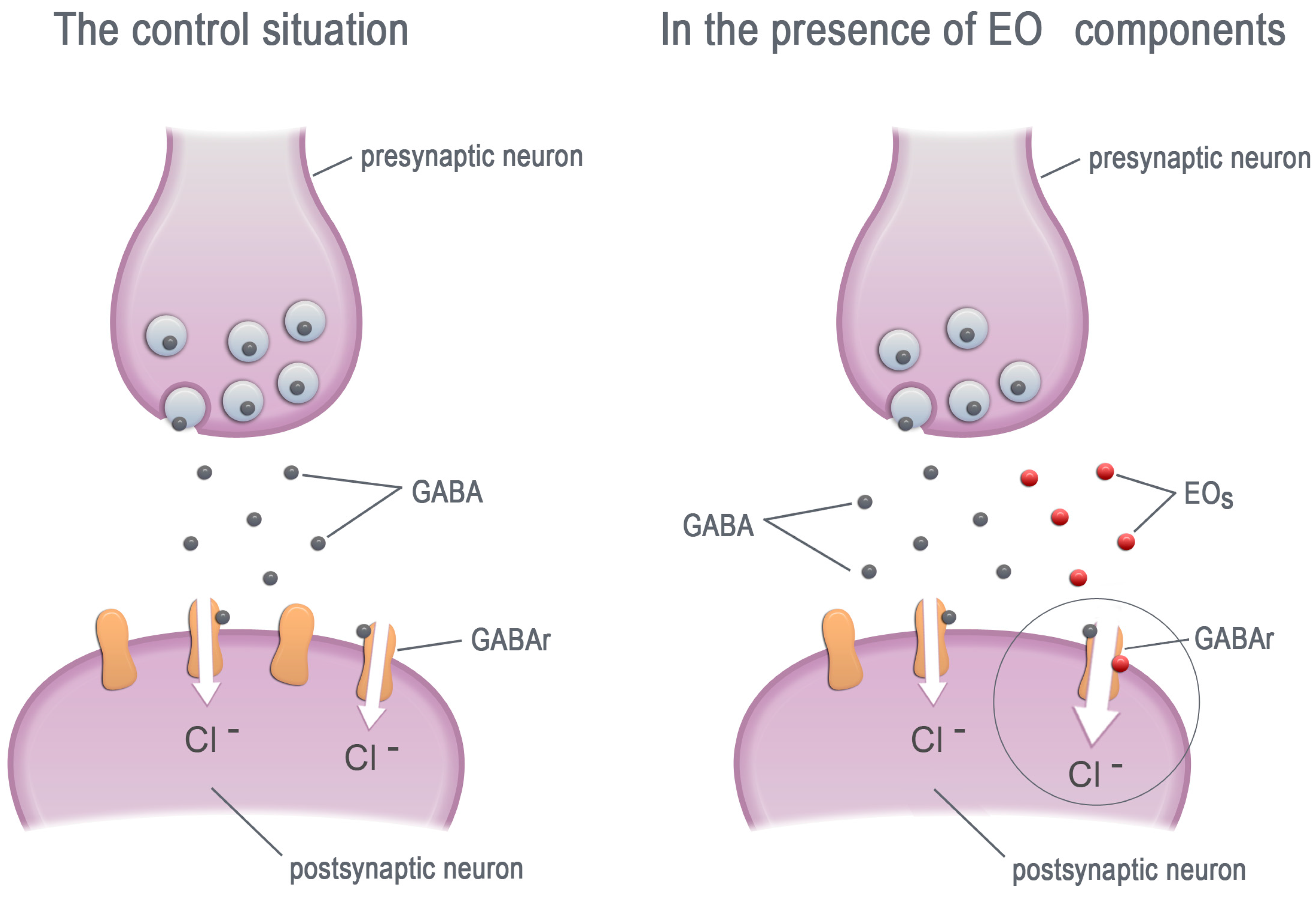
3. Essential Oils—Modifiers of GABA Receptors
3.1. Mammalian GABAA Receptors
3.2. Insect GABA Receptors
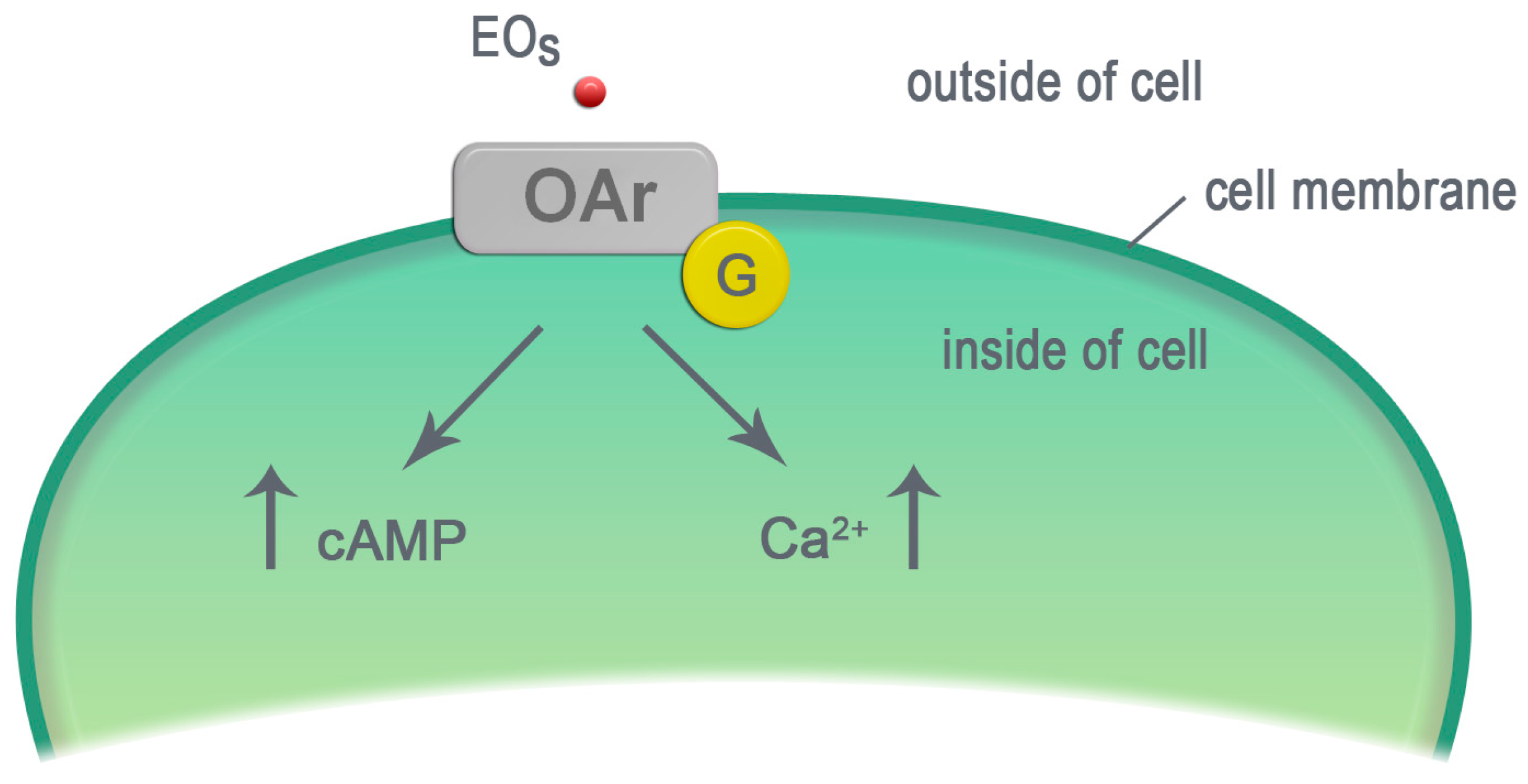
4. Essential Oils—Ligands of Octopamine Receptors
- α-adrenergic-like—the binding of OA to these receptors increases the level of the intracellular calcium; the secondary effect is an increase of the cAMP level;
- β-adrenergic-like—the binding of OA to these receptors increases the level of cAMP;
- octopamine/tyramine—the receptors are similar to α2—an adrenergic receptor in mammals. It is sensitive both to OA and TA. TA binding to this receptor causes a decrease in the cAMP level. In contrast, OA binding to the receptor causes an increase in the cAMP level;
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| No. | Essential Oil Components | AChE Source | IC50 (mM) | Ki (mM) | Reference |
|---|---|---|---|---|---|
| 1 | Anisaldehyde | Electric eel | N.A. | [122] | |
| 2 | Anisole | Human erythrocyte | N.A. | [123] | |
| 3 | Anethole | Electric eel | N.A. | [124] | |
| Electric eel | [67] | ||||
| Electric eel | 0.88 | [125] | |||
| Electric eel | 8.9 | [126] | |||
| Bovine erythrocyte | 0.2 | [127] | |||
| Electric eel | N.A. | [122] | |||
| Electric eel | N.A. | [128] | |||
| Electric eel | 0.87 | [126] | |||
| 4 | Borneol | Bovine erythrocyte | N.A. | [64] | |
| Human erythrocyte | N.A. | [68] | |||
| Bovine erythrocyte | N.A. | [63] | |||
| Electric eel | N.A. | [122] | |||
| Human erythrocyte | N.A. | [123] | |||
| Electric eel | N.A. | [124] | |||
| 5 | Bornyl acetate | Electric eel | 21.3 | [55] | |
| Human erythrocyte | N.A. | [68] | |||
| Bovine erythrocyte | N.A. | [63] | |||
| 6 | Camphene | Electric eel | N.A. | [122] | |
| 7 | Camphor | Electric eel | 0.05 | [67] | |
| Electric eel | N.A | [124] | |||
| Electric eel | 11.2 | [60] | |||
| Human erythrocyte | N.A. | [68] | |||
| Bovine erythrocyte | N.A. | [63] | |||
| Electric eel | N.A. | [122] | |||
| Human erythrocyte | N.A. | [123] | |||
| 8 | 2-Carene | Bovine erythrocyte | 0.9 | [58] | |
| 9 | 3-Carene | Human erythrocyte | 0.2 | [68] | |
| Bovine erythrocyte | 0.2 | [58] | |||
| Electric eel | 0.26 | [126] | |||
| 10 | Carvacrol | Electric eel | 0.41 | [65] | |
| Electric eel | 0.61 | [126] | |||
| Electric eel | 0.21 | [122] | |||
| Electric eel | 0.76 | [128] | |||
| 11 | Carvone | Electric eel | 0.3 | [67] | |
| Bovine erythrocyte | N.A. | [64] | |||
| Electric eel | N.A. | [122] | |||
| Electric eel | 5.5 | [60] | |||
| 12 | Caryophyllene (humulene) | Human erythrocyte | N.A. | [68] | |
| Bovine erythrocyte | 0.13 | [68] | |||
| Electric eel | N.A. | [124] | |||
| Human erythrocyte | N.A. | [68] | |||
| Electric eel | N.A. | [129] | |||
| 13 | Caryophyllene oxide | Human erythrocyte | N.A. | [68] | |
| Bovine erythrocyte | N.A. | [63] | |||
| 14 | 1,8-Cineole | Electric eel | 0.025 | [55] | |
| Electric eel | 0.1 | [124] | |||
| Electric eel | 0.71 | [126] | |||
| Bovine erythrocyte | 0.26 | [64] | |||
| Electric eel | 0.6 | [51] | |||
| Electric eel | 0.84 | [122] | |||
| Human erythrocyte | 0.4 | [68] | |||
| Electric eel | 0.04 | 0.03 | [57] | ||
| Bovine erythrocyte | 0.29 | 0.1 | [58] | ||
| Bovine erythrocyte | 0.39 | [63] | |||
| Human erythrocyte | 0.67 | [130] | |||
| 15 | Cinnamaldehyde | Electric eel | N.A. | [122] | |
| 16 | Cinnamyl alcohol | Electric eel | N.A. | [122] | |
| 17 | Citral | Electric eel | 7 | [55] | |
| Electric eel | N.A. | [124] | |||
| Electric eel | N.A. | [122] | |||
| 18 | Citronellal | Electric eel | N.A. | [122] | |
| 19 | Citronellol | Electric eel | N.A. | [122] | |
| 20 | Copaene | Human erythrocyte | N.A. | [68] | |
| 21 | Cymene | Bovine erythrocyte | N.A. | [58] | |
| 22 | Elemol | Bovine erythrocyte | 0.16 | [64] | |
| 23 | Estragole (Allylanisole) | Electric eel | 0.15 | [67] | |
| Electric eel | 12.6 | [60] | |||
| Electric eel | N.A. | [124] | |||
| Electric eel | N.A. | [122] | |||
| 24 | Eugenol | Electric eel | 2.9 | [124] | |
| Electric eel | N.A. | [122] | |||
| Human erythrocyte | N.A. | [123] | |||
| 25 | Fenchone | Electric eel | 0.4 | [67] | |
| Electric eel | 7 | [60] | |||
| 26 | Geraniol | Electric eel | 0.1 | [67] | |
| Electric eel | 15 | [60] | |||
| Electric eel | N.A. | [122] | |||
| 27 | Globulol | Human erythrocyte | N.A. | [68] | |
| 28 | Gossypol | Electric eel | 1.5 | [55] | |
| 29 | Guaiol | Human erythrocyte | N.A. | [68] | |
| 30 | Isoeugenol | Electric eel | N.A. | [122] | |
| 31 | Limonene | Electric eel | N.A. | [124] | |
| Human erythrocyte | N.A. | [68] | |||
| Electric eel | 1.61 | [125] | |||
| Electric eel | 4.33 | [126] | |||
| Electric eel | N.A. | [122] | |||
| Bovine erythrocyte | N.A. | [64] | |||
| 32 | Linalool | Electric eel | 0.3 | [67] | |
| Electric eel | 5.5 | [55] | |||
| Electric eel | N.A. | [124] | |||
| Electric eel | 15.6 | [60] | |||
| Electric eel | N.A. | [122] | |||
| Human erythrocyte | N.A. | [68] | |||
| Bovine erythrocyte | N.A. | [63] | |||
| Bovine erythrocyte | N.A. | [64] | |||
| 33 | Linalyl acetate | Bovine erythrocyte | N.A. | [64] | |
| Electric eel | N.A. | [129] | |||
| 34 | Manool | Human erythrocyte | N.A. | [68] | |
| 35 | Methylcinnamate | Electric eel | N.A. | [122] | |
| 36 | Methyleugenol | Electric eel | N.A. | [122] | |
| Electric eel | N.A. | [124] | |||
| 37 | Menthofuran | Bovine erythrocyte | N.A. | [64] | |
| 38 | Menthol | Bovine erythrocyte | N.A. | [64] | |
| 39 | Menthone | Bovine erythrocyte | N.A. | [64] | |
| Electric eel | N.A. | [122] | |||
| 40 | Methol | Human erythrocyte | N.A. | [123] | |
| 41 | Methoxycinnamaldehyde | Electric eel | N.A. | [124] | |
| 42 | Methyl acetate | Bovine erythrocyte | N.A. | [64] | |
