
Selenium and Sulfur to Produce Allium Functional Crops


 ,
,  ,
, 
Abstract
:1. Introduction
2. Absorption and Metabolism of Sulfur and Selenium in Allium
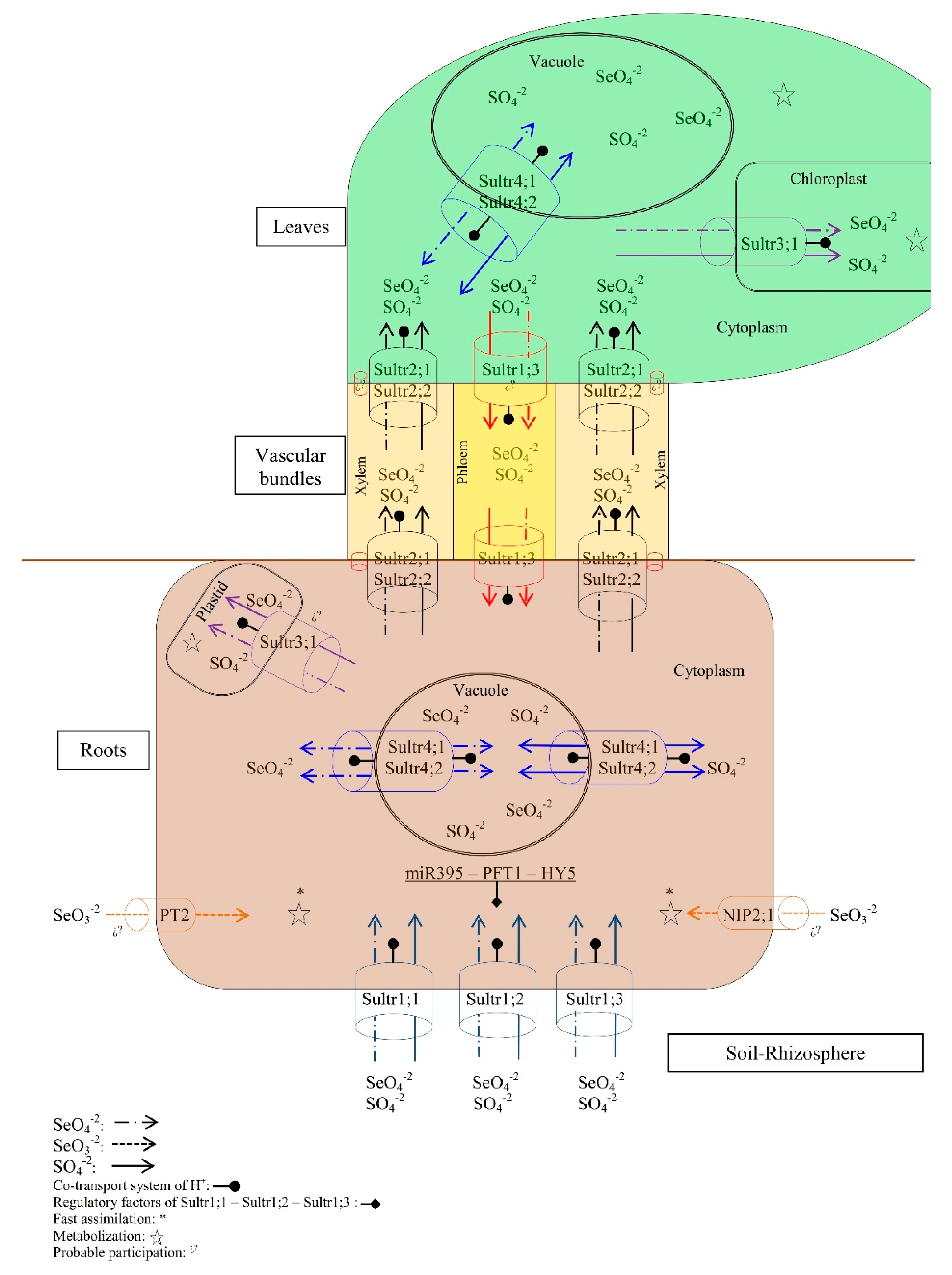
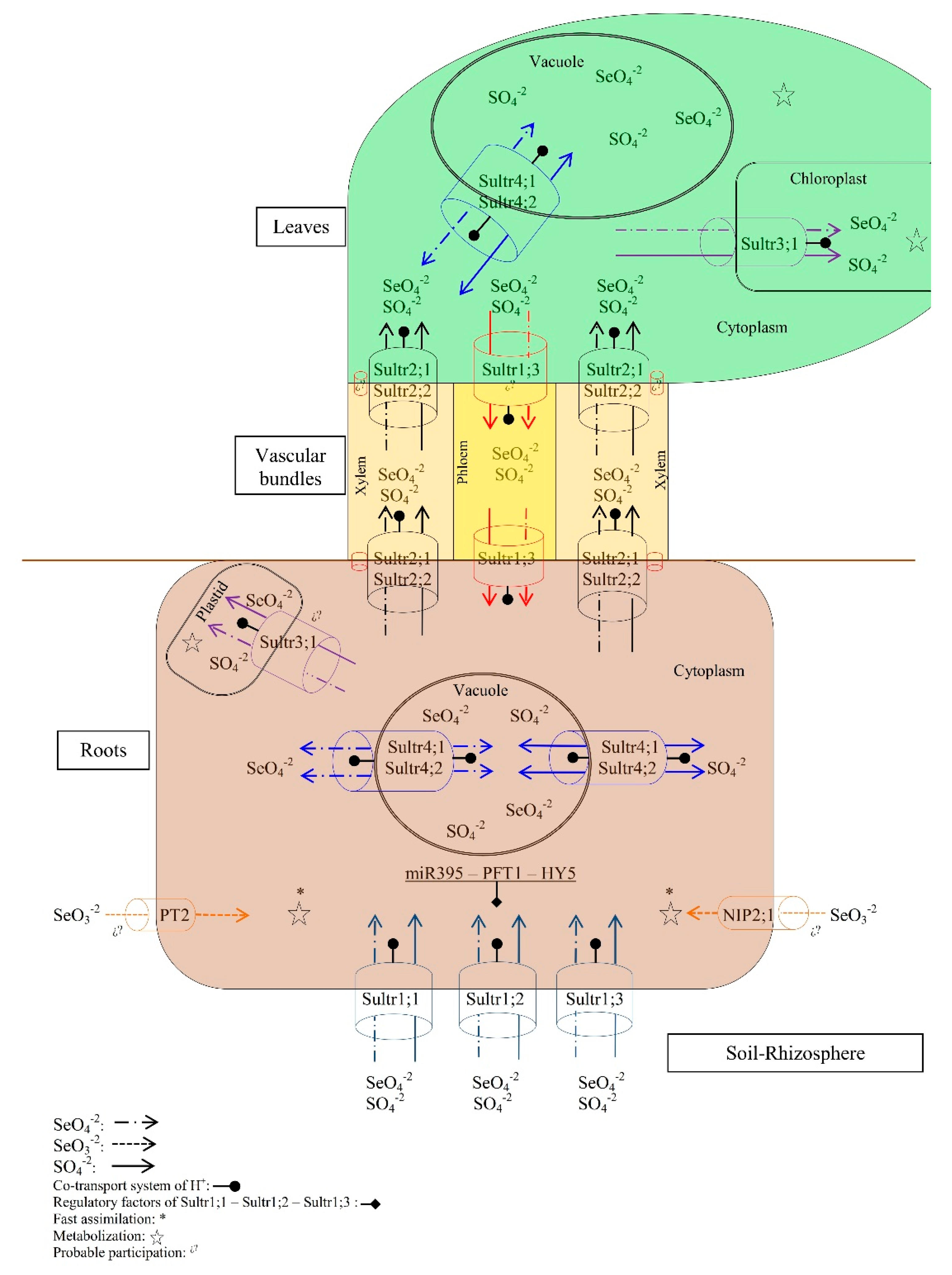
2.1. Absorption
2.2. Transport
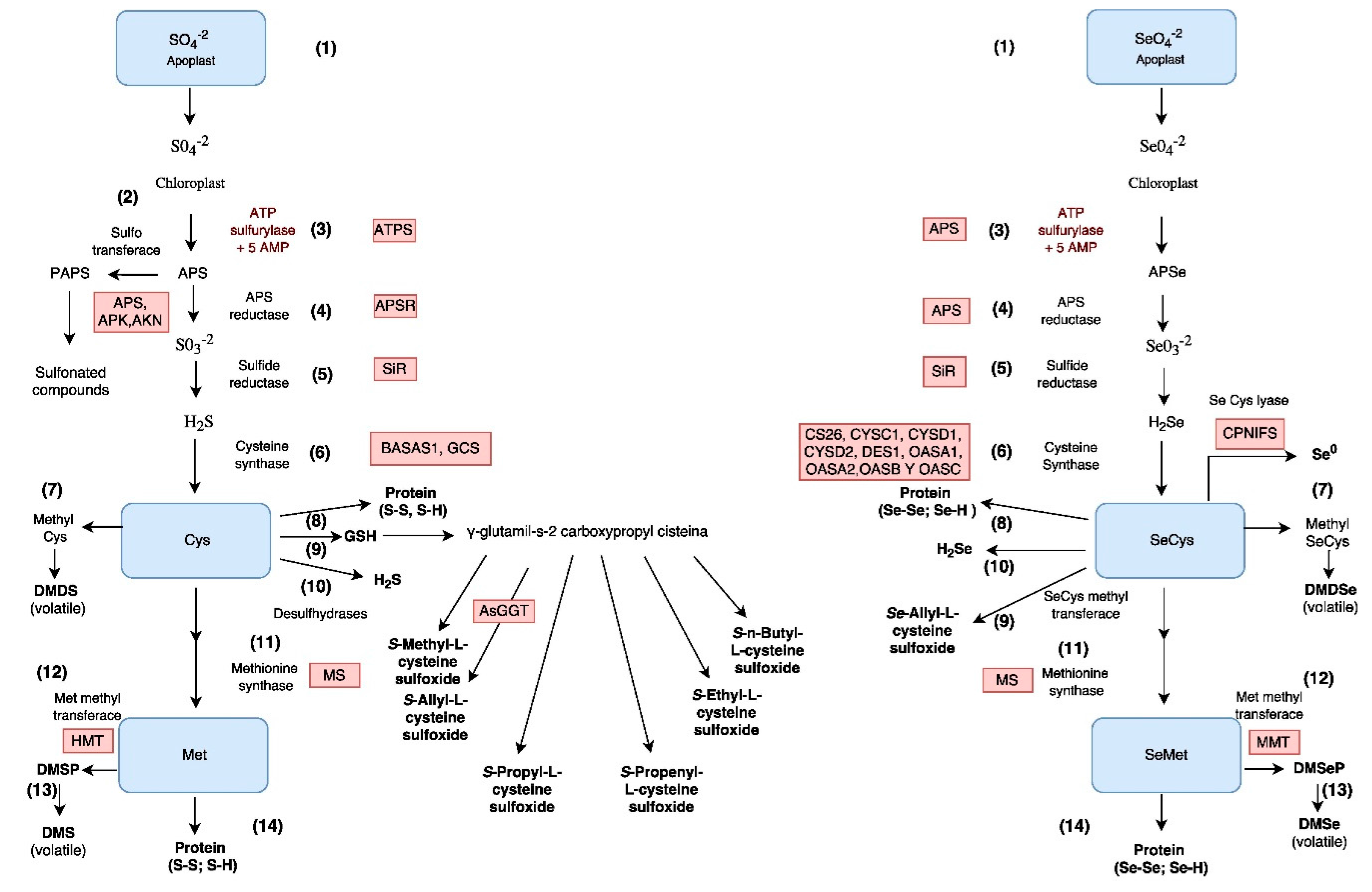
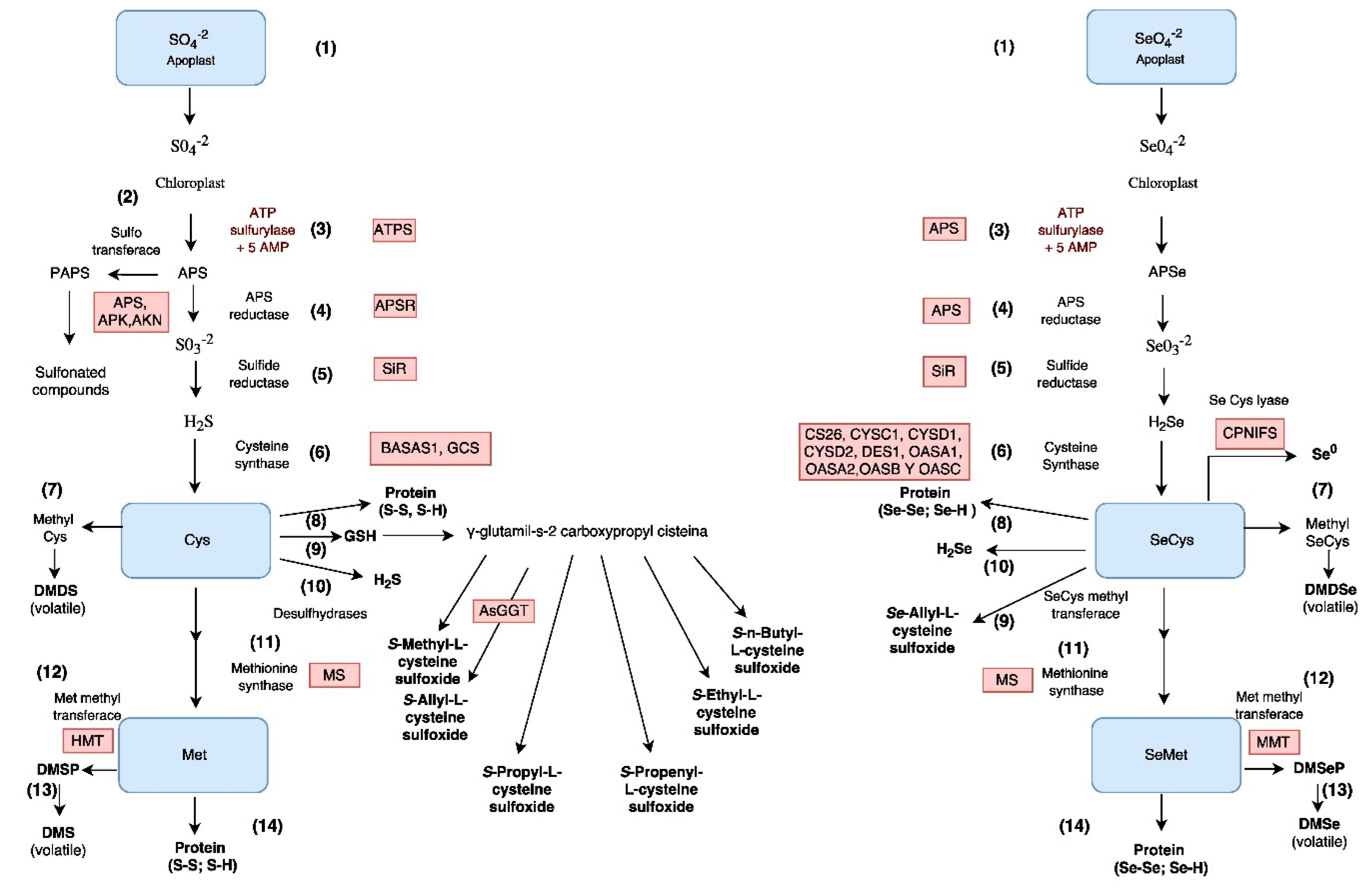
2.3. Assimilation
3. Phytochemicals of Allium spp. Derived of Se and S
Impact of Se and S on the Nutritional and Functional Quality of Allium spp.
4. Use of Selenium and Sulfur in Allium Agricultural Production
Use of Se and S in Allium Production Systems
5. Conclusions
Author Contributions
Conflicts of Interest
References
- Benton Jones, J. Plant Nutrition and Soil Fertility Manual; CRC Press: Boca Raton, FL, USA, 2012; Volume 2, p. 273. [Google Scholar]
- Haraguchi, H. Metallomics as integrated biometal science. J. Anal. At. Spectrom. 2004, 19, 5–14. [Google Scholar] [CrossRef]
- Koca, A.; Koca, I.; Tekguler, B. Two antoxidant elements of Allium vegetables: Germanium and Selenium. Acta Hortic. 2016, 297–302. [Google Scholar] [CrossRef]
- Reilly, K.; Valverde, J.; Finn, L.; Gaffney, M.; Rai, D.K.; Brunton, N. A note on the effectiveness of Selenium supplementation of Irish-grown Allium crops. Irish J. Agric. Food Res. 2014, 53, 91–99. [Google Scholar]
- Pilon-Smits, E.A.H.; Bañuelos, G.S.; Parker, D.R. Uptake, Metabolism, and Volatilization of Selenium by Terrestrial Plants. In Salinity and Drainage in San Joaquin Valley, California: Science, Technology, and Policy; Chang, A.C., Brawer Silva, D., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 147–164. [Google Scholar]
- Slekovec, M.; Goessler, W. Accumulation of Selenium in natural plants and Selenium supplemented vegetable and Selenium speciation by HPLC-ICPMS. Chem. Speciat. Bioavailab. 2005, 17, 63–73. [Google Scholar] [CrossRef]
- Cooke, J.; DeGabriel, J.L. Editorial: Plant silicon interactions between organisms and the implications for ecosystems. Front. Plant Sci. 2016, 7, 1001. [Google Scholar] [CrossRef] [PubMed]
- Medrano-Macías, J.; Leija-Martínez, P.; González-Morales, S.; Juárez-Maldonado, A.; Benavides-Mendoza, A. Use of iodine to biofortify and promote growth and stress tolerance in crops. Front. Plant Sci. 2016, 7, 1146. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; Mcgrath, S.P.; Seneviratne, S.I.; Smith, P.; Winkel, L.H. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2843–2853. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, G.; Gibbons, S. Antibacterial activity of two canthin-6-one alkaloids from Allium neapolitanum. Phyther. Res. 2007, 21, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions—A global benefit to health. Phyther. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Lundegårdh, B.; Botek, P.; Schulzov, V.; Hajšlov, J.; Strömberg, A.; Andersson, H.C. Impact of different green manures on the content of S-alk(en)yl-l-cysteine sulfoxides and l-ascorbic acid in leek (Allium porrum). J. Agric. Food Chem. 2008, 56, 2102–2111. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.R.; White, P.J.; Bryson, R.J.; Meacham, M.C.; Bowen, H.C.; Johnson, S.E.; Hawkesford, M.J.; McGrath, S.P.; Zhao, F.-J.; Breward, N.; et al. Biofortification of UK food crops with Selenium. Proc. Nutr. Soc. 2006, 65, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Meija, J.; Montes-Bayón, M.; Le Duc, D.L.; Terry, N.; Caruso, J.A. Simultaneous monitoring of volatile selenium and sulfur species from Se accumulating plants (Wild type and genetically modified) by GC/MS and GC/ICPMS using solid-phase microextraction for sample introduction. Anal. Chem. 2002, 74, 5837–5844. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.P.; Pilon-Smits, E.A.H.; Lytle, C.M.; Hwang, S.; Tai, J.; Honma, T.S.U.; Yeh, L.; Terry, N. Rate-limiting steps in Selenium assimilation and volatilization by Indian mustard. Plant Physiol. 1998, 117, 1487–1494. [Google Scholar] [CrossRef] [PubMed]
- Terry, N.; Carlson, C.; Raab, T.K.; Zayed, A.M. Rates of Selenium volatilization among crop species. J. Environ. Qual. 1992, 21, 341–344. [Google Scholar] [CrossRef]
- Zayed, A.M.; Terry, N. Selenium volatilization in broccoli as influenced by sulfate supply. J. Plant Physiol. 1992, 140, 646–652. [Google Scholar] [CrossRef]
- Barak, P.; Goldman, I.L. Antagonistic relationship between selenate and sulfate uptake in onion (Allium cepa): Implications for the production of organosulfur and organoselenium compounds in plants. J. Agric. Food Chem. 1997, 45, 1290–1294. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Stroud, J.L.; Broadley, M.R.; Foot, I.; Fairweather-Tait, S.J.; Hart, D.J.; Hurst, R.; Knott, P.; Mowat, H.; Norman, K.; Scott, P.; et al. Soil factors affecting Selenium concentration in wheat grain and the fate and speciation of Se fertilisers applied to soil. Plant Soil 2010, 332, 19–30. [Google Scholar] [CrossRef]
- Hu, Q.; Chen, L.; Xu, J.; Zhang, Y.; Pan, G. Determination of Selenium concentration in rice and the effect of foliar application of Se-enriched fertiliser or sodium selenite on the Selenium content of rice. J. Sci. Food Agric. 2002, 82, 869–872. [Google Scholar] [CrossRef]
- Kotrebai, M.; Birringer, M.; Tyson, J.F.; Block, E.; Uden, P.C. Selenium speciation in enriched and natural samples by HPLC-ICP-MS and HPLC-ESI-MS with perfluorinated carboxylic acid ion-pairing agents. Analyst 2000, 125, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Izgi, B.; Gucer, S.; Jaćimović, R. Determination of Selenium in garlic (Allium sativum) and onion (Allium cepa) by electro thermal atomic absorption spectrometry. Food Chem. 2006, 99, 630–637. [Google Scholar] [CrossRef]
- Giacosa, A.; Faliva, M.; Perna, S.; Minoia, C.; Ronchi, A.; Rondanelli, M. Selenium fortification of an italian rice cultivar via foliar fertilization with sodium selenate and its effects on human serum Selenium levels and on erythrocyte glutathione peroxidase activity. Nutrients 2014, 6, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Lisk, D.J. Enrichment of Selenium in allium vegetables for cancer prevention. Carcinogenesis 1994, 15, 1881–1885. [Google Scholar] [CrossRef] [PubMed]
- Whanger, P.D.; Ip, C.; Polan, C.E.; Uden, P.C.; Welbaum, G. Tumorigenesis, metabolism, speciation, bioavailability, and tissue deposition of Selenium in Selenium-enriched ramps (Allium tricoccum). J. Agric. Food Chem. 2000, 48, 5723–5730. [Google Scholar] [CrossRef] [PubMed]
- Haneklaus, S.; Bloem, E.; Schnug, E.; De Kok, L.J.; Stulen, I. Sulfur. In Handbook of Plant Nutrition; Barker, A.V., Pilbeam, D.J., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 183–238. [Google Scholar]
