Cadmium Disrupts Subcellular Organelles, Including Chloroplasts, Resulting in Melatonin Induction in Plants



Department of Biotechnology, Bioenergy Research Center, College of Agriculture and Life Sciences, Chonnam National University, 77 Yongbong-ro, Buk-gu, Gwangju 61186, Korea
*
Author to whom correspondence should be addressed.
Molecules 2017, 22(10), 1791; https://doi.org/10.3390/molecules22101791
Submission received: 13 September 2017
/
Revised: 18 October 2017
/
Accepted: 21 October 2017
/
Published: 23 October 2017
(This article belongs to the Special Issue Melatonin as an Antioxidant and a Functionally Pleiotropic Molecule: Synthesis, Metabolism and Activities in Organisms)
{kind=link}
{kind=link}
Abstract
:Cadmium is a well-known elicitor of melatonin synthesis in plants, including rice. However, the mechanisms by which cadmium induces melatonin induction remain elusive. To investigate whether cadmium influences physical integrities in subcellular organelles, we treated tobacco leaves with either CdCl2 or AlCl3 and monitored the structures of subcellular organelles—such as chloroplasts, mitochondria, and the endoplasmic reticulum (ER)—using confocal microscopic analysis. Unlike AlCl3 treatment, CdCl2 (0.5 mM) treatment significantly disrupted chloroplasts, mitochondria, and ER. In theory, the disruption of chloroplasts enabled chloroplast-expressed serotonin N-acetyltransferase (SNAT) to encounter serotonin in the cytoplasm, leading to the synthesis of N-acetylserotonin followed by melatonin synthesis. In fact, the disruption of chloroplasts by cadmium, not by aluminum, gave rise to a huge induction of melatonin in rice leaves, which suggests that cadmium-treated chloroplast disruption plays an important role in inducing melatonin in plants by removing physical barriers, such as chloroplast double membranes, allowing SNAT to gain access to the serotonin substrate enriched in the cytoplasm.
1. Introduction
Melatonin (N-acetyl-5-methoxytryptamine) is a well-known bioactive molecule found in all living organisms tested so far. It functions as a neurohormone in animals [1,2,3] and as a biostimulator in plants [4,5,6]. Its universal distribution is associated with its endosymbiotic origins from the alpha-protobacteria and cyanobacteria that evolved into mitochondria and chloroplasts in eukaryotes [7]. In particular, plants respond to various environmental stresses—including drought [8,9], salt [10,11], cold [12,13,14], heavy metals [15,16,17,18], high temperature [19,20], and pathogens [21,22,23,24]—by inducing melatonin synthesis to confer resistance against these stimuli. During the past decades, many abiotic stresses—such as light [25,26,27], hydrogen peroxide [28], and cold [29]—were applied to plants to induce melatonin biosynthesis, among which cadmium was the most efficient elicitor of melatonin induction. For example, in the detached leaves of rice, the highest and second highest levels of melatonin were induced by cadmium treatment and peroxydizing herbicide butafenacil, respectively, whereas lead failed to induce melatonin [30]. Cadmium also triggered increased melatonin in other plants—including tomato, alfalfa, and Arabidopsis [16,17,31]—which suggests beneficial effects of melatonin against cadmium stress. This hypothesis has been proven, as many melatonin-rich plants exhibit resistance against cadmium stress [6,16].
Melatonin synthesis requires four enzymatic steps: first, tryptophan is decarboxylated into tryptamine by tryptophan decarboxylase (TDC), and then serotonin is synthesized via tryptamine 5-hydroxylase (T5H). Next is catalysis by serotonin N-acetyltransferase (SNAT). Finally, N-acetylserotonin O-methyltransferase (ASMT) catalyzes the final reaction [32]. Cadmium and nitric oxide (NO) treatments equally induced all melatonin biosynthetic gene transcripts except SNAT, but an increase in melatonin occurred only as a result of cadmium treatment [33]. This result indicates that the ample production of serotonin by NO treatment was not coupled with melatonin synthesis because of possible inaccessibility of serotonin into chloroplasts in which the enzyme SNAT is constitutively expressed. Thus, we hypothesized that cadmium treatment may disrupt chloroplast structures that trigger SNAT to gain access to and easily encounter serotonin for melatonin production.