| 43 | Myrcene | Electric eel | N.A. | [122] | |
| 44 | Myrtenal | Electric eel | 0.17 | [122] | |
| 45 | Nerol | Electric eel | N.A. | [122] | |
| 46 | Nerolidol | Electric eel | N.A. | [122] | |
| 47 | Neryl acetate | Human erythrocyte | N.A. | [68] | |
| 48 | Phellandrene | Electric eel | 0.88 | [129] | |
| 49 | Phenylethanol | Electric eel | N.A. | [122] | |
| 50 | α-Pinene | Electric eel | 0.16 | [124] | |
| Electric eel | 10.5 | [130] | |||
| Human erythrocyte | 0.7 | [68] | |||
| Human erythrocyte | 0.63 | [131] | |||
| Bovine erythrocytes | 0.66 | [63] | |||
| Electric eel | N.A. | [122] | |||
| Bovine erythrocytes | 0.4 | [58] | |||
| 51 | β-Pinene | Electric eel | N.A. | [124] | |
| Human erythrocyte | 1.5 | [68] | |||
| Bovine erythrocyte | 1.5 | [63] | |||
| Electric eel | N.A. | [122] | |||
| 52 | Piperitenone oxide | Bovine erythrocyte | 0.38 | [64] | |
| 53 | Piperitenone | Bovine erythrocyte | 0.72 | [64] | |
| Bovine erythrocyte | 0.83 | [64] | |||
| 54 | Pulegone | Electric eel | 0.85 | [55] | |
| Bovine erythrocyte | 0.89 | [64] | |||
| 55 | Sabinene | Human erythrocyte | N.A. | [68] | |
| Electric eel | 1.25 | [125] | |||
| 56 | Sclareol | Human erythrocyte | N.A. | [68] | |
| 57 | α-Terpinene | Bovine erythrocyte | N.A. | [58] | |
| 58 | γ-Terpinene | Electric eel | 0.2 | [67] | |
| Electric eel | N.A. | [124] | |||
| Electric eel | 5.8 | [60] | |||
| Bovine erythrocyte | N.A. | [58] | |||
| 59 | α-Terpineol | Electric eel | 8.43 | [124] | |
| Human erythrocyte | N.A. | [68] | |||
| 60 | Terpinen-4-ol | Electric eel | 20.7 | [124] | |
| Electric eel | 10.30 | 4.7 | [57] | ||
| Bovine erythrocyte | N.A. | 2 | [58] | ||
| Electric eel | N.A. | [129] | |||
| Electric eel | N.A. | [122] | |||
| 61 | Terpinolene | Electric eel | 1.1 | [129] | |
| 62 | α-Thujone | Human erythrocyte | N.A. | [68] | |
| Electric eel | N.A. | [122] | |||
| 63 | Thymohydroquinone | Electric eel | 0.24 | [65] | |
| 64 | Thymol | Electric eel | 4.9 | [65] | |
| Electric eel | 1.39 | [126] | |||
| Electric eel | N.A. | [122] | |||
| 65 | Thymoquinone | Electric eel | 0.85 | [65] | |
| 66 | Viridiflorol | Bovine erythrocyte | 0.11 | [64] | |
| 67 | Verbenone | Electric eel | 2.66 | [122] | |
| Electric eel | 0.73 | [128] |
References
- Bassolé, I.H.N.; Juliani, H.R. Essential oils in combination and their antimicrobial properties. Molecules 2012, 17, 3989–4006. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against insects—A review. Plant Protect. Sci. 2016, 52, 229–241. [Google Scholar]
- Pavela, R.; Benelli, G. Ethnobotanical knowledge on botanical repellents employed in the African region against mosquito vectors—A review. Exp. Parasitol. 2016, 167, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Mihajilov-Krstev, T.; Jovanović, B.; Jović, J.; Ilić, B.; Miladinović, D.; Matejić, J.; Rajković, J.; Dorđević, L.; Cvetković, V.; Zlatković, B. Antimicrobial, antioxidative, and insect repellent effects of Artemisia absinthium essential oil. Planta Medica 2014, 80, 1698–1705. [Google Scholar] [CrossRef] [PubMed]
- Tisgratog, R.; Sanguanpong, U.; Grieco, J.P.; Ngoen-Kluan, R.; Chareonviriyaphap, T. Plants traditionally used as mosquito repellents and the implication for their use in vector control. Acta Trop. 2016, 157, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Price, D.N.; Berry, M.S. Neurophysiological effects of naturally occurring defensive compounds on the freshwater snail Planorbis corneus: Comparison with effects in insects. J. Chem. Ecol. 2008, 34, 994–1004. [Google Scholar] [CrossRef] [PubMed]
- Cilek, J.E.; Hallmon, C.F.; Johnson, R. Limited efficacy of commercially formulated essential oils on vegetation against female Aedes albopictus and Culex quinquefasciatus. J. Agric. Urban Entomol. 2010, 27, 54–60. [Google Scholar] [CrossRef]
- Liu, Z.L.; Zhao, N.N.; Liu, C.M.; Zhou, L.; Du, S.S. Identification of insecticidal constituents of the essential oil of Curcuma wenyujin rhizomes active against Liposcelis bostrychophila Badonnel. Molecules 2012, 17, 12049–12060. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.C.; Li, Y.P.; Li, H.Q.; Deng, Z.W.; Zhou, L.; Liu, Z.L.; Du, S.S. Identification of repellent and insecticidal constituents of the essential oil of Artemisia rupestris L. aerial parts against Liposcelis bostrychophila Badonnel. Molecules 2013, 18, 10733–10746. [Google Scholar] [PubMed]
- Liu, X.C.; Zhou, L.G.; Liu, Z.L.; Du, S.S. Identification of insecticidal constituents of the essential oil of Acorus calamus rhizomes against Liposcelis bostrychophila Badonnel. Molecules 2013, 18, 5684–5696. [Google Scholar] [CrossRef] [PubMed]
- Du, S.S.; Yang, K.; Wang, C.F. Chemical constituents and activities of the essential oil from Myristica fragrans against cigarette beetle Lasioderma serricorne. Chem. Biodivers. 2014, 11, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.C.; Liang, Y.; Shi, W.P.; Liu, Q.Z.; Zhou, L.; Liu, Z.L. Repellent and insecticidal effects of the essential oil of Kaempferia galanga rhizomes to Liposcelis bostrychophila (Psocoptera: Liposcelidae). J. Econ. Entomol. 2014, 107, 1706–1712. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; You, C.X.; Wang, C.F.; Yang, K.; Chen, R.; Zhang, W.J.; Du, S.S.; Geng, Z.F.; Deng, Z.W. Chemical constituents and insecticidal activities of the essential oil from Amomum tsaoko against two stored-product insects. J. Oleo Sci. 2014, 63, 1019–1026. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Chen, Z.; Yin, Z. In vitro acaricidal activity of 1,8-cineole against Sarcoptes scabiei var cuniculi and regulating effects on enzyme activity. Parasitol. Res. 2015, 114, 2959–2967. [Google Scholar] [PubMed]