- Minard, H.R.G. Effect of clove size, spacing, fertilisers, and lime on yield and nutrient content of garlic (Allium sativum). N. Z. J. Exp. Agric. 1978, 6, 139–143. [Google Scholar] [CrossRef]
- Randle, W.M.; Kopsell, D.E.; Kopsell, D.A.; Snyder, R.L. Total sulfur and sulfate accumulation in onion is affected by sulfur fertility. J. Plant Nutr. 1999, 22, 45–51. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2011. [Google Scholar]
- Qureshi, A.A.; Lawande, K.E.; Patil, V.B.; Mani, S. Relationship between Selenium and Sulfur assimilation and resultant interaction on quality parameters in onion. Commun. Soil Sci. Plant Anal. 2012, 43, 1625–1631. [Google Scholar] [CrossRef]
- El-Ramady, H.R.; Domokos-Szabolcsy, É.; Shalaby, T.A.; Prokisch, J.; Fári, M. Selenium in agriculture: Water, air, soil, plants, food, animals and nanoselenium. In CO2 Sequestration, Biofuels and Depollution; Lichtfouse, E., Schwarzbauer, J., Robert, D., Eds.; Springer: Cham, Switzerland, 2015; pp. 153–232. [Google Scholar]
- Suzuki, I. Oxidation of inorganic sulfur compounds: Chemical and enzymatic reactions. Can. J. Microbiol. 1999, 45, 97–105. [Google Scholar] [CrossRef]
- Droux, M. Sulfur assimilation and the role of Sulfur in plant metabolism: A survey. Photosynth. Res. 2004, 79, 331–348. [Google Scholar] [CrossRef] [PubMed]
- Kertesz, M.A.; Mirleau, P. The role of soil microbes in plant Sulphur nutrition. J. Exp. Bot. 2004, 55, 1939–1945. [Google Scholar] [CrossRef] [PubMed]
- Lucheta, A.; Lambais, M. Sulfur in agriculture. Rev. Bras. Ciência do Solo 2012, 36, 1369–1379. [Google Scholar] [CrossRef]
- Griffith, C.M.; Woodrow, J.E.; Seiber, J.N. Environmental behavior and analysis of agricultural Sulfur. Pest Manag. Sci. 2015, 71, 1486–1496. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J.; et al. Interactions between Selenium and sSlphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [PubMed]
- El-Ramady, H.; Abdalla, N.; Alshaal, T.; El-Henawy, A.; Faizy, S.E.D.A.; Shams, M.H.; Shalaby, T.; Bayoumi, Y.; Elhawat, N.; Shehata, S.; et al. Selenium and its role in higher plants. In Pollutants in Buildings, Water and Living Organisms; Lichtfouse, E., Schwarzbauer, J., Robert, D., Eds.; Springer: Cham, Switzerland, 2015; pp. 235–296. [Google Scholar]
- Terry, N.; Zayed, A.M.; de Souza, M.P.; Tarun, A. Selenium in higher plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed]
- Arnault, I.; Auger, J. Seleno-compounds in garlic and onion. J. Chromatogr. A 2006, 1112, 23–30. [Google Scholar] [CrossRef] [PubMed]
- El-Ramady, H.; Abdalla, N.; Taha, H.S.; Alshaal, T.; El-Henawy, A.; Faizy, S.E.D.A.; Shams, M.S.; Youssef, S.M.; Shalaby, T.; Bayoumi, Y.; et al. Selenium and nano-selenium in plant nutrition. Environ. Chem. Lett. 2016, 14, 123–147. [Google Scholar] [CrossRef]
- Buchner, P. Plant sulfate transporters. In Plant Membrane and Vacuolar Transporters; Jaiwal, P.K., Singh, R.P., Dhankher, O.P., Eds.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Hawkesford, M.J. Transporter gene families in plants: The sulphate transporter gene family—Redundancy or specialization? Physiol. Plant. 2003, 117, 155–163. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; De Kok, L.J. Managing Sulphur metabolism in plants. Plant Cell Environ. 2006, 29, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Maruyama-Nakashita, A.; Inoue, E.; Watanabe-Takahashi, A.; Yamaya, T.; Takahashi, H. Transcriptome profiling of Sulfur-responsive genes in Arabidopsis reveals global effects of Sulfur nutrition on multiple metabolic pathways. Plant Physiol. 2003, 132, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Buchner, P.; Yoshimoto, N.; Hawkesford, M.J.; Shiu, S.-H. Evolutionary relationships and functional diversity of plant sulfate transporters. Front. Plant Sci. 2012, 2, 119. [Google Scholar] [CrossRef] [PubMed]
- Shibagaki, N.; Rose, A.; McDermott, J.P.; Fujiwara, T.; Hayashi, H.; Yoneyama, T.; Davies, J.P. Selenate-resistant mutants of Arabidopsis thaliana identify Sultr1;2, a sulfate transporter required for efficient transport of sulfate into roots. Plant J. 2002, 29, 475–486. [Google Scholar] [CrossRef] [PubMed]
- Van Hoewyk, D.; Takahashi, H.; Inoue, E.; Hess, A.; Tamaoki, M.; Pilon-Smits, E.A.H. Transcriptome analyses give insights into Selenium-stress responses and Selenium tolerance mechanisms in Arabidopsis. Physiol. Plant. 2008, 132, 236–253. [Google Scholar] [CrossRef] [PubMed]
- Kopsell, D.A.; Randle, W.M. Selenium affects the S-alk(en)yl cysteine sulfoxides among short-day onion cultivars. J. Am. Soc. Hortic. Sci. 1999, 124, 307–311. [Google Scholar]
- Cherest, H.; Davidian, J.-C.; Thomas, D.; Benes, V.; Ansorge, W.; Surdin-Kerjan, Y. Molecular characterization of two high affinity sulfate transporters in Saccharomyces cerevisiae. Genetics 1997, 145, 627–635. [Google Scholar] [PubMed]
- Takahashi, H.; Watanabe-Takahashi, A.; Smith, F.W.; Blake-Kalff, M.; Hawkesford, M.J.; Saito, K. The roles of three functional sulphate transporters involved in uptake and translocation of sulphate in Arabidopsis thaliana. Plant J. 2000, 23, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, N.; Takahashi, H.; Smith, F.W.; Yamaya, T.; Saito, K. Two distinct high-affinity sulfate transporters with different inducibilities mediate uptake of sulfate in Arabidopsis roots. Plant J. 2002, 29, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, T.; Watanabe-Takahashi, A.; Hayashi, N.; Ohnishi, M.; Mimura, T.; Buchner, P.; Hawkesford, M.J.; Yamaya, T.; Takahashi, H. Vacuolar sulfate transporters are essential determinants controlling internal distribution of sulfate in Arabidopsis. Plant Cell 2004, 16, 2693–2704. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, N.; Inoue, E.; Saito, K.; Yamaya, T.; Takahashi, H. Phloem-localizing sulfate transporter, Sultr1;3, mediates re-distribution of Sulfur from source to sink organs in Arabidopsis. Plant Physiol. 2003, 131, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Kopriva, S.; Calderwood, A.; Weckopp, S.C.; Koprivova, A. Plant Sulfur and Big Data. Plant Sci. 2015, 241, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Bell, C.I.; Tremblay, N.; Dorais, M.; Gosselin, A. Uptake and translocation of sulphate in tomato seedlings in relation to sulphate supply. J. Plant Nutr. 2002, 25, 1471–1485. [Google Scholar] [CrossRef]
- Briggs, W.H.; Goldman, I.L. Variation in economically and ecologically important traits in onion plant organs during reproductive development. Plant. Cell Environ. 2002, 25, 1031–1037. [Google Scholar] [CrossRef]