To prove this hypothesis, we treated tobacco leaves with both cadmium and aluminum and examined them using confocal microscopic analysis to see whether the structures of subcellular organelles—such as chloroplasts, mitochondria, and ER—were affected. We found that cadmium disrupted chloroplasts, mitochondria, and ER structures, whereas aluminum (Al) only disrupted ER slightly. In addition, Al treatment failed to induce melatonin in rice seedlings, whereas Cd did induce melatonin. The reason for using rice plant species to measure melatonin was its easy and ample induction of melatonin upon Cd compared to tobacco. In fact, tobacco leaves marginally induced melatonin in response to Cd as did tomato [16]. These results suggest that chloroplast disruption due to cadmium plays an important role in inducing melatonin synthesis in plants.
2. Results and Discussion
2.1. Cadmium Treatment Disrupts the Structures of Chloroplasts, Mitochondria, and ER
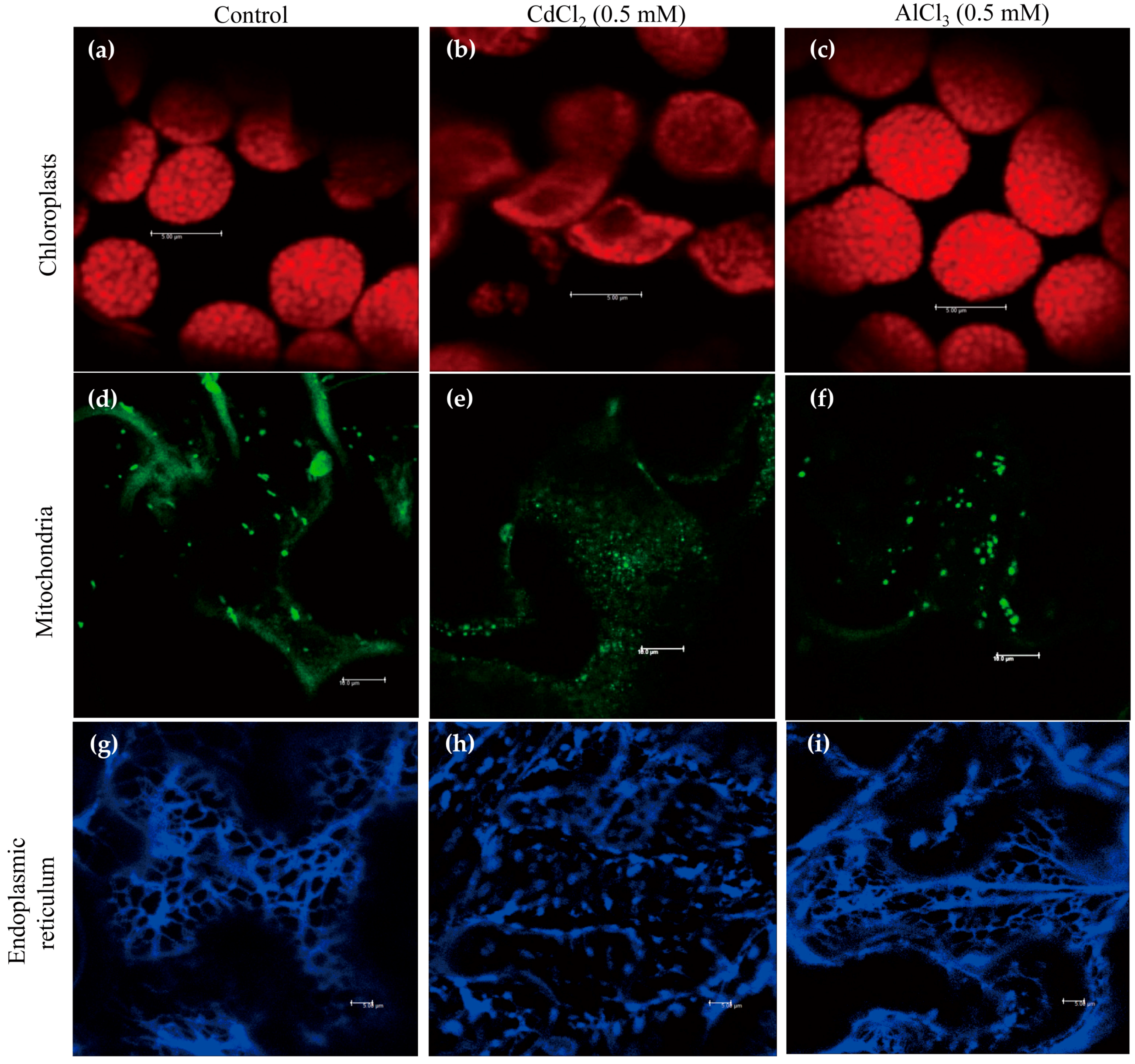
We used 0.5 mM cadmium to investigate whether cadmium treatment affects the structures of various subcellular organelles, such as chloroplasts, mitochondria, and ER, as this concentration induced the highest level of melatonin in rice [33]. Aluminum was used for the control treatment because it increased melatonin in soybean [15]. Visualization of these organelle structures was detected by chlorophyll fluorescence, MitoTracker Green FM staining, and HDEL-CFP (cyan fluorescent protein) marker protein.