- Liu, X.C.; Liu, Z.L. Analysis of the essential oil of Illicium henryi Diels root bark and its insecticidal activity against Liposcelis bostrychophila Badonnel. J. Food Prot. 2015, 78, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; You, C.X.; Yang, K.; Wu, Y.; Chen, R.; Zhang, W.J.; Liu, Z.L.; Du, S.S.; Deng, Z.W.; Geng, Z.F.; et al. Bioactivity of essential oil of Zingiber purpureum Rhizomes and its main compounds against two stored product insects. J. Econ. Entomol. 2015, 108, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, W.J.; Wang, P.J.; Yang, K.; Huang, D.Y.; Wei, J.Y.; Tian, Z.F.; Bai, J.F.; Du, S.S. Contact toxicity and repellency of the essential oil of Liriope muscari (DECN.) Bailey against three insect tobacco storage pests. Molecules 2015, 20, 1676–1685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.J.; Yang, K.; You, C.X.; Wang, Y.; Wang, C.F.; Wu, Y.; Geng, Z.F.; Su, Y.; Du, S.S.; Deng, Z.W. Bioactivity of essential oil from Artemisia stolonifera (Maxim.) Komar. and its main compounds against two stored-product insects. J. Oleo Sci. 2015, 64, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.A.; Roman-Campos, D.; Carvalho, M.S.; Miranda, F.M.; Carneiro, D.C.; Cavalcante, P.H.; Cândido, E.A.; Filho, L.X.; Cruz, J.S.; Gondim, A.N. Cardiodepressive effect elicited by the essential oil of Alpinia speciosa is related to L-type Ca2+ current blockade. Phytomedicine 2011, 18, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, E.L.D.; Lima, J.K.A.; Souza, F.H.O.; Silva, I.M.; Santos, A.A.; Araújo, A.P.; Blank, A.F.; Lima, R.N.; Alves, P.B.; Bacci, L. Insecticidal and repellence activity of the essential oil of Pogostemon cablin against urban ants species. Acta Trop. 2013, 127, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Dutra, K.A.; de Oliveira, J.V.; Navarro, D.M.; Barbosa, D.A.; Santos, J.P. Control of Callosobruchus maculatus (FABR.) (Coleoptera: Chrysomelidae: Bruchinae) in Vigna unguiculata (L.) WALP. with essential oils from four Citrus spp. plants. J. Stored Prod. Res. 2016, 68, 25–32. [Google Scholar] [CrossRef]
- Nathan, S.S.; Hisham, A.; Jayakumar, G. Larvicidal and growth inhibition of the malaria vector Anopheles stephensi by triterpenes from Dysoxylum malabaricum and Dysoxylum beddomei. Fitoterapia 2008, 79, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Cosimi, S.; Rossi, E.; Cioni, P.L.; Canale, A. Bioactivity and qualitative analysis of some essential oils from Mediterranean plants against stored-product pests: Evaluation of repellency against Sitophilus zeamais Motschulsky, Cryptolestes ferrugineus (Stephens) and Tenebrio molitor (L.). J. Stored Prod. Res. 2009, 45, 125–132. [Google Scholar] [CrossRef]
- Reegan, A.D.; Gandhi, M.R.; Paulraj, M.G.; Balakrishna, K.; Ignacimuthu, S. Effect of niloticin, a protolimonoid isolated from Limonia acidissima L. (Rutaceae) on the immature stages of dengue vector Aedes aegypti L. (Diptera: Culicidae). Acta Trop. 2014, 139, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Benelli, G. α-Humulene and β-elemene from Syzygium zeylanicum (Myrtaceae) essential oil: Highly effective and eco-friendly larvicides against Anopheles subpictus, Aedes albopictus, and Culex tritaeniorhynchus (Diptera: Culicidae). Parasitol. Res. 2016, 115, 2771–2778. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.J.; Juárez, M.L.; Alzogaray, R.A.; Arrighi, F.; Arroyo, L.; Gastaminza, G.; Willink, E.; Bardón, A.V.; Vera, T. Toxic effect of citrus peel constituents on Anastrepha fraterculus Wiedemann and Ceratitis capitata Wiedemann immature stages. J. Agric. Food Chem. 2014, 62, 10084–10091. [Google Scholar] [CrossRef] [PubMed]
- Govindarajan, M.; Sivakumar, R.; Rajeswary, M.; Veerakumar, K. Mosquito larvicidal activity of thymol from essential oil of Coleus aromaticus Benth. against Culex tritaeniorhynchus, Aedes albopictus, and Anopheles subpictus (Diptera: Culicidae). Parasitol. Res. 2013, 112, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Price, D.N.; Berry, M.S. Comparison of effects of octopamine and insecticidal essential oils on activity in the nerve cord, foregut, and dorsal unpaired median neurons of cockroaches. J. Insect Physiol. 2006, 52, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitol. Res. 2015, 114, 3835–3853. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R. Acute, synergistic and antagonistic effects of some aromatic compounds on the Spodoptera littoralis Boisd. (Lep., Noctuidae) larvae. Ind. Crop. Prod. 2014, 60, 247–258. [Google Scholar] [CrossRef]
- Novato, T.P.; Araújo, L.X.; de Monteiro, C.M.; Maturano, R.; Senra Tde, O.; da Silva Matos, R.; Gomes, G.A.; de Carvalho, M.G.; Daemon, E. Evaluation of the combined effect of thymol, carvacrol and (E)-cinnamaldehyde on Amblyomma sculptum (Acari: Ixodidae) and Dermacentor nitens (Acari: Ixodidae) larvae. Vet. Parasitol. 2015, 212, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Ingkaninan, K.; Temkitthawon, P.; Chuenchom, K.; Yuyaem, T.; Thongnoi, W. Screening for acetylcholinesterase inhibitory activity in plants used in Thai traditional rejuvenating and neurotonic remedies. J. Ethnopharmacol. 2003, 89, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Gnagey, A.L.; Forte, M.; Rosenberry, T.L. Isolation and characterization of acetylcholinesterase from Drosophila. J. Biol. Chem. 1987, 262, 13290–13298. [Google Scholar] [PubMed]
- Bourguet, D.; Roig, A.; Toutant, J.P.; Arpagaus, M. Analysis of molecular forms and pharmacological properties of acetylcholinesterase in several mosquito species. Neurochem. Int. 1997, 31, 65–72. [Google Scholar] [CrossRef]