- Durenkamp, M.; De Kok, L.J. Impact of pedospheric and atmospheric Sulphur nutrition on Sulphur metabolism of Allium cepa L., a species with a potential sink capacity for secondary Sulphur compounds. J. Exp. Bot. 2004, 55, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Pandey, C.; Gupta, M. Selenium and auxin mitigates Arsenic stress in rice (Oryza sativa L.) by combining the role of stress indicators, modulators and genotoxicity assay. J. Hazard. Mater. 2015, 287, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Pazurkiewicz-Kocot, K.; Kita, A.; Pietruszka, M. Effect of Selenium on Magnesium, Iron, Manganese, Copper, and Zinc accumulation in corn treated by Indole-3-acetic acid. Commun. Soil Sci. Plant Anal. 2008, 39, 2303–2318. [Google Scholar] [CrossRef]
- Cao, M.J.; Wang, Z.; Wirtz, M.; Hell, R.; Oliver, D.J.; Xiang, C. Bin SULTR3;1 is a chloroplast-localized sulfate transporter in Arabidopsis thaliana. Plant J. 2013, 73, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Cao, M.J.; Wang, Z.; Zhao, Q.; Mao, J.L.; Speiser, A.; Wirtz, M.; Hell, R.; Zhu, J.K.; Xiang, C. Bin Sulfate availability affects ABA levels and germination response to ABA and salt stress in Arabidopsis thaliana. Plant J. 2014, 77, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Bulska, E.; Wierzbicka, I.A.; Wysocka, M.H.; Proost, K.; Janssens, K.; Falkenberg, G. In vivo investigation of the distribution and the local speciation of Selenium in Allium cepa L. by means of microscopic X-ray Absorption Near-Edge Structure Spectroscopy and Confocal Microscopic X-ray Fluorescence Analysis. Anal. Chem. 2006, 78, 7616–7624. [Google Scholar] [CrossRef] [PubMed]
- Lavu, R.V.S.; Du Laing, G.; Van De Wiele, T.; Pratti, V.L.; Willekens, K.; Vandecasteele, B.; Tack, F. Fertilizing soil with Selenium fertilizers: Impact on concentration, speciation, and bioaccessibility of Selenium in leek (Allium ampeloprasum). J. Agric. Food Chem. 2012, 60, 10930–10935. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.D.; Yu, X.H.; Zhou, S.M.; Liu, S.Q. De novo assembly and characterization of the welsh onion (Allium fistulosum L.) transcriptome using Illumina technology. Mol. Genet. Genom. 2016, 291, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Chao, D.-Y.; Baraniecka, P.; Danku, J.; Koprivova, A.; Lahner, B.; Luo, H.; Yakubova, E.; Dilkes, B.; Kopriva, S.; Salt, D.E. Variation in sulfur and selenium accumulation is controlled by naturally occurring isoforms of the key Sulfur assimilation enzyme ADENOSINE 5’-PHOSPHOSULFATE REDUCTASE2 across the Arabidopsis species range. Plant Physiol. 2014, 166, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- McCallum, J.A.; Pither-Joyce, M.; Shaw, M. Sulfur deprivation and genotype affect gene expression and metabolism of onion roots. J. Am. Soc. Hortic. Sci. 2002, 127, 583–589. [Google Scholar]
- McManus, M.T.; Joshi, S.; Searle, B.; Pither-Joyce, M.; Shaw, M.; Leung, S.; Albert, N.; Shigyo, M.; Jakse, J.; Havey, M.J.; et al. Genotypic variation in Sulfur assimilation and metabolism of onion (Allium cepa L.) III. Characterization of sulfite reductase. Phytochemistry 2012, 83, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Feldman-Salit, A.; Wirtz, M.; Hell, R.; Wade, R.C. A mechanistic model of the Cysteine Synthase Complex. J. Mol. Biol. 2009, 386, 37–59. [Google Scholar] [CrossRef] [PubMed]
- Novoselov, S.V.; Rao, M.; Onoshko, N.V.; Zhi, H.; Kryukov, G.V.; Xiang, Y.; Weeks, D.P.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins and selenocysteine insertion system in the model plant cell system, Chlamydomonas reinhardtii. EMBO J. 2002, 21, 3681–3693. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.G.; Hughes, J.; Tregova, A.; Milne, J.; Tomsett, A.B.; Collin, H.A. Biosynthesis of the flavour precursors of onion and garlic. J. Exp. Bot. 2004, 55, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.; Tregova, A.; Tomsett, A.B.; Jones, M.G.; Cosstick, R.; Collin, H.A. Synthesis of the flavour precursor, alliin, in garlic tissue cultures. Phytochemistry 2005, 66, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Edmands, W.M.B.; Gooderham, N.J.; Holmes, E.; Mitchell, S.C. S-Methyl-l-cysteine sulphoxide: The Cinderella phytochemical? Toxicol. Res. 2013, 2, 11–22. [Google Scholar] [CrossRef]
- Otte, M.L.; Wilson, G.; Morris, J.T.; Moran, B.M. Dimethylsulphoniopropionate (DMSP) and related compounds in higher plants. J. Exp. Bot. 2004, 55, 1919–1925. [Google Scholar] [CrossRef] [PubMed]
- Charlson, R.J.; Lovelock, J.E.; Andreae, M.O.; Warren, S.G. Oceanic phytoplankton, atmospheric Sulphur, cloud albedo and climate. Nature 1987, 326, 655–661. [Google Scholar] [CrossRef]
- Van Hoewyk, D.; Garifullina, G.F.; Ackley, A.R.; Abdel-Ghany, S.E.; Marcus, M.A.; Fakra, S.; Ishiyama, K.; Inoue, E.; Pilon, M.; Takahashi, H.; et al. Overexpression of AtCpNifS enhances Selenium tolerance and accumulation in Arabidopsis. Plant Physiol. 2005, 139, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Bloem, E.; Riemenschneider, A.; Volker, J.; Papenbrock, J.; Schmidt, A.; Salac, I.; Haneklaus, S.; Schnug, E. Sulphur supply and infection with Pyrenopeziza brassicae influence l-cysteine desulphydrase activity in Brassica napus L. J. Exp. Bot. 2004, 55, 2305. [Google Scholar] [CrossRef] [PubMed]
- Rausch, T.; Wachter, A. Sulfur metabolism: A versatile platform for launching defence operations. Trends Plant Sci. 2005, 10, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Dini, I.; Tenore, G.C.; Dini, A. S-Alkenyl Cysteine Sulfoxide and Its antioxidant properties from Allium cepa var. tropeana (Red Onion) seeds. J. Nat. Prod. 2008, 71, 2036–2037. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.; Whiteman, M.; Moore, K.; Zhun, Y. Bioactive S-alk(en)yl cysteine sulfoxide metabolites in the genus Allium: The chemistry of potential therapeutic agents. Nat. Prod. Rep. 2005, 22, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Dugravot, S.; Brunissen, L.; Létocart, E.; Tjallingii, W.F.; Vincent, C.; Giordanengo, P.; Cherqui, A. Local and systemic responses induced by aphids in Solanum tuberosum plants. Entomol. Exp. Appl. 2007, 123, 271–277. [Google Scholar] [CrossRef]
- Varin, L.; Marsolais, F.; Richard, M.; Rouleau, M. Sulfation and sulfotransferases 6: Biochemistry and molecular biology of plant sulfotransferases. FASEB J. 1997, 11, 517–525. [Google Scholar] [PubMed]
- Riemenschneider, A.; Riedel, K.; Hoefgen, R.; Papenbrock, J.; Hesse, H. Impact of reduced O-Acetylserine(thiol)lyase Isoform contents on potato plant metabolism. Plant Physiol. 2005, 137, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Randle, W.M.; Lancaster, J.E. Sulphur compounds in Alliums in relation to flavour quality. In Allium Crop Science: Recent Advances; Rabinowitch, H.D., Currah, L., Eds.; CAB International: Wallingford, UK, 2002; pp. 329–356. [Google Scholar]