In response to cadmium treatment, chloroplasts were swollen, distorted, and worn down, whereas control and aluminum treatments maintained the integrity of the chloroplasts (Figure 1a–c). Similarly, cadmium treatment abolished the structures of the mitochondria, but no such damage was observed in mitochondria treated with aluminum, which suggests the susceptibility of mitochondria to cadmium (Figure 1d–f). By contrast, cadmium and aluminum treatments ruined a tubular ER network, whereas the control showed a normal ER network (Figure 1g–i). Of the three subcellular organelles, ER seemed to be the most sensitive to heavy metals because ER structures were labile to aluminum treatment.
These results clearly show that cadmium treatment severely disrupts the integrity of chloroplasts, mitochondria, and ER. Such disruption could result in the leaking of endogenous SNAT enzymes [9,32] out of these organelles’ double membranes, where they could readily encounter cytoplasmic serotonin for N-acetylserotonin synthesis followed by the production of melatonin via the catalytic reaction of cytoplasmic ASMT enzyme in response to cadmium stress. Unlike cadmium, aluminum treatment had no effect on the wreckage of chloroplasts and mitochondria except ER, indicative of an ineffective elicitor for melatonin induction. ER also plays an important role in serotonin synthesis rather than melatonin synthesis because T5H is a P450 enzyme localized in ER responsible for serotonin synthesis. The effects of ER disruption in view of melatonin induction by cadmium remain to be examined, but it seems unlikely that ER damage is the major cause of melatonin induction by cadmium, because aluminum also destroyed the ER structure.