- Marcel, V.; Palacios, L.G.; Pertuy, C.; Masson, P.; Fournier, D. Two invertebrate acetylcholinesterases show activation followed by inhibition with substrate concentration. Biochem. J. 1998, 329, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Jung, C.S.; Koh, Y.H.; Lee, S.H. Molecular, biochemical and histochemical characterization of two acetylcholinesterase cDNAs from the German cockroach Blattella germanica. Insect Biochem. Mol. Biol. 2006, 15, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Pezzementi, L.; Rowland, M.; Wolfe, M.; Tsigelny, I. Inactivation of an invertebrate acetylcholinesterase by sulfhydryl reagents: The roles of two cysteines in the catalytic gorge of the enzyme. Invertebr. Neurosci. 2006, 6, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.P.; Singh, S.K.; Gao, Y.; Lassiter, L.T.; Mishra, R.K.; Zhu, K.Y.; Brimijoin, S. Selective and irreversible inhibitors of aphid acetylcholinesterases: Steps toward human-safe insecticides. PLoS ONE 2009, 4, e4349. [Google Scholar] [CrossRef] [PubMed]
- Polsinelli, G.A.; Singh, S.K.; Mishra, R.K.; Suranyi, R.; Ragsdale, D.W.; Pang, Y.P.; Brimijoin, S. Insect-specific irreversible inhibitors of acetylcholinesterase in pests including the bed bug, the eastern yellowjacket, German and American cockroaches, and the confused flour beetle. Chem. Biol. Interact. 2010, 187, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.P.; Brimijoin, S.; Ragsdale, D.W.; Zhu, K.Y.; Suranyi, R. Novel and viable acetylcholinesterase target site for developing effective and environmentally safe insecticides. Curr. Drug Targets 2012, 13, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.M.; Kim, J.; Kang, J.S.; Koh, S.H.; Ahn, Y.J.; Kang, K.S.; Park, I.K. Fumigant toxicity and acetylcholinesterase inhibitory activity of 4 Asteraceae plant essential oils and their constituents against Japanese termite (Reticulitermes speratus Kolbe). Pestic. Biochem. Physiol. 2014, 113, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.E.; Şenol, F.S.; Gülpinar, A.R.; Kartal, M.; Sekeroglu, N.; Deveci, M.; Kan, Y.; Sener, B. Acetylcholinesterase inhibitory and antioxidant properties of Cyclotrichium niveum, Thymus praecox subsp. caucasicus var. caucasicus, Echinacea purpurea and E. pallida. Food Chem. Toxicol. 2009, 47, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Tel, G.; Öztürk, M.; Duru, M.E.; Harmandar, M.; Topçu, G. Chemical composition of the essential oil and hexane extract of Salvia chionantha and their antioxidant and anticholinesterase activities. Food Chem. Toxicol. 2010, 48, 3189–3193. [Google Scholar] [CrossRef] [PubMed]
- Orhan, I.E.; Senol, F.S.; Ozturk, N.; Celik, S.A.; Pulur, A.; Kan, Y. Phytochemical contents and enzyme inhibitory and antioxidant properties of Anethum graveolens L. (dill) samples cultivated under organic and conventional agricultural conditions. Food Chem. Toxicol. 2013, 59, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Perry, N.S.L.; Houghton, P.J.; Jenner, P.; Keith, A.; Perry, E.K. Salvia lavandulaefolia essential oil inhibits cholinesterase in vivo. Phytomedicine 2002, 9, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Kim, E.; Lee, S.H.; Park, I.K. Inhibition of acetylcholinesterases of the pinewood nematode, Bursaphelenchus xylophilus, by phytochemicals from plant essential oils. Pestic. Biochem. Physiol. 2013, 105, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Yeom, H.J.; Jung, C.S.; Kang, J.S.; Kim, J.; Lee, J.H.; Kim, D.S.; Kim, H.S.; Park, P.S.; Kang, K.S.; Park, I.K. Insecticidal and acetylcholine esterase inhibition activity of Asteraceae plant essential oils and their constituents against adults of the German cockroach (Blattella germanica). J. Agric. Food Chem. 2015, 63, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [Google Scholar] [CrossRef]
- Park, C.G.; Jang, M.; Yoon, K.A.; Kim, J. Insecticidal and acetylcholinesterase inhibitory activities of Lamiaceae plant essential oils and their major components against Drosophila suzukii (Diptera: Drosophilidae). Ind. Crop. Prod. 2016, 89, 507–513. [Google Scholar] [CrossRef]
- Lee, S.E.; Lee, B.H.; Choi, W.S.; Park, B.S.; Kim, J.G.; Campbell, B.C. Fumigant toxicity of volatile natural products from Korean spices and medicinal plants towards the rice weevil, Sitophilus oryzae (L). Pest Manag. Sci. 2001, 57, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Picollo, M.I.; Toloza, A.C.; Mougabure Cueto, G.; Zygadlo, J.; Zerba, E. Anticholinesterase and pediculicidal activities of monoterpenoids. Fitoterapia 2008, 79, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Park, I.K. Fumigant toxicity of oriental sweetgum (Liquidambar orientalis) and valerian (Valeriana wallichii) essential oils and their components, including their acetylcholinesterase inhibitory activity, against Japanese Termites (Reticulitermes speratus). Molecules 2014, 19, 12547–12558. [Google Scholar] [CrossRef] [PubMed]
- Reegan, A.D.; Stalin, A.; Paulraj, M.G.; Balakrishna, K.; Ignacimuthu, S.; Al-Dhabi, N.A. In silico molecular docking of niloticin with acetylcholinesterase 1 (AChE1) of Aedes aegypti L. (Diptera: Culicidae): A promising molecular target. Med. Chem. Res. 2016, 25, 1411–1419. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.M.; Mohamed, M.I.E.; Badawy, M.E.I.; El-Arami, S.A.A. Fumigant and contact toxicities of monoterpenes to Sitophilus oryzae (L.) and Tribolium castaneum (herbst) and their inhibitory effects on acetylcholinesterase activity. J. Chem. Ecol. 2009, 35, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.F.; Byrne, O. Plant-insect coevolution and inhibition of acetylcholinesterase. J. Chem. Ecol. 1988, 14, 1965–1975. [Google Scholar] [CrossRef] [PubMed]
- Park, T.J.; Seo, H.K.; Kang, B.J.; Kim, K.T. Noncompetitive inhibition by camphor of nicotinic acetylcholine receptors. Biochem. Pharmacol. 2001, 61, 787–793. [Google Scholar] [CrossRef]