- Yoshimoto, N.; Yabe, A.; Sugino, Y.; Murakami, S.; Sai-Ngam, N.; Sumi, S.-I.; Tsuneyoshi, T.; Saito, K. Garlic γ-glutamyl transpeptidases that catalyze deglutamylation of biosynthetic intermediate of alliin. Front. Plant Sci. 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.; Yang, W.; Arnault, I.; Pannier, F.; Potin-Gautier, M. High-performance liquid chromatographic-inductively coupled plasma mass spectrometric evidence for Se-“alliins” in garlic and onion grown in Se-rich soil. J. Chromatogr. A 2004, 1032, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, R.A. Emission of biogenic hydrogen sulfide. Tellus 1974, 26, 254–260. [Google Scholar] [CrossRef]
- Vairavamurthy, A.; Andreae, M.O.; Iverson, R.L. Biosynthesis of dimethlysulfide and dimethylpropiothetin by Hymenomonas carterae in relation to Sulfur source and salinity variations. Limnol. Oceanogr. 1985, 30, 59–70. [Google Scholar] [CrossRef]
- Calderwood, A.; Kopriva, S. Hydrogen sulfide in plants: From dissipation of excess Sulfur to signaling molecule. Nitric Oxide 2014, 41, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D. The evolutionary ecology of mast seeding. Trends Ecol. Evol. 1994, 9, 465–470. [Google Scholar] [CrossRef]
- Wilson, L.G.; Bressan, R.A.; Filner, P. Light-dependent mmission of Hydrogen Sulfide from plants. Plant Physiol. 1978, 61, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, J.; Wilson, L.G.; Filner, P. Resistance to Injury by Sulfur Dioxide: Correlation with Its Reduction to, and Emission of, Hydrogen Sulfide in Cucurbitaceae. Plant Physiol. 1982, 70, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Bloem, E.; Haneklaus, S.; Salac, I.; Wickenhäuser, P.; Schnug, E. Facts and fiction about Sulfur metabolism in relation to plant-pathogen interactions. Plant Biol. 2007, 9, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Papenbrock, J.; Riemenschneider, A.; Kamp, A.; Schulz-Vogt, H.N.; Schmidt, A. Characterization of cysteine-degrading and H2S-releasing enzymes of higher plants-from the field to the test tube and back. Plant Biol. 2007, 9, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Andreae, M.O. Ocean-atmosphere interactions in the global biogeochemical Sulfur cycle. Mar. Chem. 1990, 30, 1–29. [Google Scholar] [CrossRef]
- Andreae, M.O.; Crutzen, P.J. Atmospheric aerosols: Biogeochemical sources and role in atmospheric chemistry. Science 1997, 276, 1052–1058. [Google Scholar] [CrossRef]
- Faloona, I. Sulfur processing in the marine atmospheric boundary layer: A review and critical assessment of modeling uncertainties. Atmos. Environ. 2009, 43, 2841–2854. [Google Scholar] [CrossRef]
- Beilstein, M.A.; Whanger, P.D.; Yang, G.Q. Chemical forms of Selenium in corn and rice grown in a high Selenium area of China. Biomed. Environ. Sci. BES 1991, 4, 392–398. [Google Scholar] [PubMed]
- Virupaksha, T.K.; Shrift, A. Biochemical differences between Selenium accumulator and non-accumulator astragalus species. Biochim. Biophys. Acta Gen. Subj. 1965, 107, 69–80. [Google Scholar] [CrossRef]
- Wang, Y.; Böck, A.; Neuhierl, B. Acquisition of selenium tolerance by a Selenium non-accumulating Astragalus species via selection. Biofactors 1999, 9, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.G.; Pilon-Smits, E.A.H.; Zhao, F.J.; Williams, P.N.; Meharg, A.A. Selenium in higher plants: Understanding mechanisms for biofortification and phytoremediation. Trends Plant Sci. 2009, 14, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Block, E. The organosulfur chemistry of the genus Allium—Implications for the organic chemistry of Sulfur. Angew. Chem. Int. Ed. Engl. 1992, 31, 1135–1178. [Google Scholar] [CrossRef]
- Block, E.; Gulati, H.; Putman, D.; Sha, D.; You, N.; Zhao, S.-H. Allium chemistry: Synthesis of 1-[alk(en)ylsulfinyl]propyl alk(en)yl disulfides (cepaenes), antithrombotic flavorants from homogenates of onion (Allium cepa). J. Agric. Food Chem. 1997, 45, 4414–4422. [Google Scholar] [CrossRef]
- Wagner, H.; Dorsch, W.; Bayer, T.; Breu, W.; Willer, F. Antiasthmatic effects of onions: Inhibition of 5-lipoxygenase and cyclooxygenase in vitro by thiosulfinates and “Cepaenes”. Prostaglandins Leukot. Essent. Fat. Acids 1990, 39, 59–62. [Google Scholar] [CrossRef]
- Rizwani, G.H.; Shareef, H. Genus Allium: The potential nutritive and therapeutic source. J. Pharm. Nutr. Sci. 2011, 1, 158–163. [Google Scholar] [CrossRef]
- Yun, H.-M.; Ban, J.O.; Park, K.-R.; Lee, C.K.; Jeong, H.-S.; Han, S.B.; Hong, J.T. Potential therapeutic effects of functionally active compounds isolated from garlic. Pharmacol. Ther. 2014, 142, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Miron, T.; Rabinkov, A.; Mirelman, D.; Wilchek, M.; Weiner, L. The mode of action of allicin: Its ready permeability through phospholipid membranes may contribute to its biological activity. Biochim. Biophys. Acta (BBA) Biomembr. 2000, 1463, 20–30. [Google Scholar] [CrossRef]
- Slusarenko, A.J.; Patel, A.; Portz, D. Control of plant diseases by natural products: Allicin from garlic as a case study. Eur. J. Plant Pathol. 2008, 121, 313–322. [Google Scholar] [CrossRef]
- Mahboubi, M.; Kazempour, N. Chemical composition, antioxidant and antimicrobial activity of Allium hirtifolium essential oil. J. Microbiol. Biotechnol. Food Sci. 2014, 3, 402. [Google Scholar]
- Hosono, T.; Hosono-Fukao, T.; Inada, K.; Tanaka, R.; Yamada, H.; Iitsuka, Y.; Seki, T.; Hasegawa, I.; Ariga, T. Alkenyl group is responsible for the disruption of microtubule network formation in human colon cancer cell line HT-29 cells. Carcinogenesis 2008, 29, 1400–1406. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Choi, S.; Johnson, D.E.; Vogel, V.G.; Johnson, C.S.; Trump, D.L.; Lee, Y.J.; Singh, S.V. Diallyl trisulfide-induced apoptosis in human prostate cancer cells involves c-Jun N-terminal kinase and extracellular-signal regulated kinase-mediated phosphorylation of Bcl-2. Oncogene 2004, 23, 5594–5606. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Lew, K.L.; Kim, Y.-A.; Zeng, Y.; Hahm, E.-R.; Dhir, R.; Singh, S.V. Diallyl Trisulfide suppresses growth of PC-3 human prostate cancer Xenograft In vivo in association with Bax and Bak induction. Clin. Cancer Res. 2006, 12, 6836–6843. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Lawson, L.D.; Milner, J.A. Allyl sulfides from garlic suppress the in vitro proliferation of human a549 lung tumor cells. Nutr. Cancer 1997, 29, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Pinto, J.T.; Gundersen, G.G.; Weinstein, I.B. Effects of a series of organosulfur compounds on mitotic arrest and induction of apoptosis in colon cancer cells. Mol. Cancer Ther. 2005, 4, 1388–1398. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, S.G.; Milner, J.A. Diallyl disulfide induces apoptosis of human colon tumor cells. Carcinogenesis 1996, 17, 669. [Google Scholar] [CrossRef] [PubMed]