2.2. Melatonin Induction in Rice Seedlings Following Cadmium and Aluminum Treatments
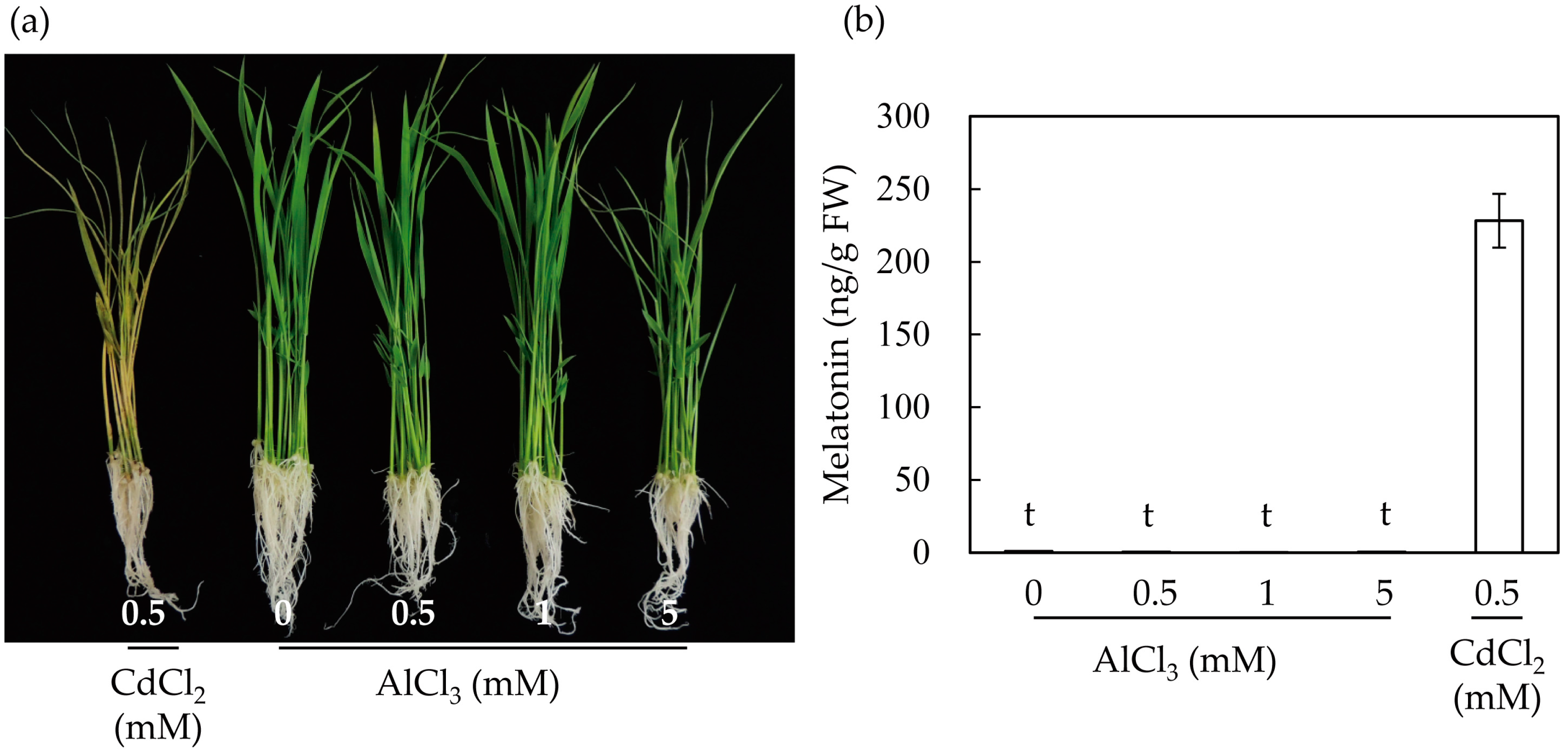
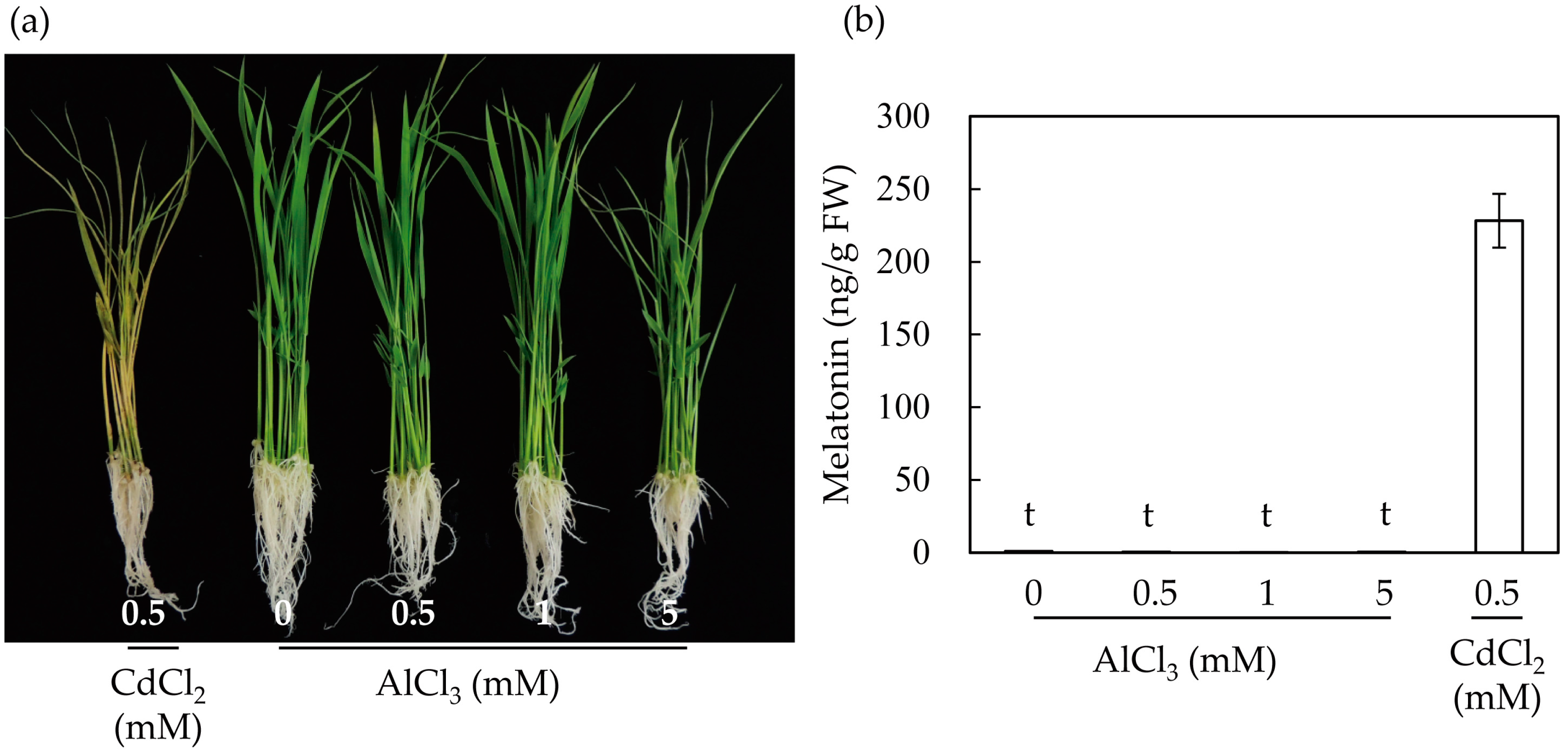
To further determine the effects of aluminum on the induction of melatonin in rice plants, we treated seven-day-old rice seedlings with varying levels of aluminum for three days. As a control, rice seedlings were treated with 0.5 mM cadmium for three days. The rice seedlings treated with cadmium showed phenotypes with severely necrotizing leaves (Figure 2a). By contrast, rice seedlings treated with 0.5–1 mM aluminum exhibited relatively healthy phenotypes, but rice seedlings treated with 5 mM aluminum showed necrosis in some parts of the leaves. Next, we determined the melatonin levels in the rice seedlings treated with cadmium and aluminum. As shown in Figure 2b, melatonin induction was not observed in the rice seedlings treated with aluminum, even at a concentration of 5 mM aluminum, whereas treatment with cadmium (0.5 mM) produced 228 ng/g fresh weight (FW). Together, these results suggest that in light of the subcellular organelles and SNAT enzyme catalysis, one of the main reasons for melatonin induction was the disruption of chloroplasts upon treatment with cadmium.
3. Materials and Method
3.1. Confocal Microscopic Analysis