- Mills, C.; Cleary, B.J.; Gilmer, J.F.; Walsh, J.J. Inhibition of acetylcholinesterase by tea tree oil. J. Pharm. Pharmacol. 2004, 56, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Yamafuji, C. Inhibition of acetylcholinesterase activity by bicyclic monoterpenoids. J. Agric. Food Chem. 2005, 53, 1765–1768. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Yamafuji, C. Inhibition of acetylcholinesterase activity by tea tree oil and constituent terpenoids. Flavour Fragr. J. 2006, 21, 198–201. [Google Scholar] [CrossRef]
- López, M.D.; Campoy, F.J.; Pascual-Villalobos, M.J.; Muñoz-Delgado, E.; Vidal, C.J. Acetylcholinesterase activity of electric eel is increased or decreased by selected monoterpenoids and phenylpropanoids in a concentration-dependent manner. Chem. Biol. Interact. 2015, 229, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Radić, Z. The cholinesterases: From genes to proteins. Annu. Rev. Pharmacol. Toxicol. 1994, 34, 281–320. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Ruiz, P.; Rubio, L.; García-Palomero, E.; Dorronsoro, I.; del Monte-Millán, M.; Valenzuela, R.; Usán, P.; de Austria, C.; Bartolini, M.; Andrisano, V.; et al. Design, synthesis, and biological evaluation of dual binding site acetylcholinesterase inhibitors: New disease-modifying agents for Alzheimer’s disease. J. Med. Chem. 2005, 48, 7223–7233. [Google Scholar] [CrossRef] [PubMed]
- Savelev, S.U.; Okello, E.; Perry, N.S.L.; Wilkins, R.M.; Perry, E.K. Synergistic and antagonistic interactions of anticholinesterase terpenoids in Salvia lavandulaefolia essential oil. Pharmacol. Biochem. Behav. 2003, 75, 661–668. [Google Scholar] [CrossRef]
- Miyazawa, M.; Watanabe, H.; Umemoto, K.; Kameoka, H. Inhibition of acetylcholinesterase activity by essential oils of mentha species. J. Agric. Food Chem. 1998, 46, 3431–3434. [Google Scholar] [CrossRef]
- Jukic, M.; Politeo, O.; Maksimovic, M.; Milos, M.; Milos, M. In vitro acetylcholinesterase inhibitory properties of thymol, carvacrol and their derivatives thymoquinone and thymohydroquinone. Phytother. Res. 2007, 21, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Dambolena, J.S.; Zunino, M.P.; Herrera, J.M.; Pizzolitto, R.P.; Areco, V.A.; Zygadlo, J.A. Terpenes: Natural products for controlling insects of importance to human health—A structure-activity relationship study. Psyche 2016, 2016, 4595823. [Google Scholar] [CrossRef]
- López, M.D.; Pascual-Villalobos, M.J. Mode of inhibition of acetylcholinesterase by monoterpenoids and implications for pest control. Ind. Crop. Prod. 2010, 31, 284–288. [Google Scholar] [CrossRef]
- Savelev, S.U.; Okello, E.J.; Perry, E.K. Butyryl- and acetyl-cholinesterase inhibitory activities in essential oils of Salvia species and their constituents. Phytother. Res. 2004, 18, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Sattelle, D.B. GABA receptors of insects. Adv. Insect Physiol. 1990, 22, 1–113. [Google Scholar]
- Ben-Ari, Y.; Khalilov, I.; Kahle, K.T.; Cherubini, E. The GABA excitatory/inhibitory shift in brain maturation and neurological disorders. Neuroscientist 2012, 18, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Kandel, E.R. Principles of Neural Science, 5th ed.; Schwartz, J.H., Jessell, T.M., Siegelbaum, S., Hudspeth, A.J., Eds.; McGraw-Hill: New York, NY, USA, 2013. [Google Scholar]
- Sigel, E.; Steinmann, M.E. Structure, function, and modulation of GABAA receptors. J. Biol. Chem. 2012, 287, 40224–40231. [Google Scholar] [CrossRef] [PubMed]
- Bloomquist, J.R. Chloride channels as tools for developing selective insecticides. Arch. Insect Biochem. Physiol. 2003, 54, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W.; Sieghart, W. GABA A receptors: Subtypes provide diversity of function and pharmacology. Neuropharmacology 2009, 56, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Priestley, C.M.; Williamson, E.M.; Wafford, K.A.; Sattelle, D.B. Thymol, a constituent of thyme essential oil, is a positive allosteric modulator of human GABAA receptors and a homo-oligomeric GABA receptor from Drosophila melanogaster. Br. J. Pharmacol. 2003, 140, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.C.; Turcotte, C.M.; Betts, B.A.; Yeung, W.Y.; Agyeman, A.S.; Burk, L.A. Modulation of human GABAA and glycine receptor currents by menthol and related monoterpenoids. Eur. J. Pharmacol. 2004, 506, 9–16. [Google Scholar] [CrossRef] [PubMed]
- García, D.A.; Bujons, J.; Vale, C.; Suñol, C. Allosteric positive interaction of thymol with the GABAA receptor in primary cultures of mouse cortical neurons. Neuropharmacology 2006, 50, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Watt, E.E.; Betts, B.A.; Kotey, F.O.; Humbert, D.J.; Griffith, T.N.; Kelly, E.W.; Veneskey, K.C.; Gill, N.; Rowan, K.C.; Jenkins, A.; et al. Menthol shares general anesthetic activity and sites of action on the GABA(A) receptor with the intravenous agent, propofol. Eur. J. Pharmacol. 2008, 590, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.J.; Aoshima, H.; Koda, H.; Kiso, Y. Fragrances in oolong tea that enhance the response of GABAA receptors. Biosci. Biotechnol. Biochem. 2004, 68, 1842–1848. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Huang, C.; Peng, Z.; Xie, Y.; Deng, S.; Nie, Y.Z.; Xu, T.L.; Ge, W.H.; Li, W.G.; Li, F. Electrophysiological characterization of methyleugenol: A novel agonist of GABA(A) receptors. ACS Chem. Neurosci. 2014, 5, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Höld, K.M.; Sirisoma, N.S.; Ikeda, T.; Narahashi, T.; Casida, J.E. α-Thujone (the active component of absinthe): γ-aminobutyric acid type A receptor modulation and metabolic detoxification. Proc. Natl. Acad. Sci. USA 2000, 97, 3826–3831. [Google Scholar]