- Sparnins, V.L.; Barany, G.; Wattenberg, L.W. Effects of organosulfur compounds from garlic and onions on benzo[a]pyrene-induced neoplasia and glutathione S-transferase activity in the mouse. Carcinogenesis 1988, 9, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, S.; Asgary, S.; Moshtaghian, J.; Rafieian, M.; Adelnia, A.; Shamsi, F. Liver-protective effects of hydroalcoholic extract of Allium hirtifolium Boiss. in rats with alloxan-induced diabetes mellitus. ARYA Atheroscler. J. 2010, 6, 11–15. [Google Scholar]
- Huang, Z.; Ren, J.W. Antibacterial activity of elephant garlic and its effect against U2OS human osteosarcoma cells. Iran. J. Basic Med. Sci. 2013, 16, 1088–1094. [Google Scholar] [PubMed]
- El-Shenawy, S.M.; Yassin, N.A.; Badary, O.A.; EL-Moneem, M.; AL-Shafeiy, H.M. Study of the effect of Allium porrum on osteoporosis induced in rats. Der. Pharm. Lett. 2013, 5, 188–198. [Google Scholar]
- Kratchanova, M.; Nikolova, M.; Pavlova, E.; Yanakieva, I.; Kussovski, V. Composition and properties of biologically active pectic polysaccharides from leek (Allium porrum). J. Sci. Food Agric. 2010, 90, 2046–2051. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, E.; Lanzotti, V.; Taglialatela-Scafati, O.; Cicala, C. The flavonoids of leek, Allium porrum. Phytochemistry 2001, 57, 565–569. [Google Scholar] [CrossRef]
- Sedighi, M.; Rafieian-Kopaei, M.; Noori-Ahmadabadi, M. Effect of Allium ampeloprasum on ileum function: Involvement of beta-adrenergic receptors and voltage dependent calcium channels. Life Sci. J. 2012, 9, 1660–1667. [Google Scholar]
- Štajner, D.; Popović, B.M.; Ćalić-Dragosavac, D.; Malenčić, Đ.; Zdravković-Korać, S. Comparative study on Allium schoenoprasum cultivated plant and Allium schoenoprasum tissue culture organs antioxidant status. Phyther. Res. 2011, 25, 1618–1622. [Google Scholar] [CrossRef] [PubMed]
- Nguansangiam, S.; Angsubhakorn, S.; Bhamarapravati, S.; Suksamrarn, A. Effects of elephant garlic volatile oil (Allium ampeloprasum) and T-2 Toxin on Murine Skin. Southeast Asian J. Trop. Med. Public Health 2003, 34, 899–905. [Google Scholar] [PubMed]
- Nasir, A.S. Hepatoprotective and some haematological parameters effect of Allium ampeloprasum against carbon tetrachloride induced liver toxicity in albino rats. Kufa J. Vet. Med. Sci. 2012, 3, 117–126. [Google Scholar]
- Badary, O.A.; Yassin, N.A.Z.; El-Shenawy, S.M.A.; EL-Moneem, M.A.; AL-Shafeiy, H.M. Study of the effect of Allium porrum on hypertension induced in rats. Rev. Latinoam. Química 2013, 41, 149–160. [Google Scholar]
- Rahimi-Madiseh, M.; Heidarian, E.; Kheiri, S.; Rafieian-Kopaei, M. Effect of hydroalcoholic Allium ampeloprasum extract on oxidative stress, diabetes mellitus and dyslipidemia in alloxan-induced diabetic rats. Biomed. Pharmacother. 2017, 86, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Parvu, A.E.; Parvu, M.; Vlase, L.; Miclea, P.; Mot, A.C.; Silaghi-Dumitrescu, R. Anti-inflammatory effects of Allium schoenoprasum L. leaves. J. Physiol. Pharmacol. 2014, 65, 309–315. [Google Scholar] [PubMed]
- Dobhal, Y.; Parcha, V.; Dhasmana, D.C. Cardioprotective potential of Allium humile leaves extract. Orient. Pharm. Exp. Med. 2014, 14, 157–162. [Google Scholar] [CrossRef]
- Dobhal, Y.; Parcha, V.; Dhasmana, D.C. Effect of cardioactive principle of methanolic extract of Allium humile leaves on global ischaemic rat heart. Pharm. Biol. Eval. 2015, 2, 20–23. [Google Scholar]
- Rafieian-kopaei, M.; Keshvari, M.; Asgary, S.; Salimi, M.; Heidarian, E. Potential role of a nutraceutical spice (Allium hirtifolium) in reduction of atherosclerotic plaques. J. HerbMed Pharmacol. 2013, 2, 23–28. [Google Scholar]
- Suzuki, K.T.; Tsuji, Y.; Ohta, Y.; Suzuki, N. Preferential organ distribution of methylselenol source Se-methylselenocysteine relative to methylseleninic acid. Toxicol. Appl. Pharmacol. 2008, 227, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Lisk, D.J. Efficacy of cancer prevention by high-selenium garlic is primarily dependent on the action of Selenium. Carcinogenesis 1995, 16, 2649–2652. [Google Scholar] [CrossRef] [PubMed]
- El-Bayoumy, K.; Sinha, R.; Pinto, J.T.; Rivlin, R.S. Cancer chemoprevention by garlic and garlic-containing Sulfur and Selenium compounds. J. Nutr. 2006, 136, 864S–869S. [Google Scholar] [PubMed]
- Imen, A.; Najjaa, H.; Neffati, M. Influence of sulfur fertilization on S-containing, phenolic, and carbohydrate metabolites in rosy garlic (Allium roseum L.): A wild edible species in North Africa. Eur. Food Res. Technol. 2013, 237, 521–527. [Google Scholar] [CrossRef]
- Zeng, H.; Combs, G.F. Selenium as an anticancer nutrient: Roles in cell proliferation and tumor cell invasion. J. Nutr. Biochem. 2008, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ogra, Y.; Ishiwata, K.; Iwashita, Y.; Suzuki, K.T. Simultaneous speciation of Selenium and Sulfur species in selenized odorless garlic (Allium sativum L. Shiro) and shallot (Allium ascalonicum) by HPLC-inductively coupled plasma-(octopole reaction system)-mass spectrometry and electrospray ionization-tand. J. Chromatogr. A 2005, 1093, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Lisk, D.; Block, E.; Ip, C. Characterization of the biological activity of γ-glutamyl-Se—methylselenocysteine: A novel, naturally occurring anticancer agent from garlic. Cancer Res. 2001, 61, 2923–2928. [Google Scholar] [PubMed]
- Shah, M.; Kannamkumarath, S.S.; Wuilloud, J.C.A.; Wuilloud, R.G.; Caruso, J.A. Identification and characterization of Selenium species in enriched green onion (Allium fistulosum) by HPLC-ICP-MS and ESI-ITMS. J. Anal. At. Spectrom. 2004, 19, 381–386. [Google Scholar] [CrossRef]
- Tsuneyoshi, T.; Yoshida, J.; Sasaoka, T. Hydroponic cultivation offers a practical means of producing Selenium enriched garlic. J. Nutr. 2006, 136, 870–872. [Google Scholar]
- Coolong, T.W.; Randle, W.M. Sulfur and Nitrogen availability interact to affect the flavor biosynthetic pathway in onion. J. Am. Soc. Hortic. Sci. 2003, 128, 776–783. [Google Scholar]
- Forney, C.F.; Jordan, M.A.; Campbell-Palmer, L.; Fillmore, S.; McRae, K.; Best, K. Sulfur fertilization affects onion quality and flavor chemistry during storage. Acta Hortic. 2010, 877, 163–168. [Google Scholar] [CrossRef]
- Kumari, K.; Augusti, K.T. Antidiabetic and antioxidant effects of S -methyl cysteine sulfoxide isolated from onions (Allium cepa Linn) as compared to standard drugs in alloxan diabetic rats. Indian J. Exp. Biol. 2002, 40, 1005–1009. [Google Scholar] [PubMed]