Tobacco (Nicotiana benthamiana) plants were grown at 25 °C under a 16 h light/8 h dark period in an environmentally controlled growth room. The pBIN61-HDEL-CFP plasmid as the ER fluorescence marker was kindly donated by Dr. HG Kang (Texas State University, San Marcos, TX, USA). This plasmid was transformed to the Agrobacterium tumefaciens strain GV2260 using the freeze–thaw method, and transient expression analyses were performed as described by Voinnet et al. [34].
Two-week-old N. benthamiana leaves were syringe-infiltrated with GV2260 carrying the pBIN61-HDEL-CFP plasmid, then incubated in the growth room for two days. These tobacco leaves were further treated with cadmium (0.5 mM) or aluminum (0.5 mM) for 4 h, and images were analyzed using a Leica TCS-SP5 confocal microscope (×63 oil immersion objective) (Leica, Wetzlar, Germany). CFP was excited with the Mercury arc lamp (435 nm) and emitted light was collected between 480 and 520 nm. Images were processed using Leica LAS-AF Version 1.8.2 (Leica, Wetzlar, Germany).
MitoTracker Green FM probes, 50 µg (Invitrogen, Carlsbad, CA, USA), were sequentially dissolved with DMSO to obtain 50 mM stock solution and diluted with 3 mM MES solution (pH5.6) to obtain a working concentration of 1 µM. Following treatment with cadmium (0.5 mM) or aluminum (0.5 mM) for 4 h, tobacco leaves were stained with the MitoTracker probe for 15 min at room temperature and rinsed with fresh water before confocal analysis. The probe was excited with a blue argon ion laser (488 nm), and emitted light was collected between 494 and 546 nm.