- Aoshima, H.; Hamamoto, K. Potentiation of GABAA receptors expressed in Xenopus oocytes by perfume and phytoncid. Biosci. Biotechnol. Biochem. 1999, 63, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36Cl− uptake in American cockroach ventral nerve cord. Pestic. Biochem. Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Granger, R.E.; Campbell, E.L.; Johnston, G.A.R. (+)- And (−)-borneol: Efficacious positive modulators of GABA action at human recombinant α1β2γ2L GABAA receptors. Biochem. Pharmacol. 2005, 69, 1101–1111. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.J.; Hamamoto, K.; Aoshima, H.; Hara, Y. Effects of tea components on the response of GABA A receptors expressed in Xenopus oocytes. J. Agric. Food Chem. 2002, 50, 3954–3960. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Borzone, M.; Delgado-Marín, L.; García, D.A. Inhibitory effects of carvone isomers on the GABAA receptor in primary cultures of rat cortical neurons. Chirality 2014, 26, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Yip, G.M.S.; Chen, Z.W.; Edge, C.J.; Smith, E.H.; Dickinson, R.; Hohenester, E.; Townsend, R.R.; Fuchs, K.; Sieghart, W.; Evers, A.S.; et al. A propofol binding site on mammalian GABAA receptors identified by photolabeling. Nat. Chem. Biol. 2013, 9, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Jayakar, S.S.; Zhou, X.; Chiara, D.C. Multiple propofol-binding sites in a γ-aminobutyric acid type A receptor (GABAAR) identified using a photoreactive propofol analog. J. Biol. Chem. 2014, 289, 27456–27468. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.W. Allosteric ligands and their binding sites define γ-aminobutyric acid (GABA) type A receptor subtypes. Adv. Pharmacol. 2015, 73, 167–202. [Google Scholar] [PubMed]
- Ascari, J.; Sens, S.L.; Nunes, D.S.; Wisniewski, A., Jr.; Arbo, M.D.; Linck, V.M.; Lunardi, P.; Leal, M.B.; Elisabetsky, E. Sedative effects of essential oils obtained from Baccharis uncinella. Pharm. Biol. 2012, 50, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.L.; Garlet, Q.I.; Benovit, S.C.; Dolci, G.; Mallmann, C.A.; Bürger, M.E.; Baldisserotto, B.; Longhi, S.J.; Heinzmann, B.M. Sedative and anesthetic activities of the essential oils of Hyptis mutabilis (Rich.) Briq. and their isolated components in silver catfish (Rhamdia quelen). Braz. J. Med. Biol. Res. 2013, 46, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Henderson, J.E.; Soderlund, D.M.; Knipple, D.C. Characterization of a putative γ-aminobutyric-acid (GABA) receptor β-subunit gene from Drosophila melanogaster. Biochem. Biophys. Res. Commun. 1993, 193, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Aronstein, K.; Auld, V.; Ffrench-Constant, R. Distribution of two GABA receptor-like subunits in the Drosophila CNS. Invertebr. Neurosci. 1996, 2, 115–120. [Google Scholar] [CrossRef]
- Dupuis, J.P.; Bazelot, M.; Barbara, G.S.; Paute, S.; Gauthier, M.; Raymond-Delpech, V. Homomeric RDL and heteromeric RDL/LCCH3 GABA receptors in the honeybee antennal lobes: Two candidates for inhibitory transmission in olfactory processing. J. Neurophysiol. 2010, 103, 458–468. [Google Scholar] [CrossRef] [PubMed]
- McGonigle, I.; Lummis, S.C.R. Molecular characterization of agonists that bind to an insect GABA receptor. Biochemistry 2010, 49, 2897–2902. [Google Scholar] [CrossRef] [PubMed]
- Ashby, J.A.; McGonigle, I.V.; Price, K.L.; Cohen, N.; Comitani, F.; Dougherty, D.A.; Molteni, C.; Lummis, S.C. GABA binding to an insect GABA receptor: A molecular dynamics and mutagenesis study. Biophys. J. 2012, 103, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Ffrench-Constant, R.H.; Mortlock, D.P.; Shaffer, C.D.; MacIntyre, R.J.; Roush, R.T. Molecular cloning and transformation of cyclodiene resistance in Drosophila: An invertebrate gamma-aminobutyric acid subtype A receptor locus. Proc. Natl. Acad. Sci. USA 1991, 88, 7209–7213. [Google Scholar] [CrossRef] [PubMed]
- Belelli, D.; Callachan, H.; Hill-Venning, C.; Peters, J.A.; Lambert, J.J. Interaction of positive allosteric modulators with human and Drosophila recombinant GABA receptors expressed in Xenopus laevis oocytes. Br. J. Pharmacol. 1996, 118, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Hosie, A.M.; Aronstein, K.; Sattelle, D.B.; Ffrench-Constant, R.H. Molecular biology of insect neuronal GABA receptors. Trends Neurosci. 1997, 20, 578–583. [Google Scholar] [CrossRef]
- Carvalho, K.S.; Silva, S.L.; de Souza, I.A.; Gualberto, S.A.; da Cruz, R.C.; Dos Santos, F.R.; de Carvalho, M.G. Toxicological evaluation of essential oil from the leaves of Croton tetradenius (Euphorbiaceae) on Aedes aegypti and Mus musculus. Parasitol. Res. 2016, 115, 3441–3448. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, F.; Khan, Z.U.; Manzoor, F.; Jamil, M. Analysis of Insect toxicity and repellent activity of Phytochemicals from “Skimmia laureola, Nair” against “Black garden ant, Lasius niger” of Pakistan. Pak. J. Pharm. Sci. 2016, 29, 789–793. [Google Scholar] [PubMed]
- Turchen, L.M.; Piton, L.P.; Dall’Oglio, E.L.; Butnariu, A.R.; Pereira, M.J. Toxicity of Piper aduncum (Piperaceae) Essential Oil Against Euschistus heros (F.) (Hemiptera: Pentatomidae) and Non-Effect on Egg Parasitoids. Neotrop. Entomol. 2016, 45, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Waliwitiya, R.; Belton, P.; Nicholson, R.A.; Lowenberger, C.A. Effects of the essential oil constituent thymol and other neuroactive chemicals on flight motor activity and wing beat frequency in the blowfly Phaenicia sericata. Pest Manag. Sci. 2010, 66, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Nathanson, J.A. Octopamine receptors, adenosine 3′,5′-monophosphate, and neural control of firefly flashing. Science 1979, 203, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Orchard, I.; Carlisle, J.A.; Loughton, B.G.; Gole, J.W.; Downer, R.G. In vitro studies on the effects of octopamine on locust fat body. Gen. Comp. Endocrinol. 1982, 48, 7–13. [Google Scholar] [CrossRef]