- Bloem, E.; Haneklaus, S.; Schnug, E. Influence of Nitrogen and Sulfur fertilization on the Alliin content of onions and garlic. J. Plant Nutr. 2005, 27, 1827–1839. [Google Scholar] [CrossRef]
- Bloem, E.; Haneklaus, S.; Schnug, E. Influence of fertilizer practices on S-containing metabolites in garlic (Allium sativum L.) under field conditions. J. Agric. Food Chem. 2010, 58, 10690–10696. [Google Scholar] [CrossRef] [PubMed]
- Nasim, S.A.; Dhir, B.; Samar, F.; Rashmi, K.; Mahmooduzzafar, M.A. Sulphur treatment alters the therapeutic potency of alliin obtained from garlic leaf extract. Food Chem. Toxicol. 2009, 47, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, F.M. Selenium deficiency and toxicity in the environment. In Essentials of Medical Geology; Selinus, O., Ed.; Springer: Dordrecht, The Netherlands, 2013; pp. 375–416. [Google Scholar]
- De Temmerman, L.; Waegeneers, N.; Thiry, C.; Du Laing, G.; Tack, F.; Ruttens, A. Selenium content of Belgian cultivated soils and its uptake by field crops and vegetables. Sci. Total Environ. 2014, 468–469, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Wei, C.; Tu, S. The roles of Selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Hurst, R.; Siyame, E.W.P.; Young, S.D.; Chilimba, A.D.C.; Joy, E.J.M.; Black, C.R.; Ander, E.L.; Watts, M.J.; Chilima, B.; Gondwe, J.; et al. Soil-type influences human Selenium status and underlies widespread Selenium deficiency risks in Malawi. Sci. Rep. 2013, 3, 1425. [Google Scholar] [CrossRef] [PubMed]
- Hanousek, O.; Mason, S.; Santner, J.; Chowdhury, M.M.A.; Berger, T.W.; Prohaska, T. Novel diffusive gradients in thin films technique to assess labile sulfate in soil. Anal. Bioanal. Chem. 2016, 408, 6759–6767. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.C.; Peverill, K.I.; Brennan, R.F. Soil potassium—Crop response calibration relationships and criteria for field crops grown in Australia. Crop Pasture Sci. 2013, 64, 523–530. [Google Scholar] [CrossRef]
- Piotrowska-Długosz, A.; Siwik-Ziomek, A.; Długosz, J.; Gozdowski, D. Spatio-temporal variability of soil Sulfur content and arylsulfatase activity at a conventionally managed arable field. Geoderma 2017, 295, 107–118. [Google Scholar] [CrossRef]
- Ahmed, H.P.; Schoenau, J.J.; King, T.; Kar, G. Effects of seed-placed sulfur fertilizers on canola, wheat, and pea yield; Sulfur uptake; and soil sulfate concentrations over time in three prairie soils. J. Plant Nutr. 2017, 40, 543–557. [Google Scholar] [CrossRef]
- Sarfaraz, Q.; Perveen, S.; Shahab, Q.; Muhammad, D.; Bashir, S.; Ahmed, N.; Ahmed, S.; Shahid-ul-islam, M.; Asghar, I. Comparative effect of soil and foliar application of sulfur on maize. J. Agric. Vet. Sci. 2014, 7, 32–37. [Google Scholar]
- López-Gutiérrez, M.L.; Benavides-Mendoza, A.; Ortega-Ortíz, H.; Valdez-Aguilar, L.A.; Cabrera-De la Fuente, M.; Sandoval-Rangel, A. Selenium and its effect on antioxidant status and mineral composition of lettuce. Rev. Mex. Ciencias Agricolas 2015, 12, 2257–2262. [Google Scholar]
- Põldma, P.; Moor, U.; Tõnutare, T.; Herodes, K.; Rebane, R. Selenium treatment under field conditions affects mineral nutrition, yield and antioxidant properties of bulb onion (Allium cepa L.). Acta Sci. Pol. Hortorum Cultus 2013, 12, 167–181. [Google Scholar]
- Sánchez-Rodas, D.; Mellano, F.; Martínez, F.; Palencia, P.; Giráldez, I.; Morales, E. Speciation analysis of Se-enriched strawberries (Fragaria ananassa Duch) cultivated on hydroponics by HPLC-TR-HG-AFS. Microchem. J. 2016, 127, 120–124. [Google Scholar] [CrossRef]
- De los Santos-Vázquez, M.E.; Benavides-Mendoza, A.; Ruiz-Torres, N.A.; Cabrera-De la Fuente, M.; Morelos-Moreno, A. Sodium selenite treatment of vegetable seeds and seedlings and the effect on antioxidant status. Emirates J. Food Agric. 2016, 28, 589–593. [Google Scholar]
- Chen, C.C.; Sung, J.M. Priming bitter gourd seeds with Selenium solution enhances germinability and antioxidative responses under sub-optimal temperature. Physiol. Plant. 2001, 111, 9–16. [Google Scholar] [CrossRef]
- Businelli, D.; D’Amato, R.; Onofri, A.; Tedeschini, E.; Tei, F. Se-enrichment of cucumber (Cucumis sativus L.), lettuce (Lactuca sativa L.) and tomato (Solanum lycopersicum L. Karst) through fortification in pre-transplanting. Sci. Hortic. 2015, 197, 697–704. [Google Scholar] [CrossRef]
- Orvis, K.S.; Goldman, I.L. Relationship between antiplatelet activity and Sulfur fertility in hydroponic and field-grown onions (Allium cepa). HortScience 1997, 32, 523–524. [Google Scholar]
- Zhang, L.; Hu, B.; Li, W.; Che, R.; Deng, K.; Li, H.; Yu, F.; Ling, H.; Li, Y.; Chu, C. OsPT2, a phosphate transporter, is involved in the active uptake of selenite in rice. New Phytol. 2014, 201, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-F.; McGrath, S.P.; Zhao, F.-J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Q.; Mitani, N.; Yamaji, N.; Shen, R.F.; Ma, J.F. Involvement of Silicon influx transporter OsNIP2;1 in selenite uptake in rice. Plant Physiol. 2010, 153, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Hopper, J.L.; Parker, D.R. Plant availability of selenite and selenate as influenced by the competing ions phosphate and sulfate. Plant Soil 1999, 210, 199–207. [Google Scholar] [CrossRef]
- Roca-Perez, L.; Gil, C.; Cervera, M.L.; Gonzálvez, A.; Ramos-Miras, J.; Pons, V.; Bech, J.; Boluda, R. Selenium and heavy metals content in some Mediterranean soils. J. Geochem. Explor. 2010, 107, 110–116. [Google Scholar] [CrossRef]
- Zayed, A.; Lytle, C.M.; Terry, N. Accumulation and volatilization of different chemical species of Selenium by plants. Planta 1998, 206, 284–292. [Google Scholar] [CrossRef]
- Zimmerman, M.T.; Bayse, C.A.; Ramoutar, R.R.; Brumaghim, J.L. Sulfur and selenium antioxidants: Challenging radical scavenging mechanisms and developing structure-activity relationships based on metal binding. J. Inorg. Biochem. 2015, 145, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Gill, R.; Kaushik, M.; Hasanuzzaman, M.; Pereira, E.; Ahmad, I.; Tuteja, N.; Gill, S.S. ATP-sulfurylase, Sulfur-compounds, and plant stress tolerance. Front. Plant Sci. 2015, 6, 210. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Kumar, A.; Prakash, R.; Prakash, N.T. Selenium accumulation and Se-induced anti-oxidant activity in Allium cepa. Environ. Inform. Arch. 2007, 5, 328–336. [Google Scholar]
- Bystrická, J.; Kavalcová, P.; Musilová, J.; Tomáš, J.; Tóth, T.; Orsák, M. Selenium and Its influence on the content of polyphenol compounds in onion (Allium cepa L.). J. Microbiol. Biotechnol. Food Sci. 2015, 4, 23–26. [Google Scholar] [CrossRef]