The chloroplast autofluorescence was excited with a blue argon laser (488 nm), and emitted light was collected from 660 to 731 nm.
3.2. Rice Seedlings
Rice seeds (Oryza sativa cv. Dongjin) were surface-sterilized and germinated on half-strength Murashige and Skoog (MS) medium (MB Cell, Seoul, Korea) in vertically oriented polystyrene square dishes (SPL Life Science, Pocheon-si, Korea). The environmental conditions for rice seedlings’ growth were 28 °C under a 12 h light/12 h dark cycle at a photosynthetic photon flux density of 150 µmol m−2 s−1. Seven days after seeding, the seedlings were harvested for cadmium and aluminum treatments. The rice variety cv. Dongin (japonica type) was not Al tolerant and Cd sensitive. It was reported that all japonica types of rice were equally sensitive to Al stress [35].
3.3. Cadmium and Aluminum Treatments and Melatonin Quantification in Rice Seedlings
Seven-day-old seedlings were transferred to 50 mL polypropylene conical tubes containing 25 mL water with 0.5 mM cadmium (CdCl2) or varying concentrations of aluminum (AlCl3). These seedlings were incubated for three days under continuous light as described previously [32]. The rice samples (shoot parts) were frozen in liquid nitrogen and pulverized to a powder using a TissueLyser II instrument (Qiagen, Tokyo, Japan). The powder (100 mg) was extracted with 1.5 mL chloroform for 1 h at room temperature. The chloroform extracts were evaporated until dry and dissolved in 0.1 mL 42% methanol. Aliquots of 10 µL were subjected to HPLC with a fluorescence detector system (Waters, Milford, MA, USA), as described previously [32].
4. Conclusions
In summary, cadmium treatment induced melatonin synthesis in rice plants, whereas treatment with aluminum did not induce melatonin synthesis. In terms of the structures of subcellular organelles, cadmium treatment led to the wreckage of chloroplasts and mitochondria, whereas aluminum treatment did not break down either chloroplasts or mitochondria. The resulting disruption of chloroplasts by cadmium facilitates the encounter of substrate serotonin with chloroplastidic SNAT enzymes, which in turn results in a huge induction of melatonin in rice plants. Our results confirm that one of the main reasons for the failure of melatonin induction despite the huge increase in serotonin upon NO treatment [33] or senescence [5] is interference with the entry of serotonin into chloroplasts in which SNAT enzymes reside. On the other hand, the cadmium-induced melatonin synthesis via the disruption of chloroplasts in cytoplasm should be interpreted with caution because the CoA is also very important for melatonin synthesis and there is high level of CoA in chloroplast relative over in the cytoplasm. Additionally, it is worth noting that the physicochemical properties of CdCl2 and AlCl3 may play different roles in melatonin synthesis as well. For example, AlCl3 is a potent Lewis acid, whilst CdCl2 is a mild Lewis acid. Moreover, AlCl3·H2O liberates HCl whereas CdCl2 forms complex ions, such as [CdCl4]2−. Further studies in this regard are therefore needed in the near future in view of melatonin induction in plants.
Acknowledgments
This research was supported by grants (2010-0020141 and NRF-2017R1A2A2A05069235) from the Basic Science Research Program of the National Research Foundation of Korea, funded by the Ministry of Education, Republic of Korea.