- Lange, A.B.; Orchard, I. Identified octopaminergic neurons modulate contractions of locust visceral muscle via adenosine 3′,5′-monophosphate (cyclic AMP). Brain Res. 1986, 363, 340–349. [Google Scholar] [CrossRef]
- Perić-Mataruga, V.; Nenadovic, V.; Ivanovic, J. Neurohormones in insect stress: A review. Arch. Biol. Sci. 2006, 58, 1–12. [Google Scholar] [CrossRef]
- Davenport, A.P.; Evans, P.D. Stress-induced changes in the octopamine levels of insect haemolymph. Insect Biochem. 1984, 14, 135–143. [Google Scholar] [CrossRef]
- Kozanek, M.; Jurani, M.; Somogyiova, E. Influence of social stress on monoamine concentration in the central nervous system of the cockroach Nauphoeta cinerea (Blattodea). Acta Entomol. Bohemoslov. 1986, 83, 171–178. [Google Scholar]
- Zhou, C.; Rao, Y.; Rao, Y. A subset of octopaminergic neurons are important for Drosophila aggression. Nat. Neurosci. 2008, 11, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, T. Review of octopamine in insect nervous systems. Insect Physiol. 2012, 4, 1–17. [Google Scholar] [CrossRef]
- Evans, P.D.; Maqueira, B. Insect octopamine receptors: A new classification scheme based on studies of cloned Drosophila G-protein coupled receptors. Invertebr. Neurosci. 2005, 5, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.F.; Yao, Y.; Huang, J.; Ye, G.Y. Characterization of a β-adrenergic-like octopamine receptor from the rice stem borer (Chilo suppressalis). J. Exp. Biol. 2012, 215, 2646–2652. [Google Scholar] [CrossRef] [PubMed]
- Cazzamali, G.; Klaerke, D.A.; Grimmelikhuijzen, C.J.P. A new family of insect tyramine receptors. Biochem. Biophys. Res. Commun. 2005, 338, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, A.; Roselli, G.; Evans, P.D. A comparison of the signaling properties of two tyramine receptors from Drosophila. J. Neurochem. 2013, 125, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Enan, E.E. Insecticidal activity of essenial oils: Octopaminergic sites of action. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 325–337. [Google Scholar] [CrossRef]
- Pan, C.; Li, W.; Wang, Y.; Jiang, S. Octopamine levels in Blattella germanica L. tissues by capillary gas chromatography with electron capture detection. Int. J. Mol. Sci. 2005, 6, 188–197. [Google Scholar]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Enan, E.E. Molecular response of Drosophila melanogaster tyramine receptor cascade to plant essential oils. Insect Biochem. Mol. Biol. 2005, 35, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Grifman, M.; Arbel, A.; Ginzberg, D.; Glick, D.; Elgavish, S.; Shaanan, B.; Soreq, H. In vitro phosphorylation of acetylcholinesterase at non-consensus protein kinase A sites enhances the rate of acetylcholine hydrolysis. Brain Res. Mol. Brain Res. 1997, 51, 179–187. [Google Scholar] [CrossRef]
- Kaufmann, D.; Dogra, A.K.; Wink, M. Myrtenal inhibits acetylcholinesterase, a known Alzheimer target. J. Pharm. Pharmacol. 2011, 63, 1368–1371. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, M.; Yagi, N.; Miyazawa, M. Acetylcholinesterase inhibitory activity of volatile oil from Peltophorum dasyrachis Kurz ex Bakar (Yellow Batai) and bisabolane-type sesquiterpenoids. J. Agric. Food Chem. 2010, 58, 2824–2829. [Google Scholar] [CrossRef] [PubMed]
- Dohi, S.; Terasaki, M.; Makino, M. Acetylcholinesterase inhibitory activity and chemical composition of commercial essential oils. J. Agric. Food Chem. 2009, 57, 4313–4318. [Google Scholar] [CrossRef] [PubMed]
- Menichini, F.; Tundis, R.; Loizzo, M.R.; Bonesi, M.; Marrelli, M.; Statti, G.A.; Menichini, F.; Conforti, F. Acetylcholinesterase and butyrylcholinesterase inhibition of ethanolic extract and monoterpenes from Pimpinella anisoides V Brig. (Apiaceae). Fitoterapia 2009, 80, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Aazza, S.; Lyoussi, B.; Miguel, M.G. Antioxidant and antiacetylcholinesterase activities of some commercial essential oils and their major compounds. Molecules 2011, 16, 7672–7690. [Google Scholar] [CrossRef] [PubMed]
- Bhadra, S.; Mukherjee, P.K.; Kumar, N.S.; Bandyopadhyay, A. Anticholinesterase activity of standardized extract of Illicium verum Hook. f. fruits. Fitoterapia 2011, 82, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Mata, A.T.; Proença, C.; Ferreira, A.R.; Serralheiro, M.L.M.; Nogueira, J.M.F.; Araújo, M.E.M. Antioxidant and antiacetylcholinesterase activities of five plants used as Portuguese food spices. Food Chem. 2007, 103, 778–786. [Google Scholar] [CrossRef]
- Bonesi, M.; Menichini, F.; Tundis, R. Acetylcholinesterase and butyrylcholinesterase inhibitory activity of Pinus species essential oils and their constituents. J. Enzym. Inhib. Med. Chem. 2010, 25, 622–628. [Google Scholar] [CrossRef] [PubMed]
- Arruda, M.; Viana, H.; Rainha, N.; Neng, N.R.; Rosa, J.S.; Nogueira, J.M.; Barreto Mdo, C. Anti-acetylcholinesterase and antioxidant activity of essential oils from Hedychium gardnerianum Sheppard ex Ker-Gawl. Molecules 2012, 17, 3082–3092. [Google Scholar] [CrossRef] [PubMed]
- Perry, N.S.; Houghton, P.J.; Theobald, A.; Jenner, P.; Perry, E.K. In vitro inhibition of human erythrocyte acetylcholinesterase by Salvia lavandulaefolia essential oil and constituent terpenes. J. Pharm. Pharmacol. 2000, 52, 895–902. [Google Scholar] [CrossRef] [PubMed]

| No. | Essential Oils Components | AChE Source | IC50 (mM) | Ki (mM) | Reference |