- Põldma, P.; Tõnutare, T.; Viitak, A.; Luik, A.; Moor, U. Effect of Selenium treatment on mineral nutrition, bulb size, and antioxidant properties of garlic (Allium sativum L.). J. Agric. Food Chem. 2011, 59, 5498–5503. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Lian, H.F.; Liu, Y.Y.; Yu, X.H.; Sun, Y.L.; Sun, X.D.; Shi, Q.H.; Liu, S.Q. Effects of Selenium and Sulfur on antioxidants and physiological parameters of garlic plants during senescence. J. Integr. Agric. 2016, 15, 566–572. [Google Scholar] [CrossRef]
- Zhao, J.; Hu, Y.; Gao, Y.; Li, Y.; Li, B.; Dong, Y.; Chai, Z. Mercury modulates Selenium activity via altering its accumulation and speciation in garlic (Allium sativum). Metallomics 2013, 5, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Gupta, S.; Prakash, R.; Spallholz, J.; Prakash, N.T. Selenium uptake by Allium cepa grown in Se-spiked soils. Am. J. Agric. Environ. Sci. 2007, 2, 80–84. [Google Scholar]
- Afton, S.E.; Caruso, J.A. The effect of Se antagonism on the metabolic fate of Hg in Allium fistulosum. J. Anal. At. Spectrom. 2009, 24, 759–766. [Google Scholar] [CrossRef]
- Zhao, J.; Gao, Y.; Li, Y.-F.; Hu, Y.; Peng, X.; Dong, Y.; Li, B.; Chen, C.; Chai, Z. Selenium inhibits the phytotoxicity of Mercury in garlic (Allium sativum). Environ. Res. 2013, 125, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Kopsell, D.A.; Randle, W.M. Selenate concentration affects Selenium and Sulfur uptake and accumulation by “Granex 33” onions. J. Am. Soc. Hortic. Sci. 1997, 122, 721–726. [Google Scholar]
- Kápolna, E.; Shah, M.; Caruso, J.A.; Fodor, P. Selenium speciation studies in Se-enriched chives (Allium schoenoprasum) by HPLC-ICP-MS. Food Chem. 2007, 101, 1398–1406. [Google Scholar] [CrossRef]
- Randle, W.M.; Block, E.; Littlejohn, M.H.; Putman, D.; Bussard, M.L. Onion (Allium cepa L.) thiosulfinates respond to increasing sulfur fertility. J. Agric. Food Chem. 1994, 42, 2085–2088. [Google Scholar] [CrossRef]
- Liu, S.; He, H.; Feng, G.; Chen, Q. Effect of Nitrogen and Sulfur interaction on growth and pungency of different pseudostem types of Chinese spring onion (Allium fistulosum L.). Sci. Hortic. 2009, 121, 12–18. [Google Scholar] [CrossRef]
- Mishu, H.M.; Ahmed, F.; Rafii, M.Y.; Golam, F.; Latif, M.A. Effect of Sulphur on growth, yield and yield attributes in onion (Allium cepa L.). Aust. J. Crop Sci. 2013, 7, 1416–1422. [Google Scholar]
- Nasreen, S.; Imamul Haq, S.; Hossain, A. Sulphur effects on growth responses and yield of onion. Asian J. Plant Sci. 2003, 2, 807–902. [Google Scholar]
- Randle, W.M. Onion germplasm interacts with Sulfur fertility for plant sulfur utilization and bulb pungency. Euphytica 1992, 59, 151–156. [Google Scholar] [CrossRef]
- Randle, W.M.; Bussard, M.L. Pungency and sugars of short-day onions as affected by Sulfur nutrition. J. Am. Soc. Hortic. Sci. 1993, 118, 766–770. [Google Scholar]
- McCallum, J.; Porter, N.; Searle, B.; Shaw, M.; Bettjeman, B.; McManus, M. Sulfur and Nitrogen fertility affects flavour of field-grown onions. Plant Soil 2005, 269, 151–158. [Google Scholar] [CrossRef]
- Bolandnazar, S.; Mollavali, M.; Tabatabaei, S.J. Influence of NH4NO3 and K2SO4 on qualitative characteristics of onion. Sci. Hortic. 2012, 136, 24–28. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, J.; Christie, P.; Li, X. Pungency of Spring Onion as affected by inoculation with arbuscular mycorrhizal fungi and Sulfur supply. J. Plant Nutr. 2007, 30, 1023–1034. [Google Scholar] [CrossRef]
- Hamilton, B.K.; Pike, L.M.; Yoo, K.S. Clonal variations of pungency, sugar content, and bulb weight of onions due to Sulphur nutrition. Sci. Hortic. 1997, 71, 131–136. [Google Scholar] [CrossRef]
- Coolong, T.W.; Kopsell, D.A.; Kopsell, D.E.; Randle, W.M. Nitrogen and Sulfur influence nutrient usage and accumulation in onion. J. Plant Nutr. 2004, 27, 1667–1686. [Google Scholar] [CrossRef]
- Youssif, B.D.; Hosna, M.A.F.; Mervat, A.A.T. Effect of Sulphur and Sulphur oxidizing bacteria on growth and production of garlic (Allium sativum, L.) under saline conditions. Middle East J. Agric. Res. 2015, 4, 446–459. [Google Scholar]
- Kápolna, E.; Fodor, P. Speciation analysis of Selenium enriched green onions (Allium fistulosum) by HPLC-ICP-MS. Microchem. J. 2006, 84, 56–62. [Google Scholar] [CrossRef]
- Prakash, N.T.; Sharma, N.; Prakash, R.; Nathaniel, T.N.; Acharya, R.; Reddy, A.V.R. Selenium fortification and pro/anti oxidant responses in Allium cepa (onion) cultivated in Se supplemented soils. Exp. Agric. 2010, 46, 531–540. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, J.; Christie, P.; Li, X. Influence of Nitrogen and Sulfur fertilizers and inoculation with arbuscular mycorrhizal fungi on yield and pungency of spring onion. J. Plant Nutr. 2006, 29, 1767–1778. [Google Scholar] [CrossRef]
- Hamilton, B.K.; Yoo, K.S.; Pike, L.M. Changes in pungency of onions by soil type, Sulphur nutrition and bulb maturity. Sci. Hortic. 1998, 74, 249–256. [Google Scholar] [CrossRef]
- Nasreen, S.; Haque, M.M.; Hossain, M.A.; Farid, A.T.M. Nutrient uptake and yield of onion as influenced by Nitrogen and Sulphur fertilization. Bangladesh J. Agric. Res. 2007, 32, 413–420. [Google Scholar] [CrossRef]
- De Souza, L.F.G.; Filho, A.B.C.; de Túlio, F.A.; Nowaki, R.H.D. Effect of Sulphur dose on the productivity and quality of onions. Aust. J. Crop Sci. 2015, 9, 728–733. [Google Scholar]
- Díaz-Pérez, J.C.; Bautista, J.; Bateman, A.; Gunawati, G.; Riner, C. Sweet onion (Allium cepa) plant growth and bulb yield and quality as affected by Potassium and Sulfur fertilization rates. HortScience 2016, 51, 1592–1595. [Google Scholar] [CrossRef]
- Lee, E.J.; Yoo, K.S.; Jifon, J.; Patil, B.S. Application of extra Sulfur to high-Sulfur soils does not increase pungency and related compounds in shortday onions. Sci. Hortic. 2009, 123, 178–183. [Google Scholar] [CrossRef]
- Randle, W.M.; Bussard, M.L.; Warnock, D.F. Ontogeny and Sulfur fertility affect leaf Sulfur in short-day onions. J. Am. Soc. Hortic. Sci. 1993, 118, 762–765. [Google Scholar]
- Randle, W.M.; Lancaster, J.E.; Shaw, M.L.; Sutton, K.H.; Hay, R.L.; Bussard, M.L. Quantifying onion flavor compounds responding to Sulfur fertility-Sulfur increases levels of Alk(en)yl Cysteine Sulfoxides and biosynthetic intermediates. J. Am. Soc. Hortic. Sci. 1995, 120, 1075–1081. [Google Scholar]

{kind=link}
{kind=link}
| Biofortification | Phytochemical | References |
|---|---|---|
| S in A. roseum | Diallyl disulfide | [136] |
| S in A. roseum | Diallyl thiosulfinate (Allicin) | [136] |
| S in A. roseum | Methyl allyl disulfide | [136] |
| Se in A. tricoccum | Se-methylselenocysteine | [137] |
| Se in garlic and A. ascalunicum | γ-glutamyl-Se-methlyselenocysteine (γ-GluMeSeCys) | [138] |