Author Contributions
H.-Y.L. performed the experiments. K.B. analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hardeland, R. Melatonin and the pathologies of weakened or dysregulated circadian oscillators. J. Pineal Res. 2017, 62, e12377. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.V.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, A.V.; Mosser, E.A.; Oikonomou, G.; Prober, D.A. Melatonin is required for the circadian regulation of sleep. Neuron 2015, 85, 1193–1199. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: Plant growth regulator and/or biostimulator during stress? Trend Plant Sci. 2014, 19, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Choi, G.H.; Lee, H.Y.; Back, K. Chloroplast overexpression of rice caffeic acid O-methyltransferase increases melatonin production in chloroplasts via the 5-methoxytryptamine pathway in transgenic rice plants. J. Pineal Res. 2017, 63, e12412. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Back, K. Overexression of rice serotonin N-acetyltransferase 1 in transgenic rice plants confers resistance to cadmium and senescence and increases grain yiled. J. Pineal Res. 2017, 62, e12392. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and chloroplasts as the original sites of melatonin synthesis: A hypothesis related to melatonin’s primary function and evolution in eukaryotes. J. Pineal Res. 2013, 54, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Fleta-Soriano, E.; Díaz, L.; Bonet, E.; Munné-Bosch, S. Melatonin may exert a protective role against drought stress in maize. J.Agron.Crop Sci. 2017, 203, 286–294. [Google Scholar] [CrossRef]
- Wang, L.; Feng, C.; Zheng, X.; Guo, Y.; Zhou, F.; Shan, D.; Liu, X.; Kong, J. Plant mitochondria synthezise melatonin and enhance the tolerance of plants to drought stress. J. Pineal Res. 2017, 63, e12429. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xie, Y.; Gu, Q.; Zhao, G.; Zhang, Y.; Cui, W.; Xu, S.; Wang, R.; Shen, W. The AtrbohF-dependent regulation of ROS signaling is required for melatonin-induced salinity tolerance in Arabidopsis. Free Radic. Biol. Med. 2017, 108, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Tan, D.X.; Allan, A.C.; Zuo, B.; Zhao, Y.; Reiter, R.J.; Wang, L.; Wang, Z.; Guo, Y.; Zhou, J.; et al. Chloroplastidic biosynthesis of melatonin and its involvement in protection of plants from salt stress. Sci. Rep. 2017, 7, 41236. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Back, K. 2-Hydroxymelatonin promotes the resistance of rice plant to multiple simultaneous abiotic stresses (combined cold and drought). J. Pineal Res. 2016, 61, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chang, J.; Zheng, J.; Dong, Y.; Liu, Q.; Yang, X.; Wei, C.; Zhang, Y.; Ma, J.; Zhang, X. Local melatonin application induces cold tolerance in distant organs of Citrullus lanatus L. via long distance transport. Sci. Rep. 2017, 7, 40858. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Wang, M.; Liu, B.; Zhang, S. Exogenous melatonin mitigates photoinhibition by accelerating non-photochemical quenching in tomato seedlings exposed to moderate light during chilling. Front. Plant Sci. 2017, 8, 244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zeng, B.; Mao, Y.; Kong, X.; Wang, X.; Yang, Y.; Zhang, J.; Xu, J.; Rengel, Z.; Chen, Q. Melatonin alleviates aluminium toxicity through modulating antioxidative enzymes and enhancing organic acid anion edudation in soybean. Funct. Plant Biol. 2017, 44, 961–968. [Google Scholar] [CrossRef]
- Cai, S.Y.; Zhang, Y.; Xu, Y.P.; Qi, Z.Y.; Li, M.Q.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Reiter, R.J.; et al. HsfA1 upregulates melatonin biosynthesis to confer cadmium tolerance in tomato plants. J. Pineal Res. 2017, 62, e12387. [Google Scholar] [CrossRef] [PubMed]
- Gu, Q.; Chen, Z.; Yu, X.; Cui, W.; Pan, J.; Zhao, G.; Xu, S.; Wang, R.; Shen, W. Melatonin confers plant tolerance against cadmium stress via the decrease of cadmium accumulation and reestablishment of microRNA-mediated redox homeostasis. Plant Sci. 2017, 261, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Q.; Hasan, K.; Li, C.X.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Reiter, R.J.; Yu, J.Q.; Xu, M.X.; et al. Melatonin mediates selenium-induced tolerance to cadmium stress in tomato plants. J. Pineal Res. 2016, 61, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, Y.; Zhang, X.; Du, H.; Xu, B.; Huang, B. Melatonin suppression of heat-induced leaf senescence involves changes in absicsic acid and cytokinin biosynthesis and signaling pathways in perennial rhygrass (Lolium perenne L.). Environ. Exp. Bot. 2017, 138, 36–45. [Google Scholar] [CrossRef]