|---|---|---|---|---|---|
| 1 | Anisaldehyde | BxACE-1 from Bursaphelenchus xylophilus | 4.95 | [46] | |
| BxACE-2 from Bursaphelenchus xylophilus | 8.53 | [46] | |||
| BxACE-3 from Bursaphelenchus xylophilus | >50 | [46] | |||
| 2 | Camphene | Blatella germanica | N.A. | [47] | |
| 3 | Camphor | Blatella germanica | N.A. | [47] | |
| 4 | 3-Carene | BxACE-1 from Bursaphelenchus xylophilus | 0.37 | [46] | |
| BxACE-2 from Bursaphelenchus xylophilus | 8.18 | [46] | |||
| BxACE-3 from Bursaphelenchus xylophilus | >50 | [46] | |||
| 5 | Carvacrol | Musca domestica | 0.0012 | [48] | |
| Dermacentor variabilis | 0.0018 | [48] | |||
| Periplaneta americana | 0.0004 | [48] | |||
| Aedes aegypti | 0.0012 | [48] | |||
| Drosophila suzukii | N.A. | [49] | |||
| Sitophilus oryzae | 0.05 | [50] | |||
| 6 | Caryophyllene (humulene) | Blatella germanica | N.A. | [47] | |
| 7 | 1,8-Cineole | Pediculus humanus capitis | 77 | [51] | |
| Sitophilus oryzae | 0.084 | [50] | |||
| 8 | Coniferyl alcohol | BxACE-1 from Bursaphelenchus xylophilus | 1.06 | [46] | |
| BxACE-2 from Bursaphelenchus xylophilus | 1.41 | [46] | |||
| BxACE-3 from Bursaphelenchus xylophilus | 1.13 | [46] | |||
| 9 | Cymene | Sitophilus oryzae | 0.05 | [50] | |
| Drosophila suzukii | N.A. | [49] | |||
| 10 | Estragole (Allylanisole) | Blatella germanica | N.A. | [47] | |
| 11 | Eugenol | Sitophilus oryzae | 0.096 | [50] | |
| 12 | Isoeugenol | Sitophilus oryzae | 0.11 | [50] | |
| 13 | Isosafrole | Sitophilus oryzae | 0.71 | [50] | |
| 14 | Limonene | Sitophilus oryzae | 0.73 | [50] | |
| Reticulitermes speratus Kolbe | 0.95 | [41] | |||
| 15 | Linalool | Sitophilus oryzae | N.A. | [50] | |
| 16 | Methyleugenol | Sitophilus oryzae | 0.051 | [50] | |
| 17 | Menthol | Sitophilus oryzae | 0.048 | [50] | |
| Drosophila suzukii | N.A. | [49] | |||
| 18 | Menthone | Sitophilus oryzae | 0.39 | [50] | |
| Drosophila suzukii | N.A. | [49] | |||
| 19 | Nerolidol | BxACE-1 from Bursaphelenchus xylophilus | 9.98 | [46] | |
| BxACE-2 from Bursaphelenchus xylophilus | 15.28 | [46] | |||
| BxACE-3 from Bursaphelenchus xylophilus | 19.06 | [46] | |||
| 20 | Nootkatone | Musca domestica | >30 | [48] | |
| Dermacentor variabilis | >30 | [48] | |||
| Periplaneta americana | >30 | [48] | |||
| Aedes aegypti | >30 | [48] | |||
| 21 | Ocimene | Japanese termite | 0.96 | [52] | |
| Blatella germanica | N.A. | [47] | |||
| 22 | Perilla aldehyde | Drosophila suzukii | 3.06 | [49] | |
| 23 | Phellandrene | Reticulitermes speratus Kolbe | 4.92 | [41] | |
| Blatella germanica | 2.2 | [47] | |||
| 24 | α-Pinene | Sitophilus oryzae | 0.44 | [50] | |
| BxACE-1 from Bursaphelenchus xylophilus | 0.24 | [46] | |||
| BxACE-2 from Bursaphelenchus xylophilus | 0.64 | [46] | |||
| BxACE-3 from Bursaphelenchus xylophilus | 0.68 | [46] | |||
| Reticulitermes speratus Kolbe | 3 | [41] | |||
| 25 | β-Pinene | BxACE-1 from Bursaphelenchus xylophilus | 3.39 | [46] | |
| BxACE-2 from Bursaphelenchus xylophilus | 18.03 | [46] | |||
| BxACE-3 from Bursaphelenchus xylophilus | >50 | [46] | |||
| Reticulitermes speratus Kolbe | 3.08 | [41] | |||
| Sitophilus oryzae | 0.0028 | [50] | |||
| 26 | α-Terpinene | Sitophilus oryzae | 0.14 | [50] | |
| 27 | α-Terpineol | Sitophilus oryzae | 3.94 | [50] | |
| 28 | β-Thujone | Blatella germanica | N.A. | [47] | |
| 29 | Thymol | Sitophilus oryzae | 0.57 | [50] | |
| Drosophila suzukii | 4.26 | [49] |
| Competitive AChE Inhibitors | Reference | Noncompetitive AChE Inhibitors | Reference |
|---|---|---|---|
| Pulegon | [55] | Gossypol | [55] |
| Citral | [55] | Carvone | [60] |
| Linalool | [55] | Camphor | [60] |
| (−)-Bornyl acetate | [55] | ||
| 1,8-Cineol | [55,57,58] | ||
| Terpinen-4-ol | [57] | ||
| Fenchone | [60] | ||
| γ-Terpinene | [60] | ||
| Menthone | [50] | ||
| Menthol | [50] |
| EO Components | Concentration of EOs Component (mM) | Change of GABA Current | Type of Receptor or Source of Receptor | Literature |
|---|---|---|---|---|
| (−)-Borneol | 0.3 | 350% | α1β2γ2s GABAA | [76] |
| Camphor | 0.3 | 40% (inhibition) | α1β2γ2s GABAA | [76] |
| Carvone | 0.3 | 115% | α1β2γ2s GABAA | [76] |
| cis-Jasmone | 1 | 250% | Bovine GABAA | [79] |
| Geraniol | 1 | 500% | α1β1GABAA | [79] |
| (+)-Isomenthol | 1 | 327% | α1β2γ2s GABAA | [78] |
| (+)-Isopulegol | 0.3 | 380% | α1β2γ2s GABAA | [78] |
| Linalool | 1 | 350% | α1β1GABAA | [79] |
| Nerolidol | 1 | 150% | α1β1GABAA | [79] |
| Menthol | 0.32 | 200% | α1β2γ2s GABAA | [78] |
| (+)-Menthol | 0.1 | 596% | α1β2γ2s GABAA | [76] |
| (−)-Menthol | 0.3 | 600% | α1β2γ2s GABAA | [76] |
| (−)-Menthone | 0.3 | 150% | α1β2γ2s GABAA | [76] |
| Methyleugenol | 0.03 | 280% | hippocampal neurons | [80] |
| Methyl jasmonate | 1 | 230% | Bovine GABAA | [79] |
| α-Terpineol | 1 | 299% | α1β2γ2s GABAA | [78] |
| α/β-Thujone | 0.3 | 40% (inhibition) | α1β2γ2s GABAA | [76] |
| 0.1 | 715% | Drosophila melanogaster homomeric RDLac GABAr | [76] | |
| α-Thujone | 0.00066 | 208% | Rat GABAA | [81] |
| 0.003 | 70% (inhibition) | rat dorsal root ganglion neurons | [82] | |
| Thymol | 0.1 | 416% | α1β3γ2s GABAA | [75] |
| 0.01 | 150% | α1β1γ2s GABAA | [75] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review. Molecules 2018, 23, 34. https://doi.org/10.3390/molecules23010034
Jankowska M, Rogalska J, Wyszkowska J, Stankiewicz M. Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review. Molecules. 2018; 23(1):34. https://doi.org/10.3390/molecules23010034
Chicago/Turabian StyleJankowska, Milena, Justyna Rogalska, Joanna Wyszkowska, and Maria Stankiewicz. 2018. "Molecular Targets for Components of Essential Oils in the Insect Nervous System—A Review" Molecules 23, no. 1: 34. https://doi.org/10.3390/molecules23010034