| Se in garlic and chives | γ-glutamyl-Se-methlyselenocysteine (γ-GluMeSeCys) | [139,140] |
| Se in garlic and chives | Se-methylselenocysteine | [140,141] |
| S in onion | γ-glutamyl-1-propenyl cysteine sulfoxide (γGPECSO) | [142] |
| S in onion | Propyl cysteine sulfoxide (Propiin) | [143] |
| S in onion | S-methyl alkyl cysteine sulfoxides (Mettin) | [143] |
| S in onion | S-methyl-l-cysteine sulfoxide | [144] |
| S in onion | Trans-S-1-propenyl-l-cysteine sulfoxide (1-PRENCSO) | [142] |
| Se in onion | S-methyl-l-cysteine sulfoxide | [50] |
| S in onion and garlic | Propenyl cysteine sulfoxide (Isoalliin) | [145,146] |
| S in onion and garlic | S-allyl cysteine sulfoxide (Aliin) | [145,147] |
| Species | Chemical Form | Application Form | Quantity Supplied | Results | Reference |
|---|---|---|---|---|---|
| Chives | Na2SeO3 | Nutrient solution | 30 mg·L−1 | Accumulation of Se in root > leaf. Antagonism with Hg. | [179] |
| Chives | Na2SeO3 | Soil | 1, 2, 3, 5 and 15 mg·kg−1 Se | Increases Se content. | [140] |
| Chives | Se(IV), SeMet | Nutrient solution | 10 and 100 mg·L−1 | Higher stress tolerance and Se accumulation with SeMet. | [195] |
| Elephant garlic | Na2SeO3, Na2SeO4 | Soil | 0.2, 1.3, 2.6 y 3.8 mg·kg−1 Se | Accumulation of selenate > selenite. | [65] |
| Garlic | K2SeO3 y K2SeO4 | Hydroponics | 50 µmol·L−1 | Increases Se content. Antagonism with S. | [141] |
| Garlic | Na2SeO3 | Hydroponics | 3 and 6 µmol·L−1 | Low dose increases biomass and delays senescence. | [176] |
| Garlic | Na2SeO4 | Leaf spray | 10, 50 and 100 mg·L−1 | Antagonism with S, K and Ca. Increases antioxidant capacity. | [175] |
| Garlic | Na2SeO3, Na2SeO4 | Nutrient solution | 0.01, 0.1, 1, 10, 100 mg·L−1 | Inhibition of growth in high doses. Decreases Hg toxicity. | [180] |
| Onion | Na2SeO3 | Soil and foliar spray | 10, 20 and 40 kg·ha−1 on soil. 0.5, 1, 1.5, 2, 2.5 and 3 mg·L−1 leaf spray | Antagonism with S. | [31] |
| Onion | Na2SeO4 | Foliar spray | 10, 50 and 100 mg·L−1 | Antagonism with S. 50 μg·mL−1 increases Se content. | [159] |
| Onion | Na2SeO4 | Hydroponics | 1, 2, 4 and 8 mg·L−1 SeO4 | Increases Se content. Antagonism with S. | [18] |
| Onion | Na2SeO4 | Hydroponics | 2.0 mg L−1 | Increases Se content. | [50] |
| Onion | Na2SeO4 | Hydroponics | 0.5, 1.0, 1.5 and 2.0 mg·L−1 | High concentrations decrease growth. Low concentration increases S content. | [181] |
| Onion | Na2SeO3, Na2SeO4 | Nutrient solution | 5 mg·L−1 | Affects growth. Bulb accumulation > leaf > root. | [173] |
| Onion | Na2SeO3 and Na2SeO4 | Soil | 2.5, 5.0 and 7.5 mg·kg−1 | Increase the content of Se. Decreases growth. Selenite is more beneficial than selenate. | [196] |
| Onion | Se(VI) | Soil | 25 and 50 mg·kg−1 | Accumulation of Se, high dose decreases bulb size. | [178] |
| Scallion | Selenite, selenate, SeMet | Nutrient solution | 10 mg·L−1 | Higher concentration of Se with selenate. 30% of the Se is inorganic. | [182] |
| Species | Chemical Form | Application Form | Quantity Supplied | Results | Reference |
|---|---|---|---|---|---|
| Chives | K2SO4 | Soil | 60 mg·kg−1 soil | Increased S content. | [197] |
| Chives | K2SO4, MgSO4 and H2SO4 | Nutrient solution | 0.01 and 4.0 mmol·L−1 SO4 | Increased biomass, N, S, and pungency. | [184] |
| Chives | MgSO4 and K2SO4 | Soil | 0.1, 1.75 and 4.0 mM SO4 | Increased content of pyruvate, S, and dry weight. | [191] |
| Onion | CaSO4 | Hydroponics | 0.8, 4.8, 10.8 and 14.8 mol·m−3 | Increased concentration of S. Antagonism with Se. | [18] |
| Onion | CaSO4 | Soil | 22.4 kg·ha−1 S | No effects. | [198] |
| Onion | CaSO4 | Soil | 200 kg·ha−1 S | Increased S, pungency, and pyruvic acid concentration. | [189] |
| Onion | CaSO4 | Soil | 20, 40 and 60 kg·ha−1 S | Up to 40 kg ha−1 increased growth and yield. | [199] |
| Onion | Elemental sulfur (Sulfurgran®) | Soil | 15, 30, 45, 60 and 90 kg·ha−1 | Improves growth and yield. | [200] |
| Onion | H2SO4 | Nutrient solution | 0.1 and 4.0 meq·L−1 | Increased pungency and concentration of S. | [187] |
| Onion | H2SO4 | Nutrient solution | 0.1 and 4.0 meq·L−1 | Increased pungency and concentration of S. | [188] |
| Onion | K2Mg2(SO4)3 | Soil | 30 and 50 kg·ha−1 S | Increased S and pyruvate content. | [143] |
| Onion | K2O3S2 | Soil | 80, 126, 172, 218 and 264 kg·ha−1 S | No effect on growth and yield. | [201] |
| Onion | Liquid sulfur (17%) | Soil surface | 13 and 26 kg·ha−1 | No effect. | [202] |
| Onion | MgSO4 | Nutrient solution | 0.1 and 4.0 meq·L−1 | Increased pyruvic acid and S concentration. | [203] |
| Onion | MgSO4 | Nutrient solution | 0.1, 0.48, 0.85, 1.6, and 3.1 meq·L−1 | Increased bulb fresh weight up to 1.6 meq L−1. | [204] |
| Onion | MgSO4 | Nutrient solution | 5, 25, 50, 75 and 150 mg·L−1 | Increased total S and sulfates. Differences in varieties in pyruvic acid. | [29] |
| Onion | MgSO4 | Nutrient solution | 5, 45 and 125 mg·L−1 | S application increased bulb weight. | [142] |
| Onion | MgSO4 | Nutrient solution | 1.7, 15 and 41.7 mg·L−1 | Positive effect in pungency. Antagonism with B, Fe, Mn, and Zn. | [193] |
| Onion | MgSO4 and CaSO4 | Nutrient solution | 2 and 123 mg·L−1 | S in leaves, pungency, and yield decreases with low S level. | [192] |
| Onion | Na2SO4 | Soil | 15, 30 and 45 kg·ha−1 S | Increased pyruvic acid. Antagonism with Se. | [31] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Morales, S.; Pérez-Labrada, F.; García-Enciso, E.L.; Leija-Martínez, P.; Medrano-Macías, J.; Dávila-Rangel, I.E.; Juárez-Maldonado, A.; Rivas-Martínez, E.N.; Benavides-Mendoza, A. Selenium and Sulfur to Produce Allium Functional Crops. Molecules 2017, 22, 558. https://doi.org/10.3390/molecules22040558
González-Morales S, Pérez-Labrada F, García-Enciso EL, Leija-Martínez P, Medrano-Macías J, Dávila-Rangel IE, Juárez-Maldonado A, Rivas-Martínez EN, Benavides-Mendoza A. Selenium and Sulfur to Produce Allium Functional Crops. Molecules. 2017; 22(4):558. https://doi.org/10.3390/molecules22040558
Chicago/Turabian StyleGonzález-Morales, Susana, Fabián Pérez-Labrada, Ema Laura García-Enciso, Paola Leija-Martínez, Julia Medrano-Macías, Irma Esther Dávila-Rangel, Antonio Juárez-Maldonado, Erika Nohemí Rivas-Martínez, and Adalberto Benavides-Mendoza. 2017. "Selenium and Sulfur to Produce Allium Functional Crops" Molecules 22, no. 4: 558. https://doi.org/10.3390/molecules22040558
APA StyleGonzález-Morales, S., Pérez-Labrada, F., García-Enciso, E. L., Leija-Martínez, P., Medrano-Macías, J., Dávila-Rangel, I. E., Juárez-Maldonado, A., Rivas-Martínez, E. N., & Benavides-Mendoza, A. (2017). Selenium and Sulfur to Produce Allium Functional Crops. Molecules, 22(4), 558. https://doi.org/10.3390/molecules22040558