- Xu, W.; Cai, S.Y.; Zhang, Y.; Wang, Y.; Ahammed, G.J.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Yu, J.Q.; Reiter, R.J.; et al. Melatonin enhances thermotolerance by promoting cellular protein protection in tomato plants. J. Pineal Res. 2016, 61, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Back, K. Mitogen-activated protein kinase pathways are required for melatonin-mediated defense responses in plants. J. Pineal Res. 2016, 60, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wei, Y.; He, C. Melatonin-induced CBF/DREB1s are essential for diurnal change of disease resistance and CCA1 expression in Arabidopsis. Plant Physiol. Biochem. 2016, 100, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Hu, W.; Wang, Q.; Zeng, H.; Li, X.; Yan, Y.; Reiter, R.J.; He, C.; Shi, H. Identification, transcriptional and functional analysis of heat-shock protein 90s in banana (Musa acuminata L.) highlight their novel role in melatonin-mediated plant response to Fusarium wilt. J. Pineal Res. 2017, 62, e12367. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Back, K. Melatonin is required for H2O2-and NO-mediated defense signaling through MAPKKK3 and OXI1 in Arabidopsis thaliana. J. Pineal Res. 2017, 62, e12379. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; KrishnaRaj, S.; Saxena, P.K. Tryptophan is a precursor for melatonin and serotonin biosynthesis in in vitro regenerated St. John’s wort (Hypericum perforatum L. cv. Anthos) plants. Plant Cell Rep. 2000, 19, 698–704. [Google Scholar] [CrossRef]
- Badria, F.A. Melatonin, serotonin, and tryptamine in some Egyptian food and medicinal plants. J. Med. Food 2002, 5, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Afreen, F.; Zobayed, S.M.A.; Kozai, T. Melatonin in Glycyrrhiza uralensis: Response of plant roots to spectral quality of light and UV-B radiation. J. Pineal Res. 2006, 41, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Chemical stress by different agents affects the melatonin content of barley roots. J. Pineal Res. 2009, 46, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Alan, A.R.; Cao, J.; Saxena, P.K. Melatonin and serotonin in flowers and fruits of Datura metel L. J. Pineal Res. 2009, 47, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Lee, H.Y.; Hwang, O.J.; Lee, H.J.; Lee, K.; Back, K. Coordinated regulation of melatonin synthesis and degradation genes in rice leaves in response to cadmium treatment. J. Pineal Res. 2015, 58, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Lee, H.J.; Lee, H.Y.; Back, K. Cloning and functional characterization of the Arabidopsis N-acetylserotonin O-methyltransferase responsible for melatonin synthesis. J. Pineal Res. 2016, 60, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Back, K.; Tan, D.X.; Reiter, R.J. Melatonin biosynthesis in plants: Multiple pathways catalyze tryptophan to melatonin in the cytoplasm or chloroplasts. J. Pineal Res. 2016, 61, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Choi, G.H.; Back, K. Cadmium-induced melatonin synthesis in rice requires light, hydrogen peroxide, and nitric oxide: Key regualtory roles for tryptophan decarboxylase and caffeic acid O-methyltransferase. J. Pineal Res. 2017, 63, e12441. [Google Scholar] [CrossRef] [PubMed]
- Voinnet, O.; Rivas, S.; Mestre, P.; Baulcombe, D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003, 33, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.J.; Seo, Y.J.; Futakuchi, K.; Vijarnsorn, P.; Ishii, R. Effects of aluminium toxicity on flowering time and grain yield on rice genotypes differing in Al-tolerance. J. Crop Sci. Biotechnol. 2011, 14, 305–309. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |
Figure 1.
Effects of cadmium and aluminum on the structures of subcellular organelles as shown by confocal microscopy. (a–c) Structures of chloroplasts: (a) Control; (b) Cadmium treatment; (c) Aluminum treatment; (d–f) Structures of mitochondria: (d) Control; (e) Cadmium treatment; (f) Aluminum treatment; (g–i) Structures of ER: (g) Control; (h) Cadmium treatment; (i) Aluminum treatment. Tobacco leaves were infiltrated with heavy metals (0.5 mM) for 4 h and subjected to confocal microscopic analysis. Bars = 10 µm (chloroplasts and mitochondria), 5 µm (ER).
Figure 1.
Effects of cadmium and aluminum on the structures of subcellular organelles as shown by confocal microscopy. (a–c) Structures of chloroplasts: (a) Control; (b) Cadmium treatment; (c) Aluminum treatment; (d–f) Structures of mitochondria: (d) Control; (e) Cadmium treatment; (f) Aluminum treatment; (g–i) Structures of ER: (g) Control; (h) Cadmium treatment; (i) Aluminum treatment. Tobacco leaves were infiltrated with heavy metals (0.5 mM) for 4 h and subjected to confocal microscopic analysis. Bars = 10 µm (chloroplasts and mitochondria), 5 µm (ER).
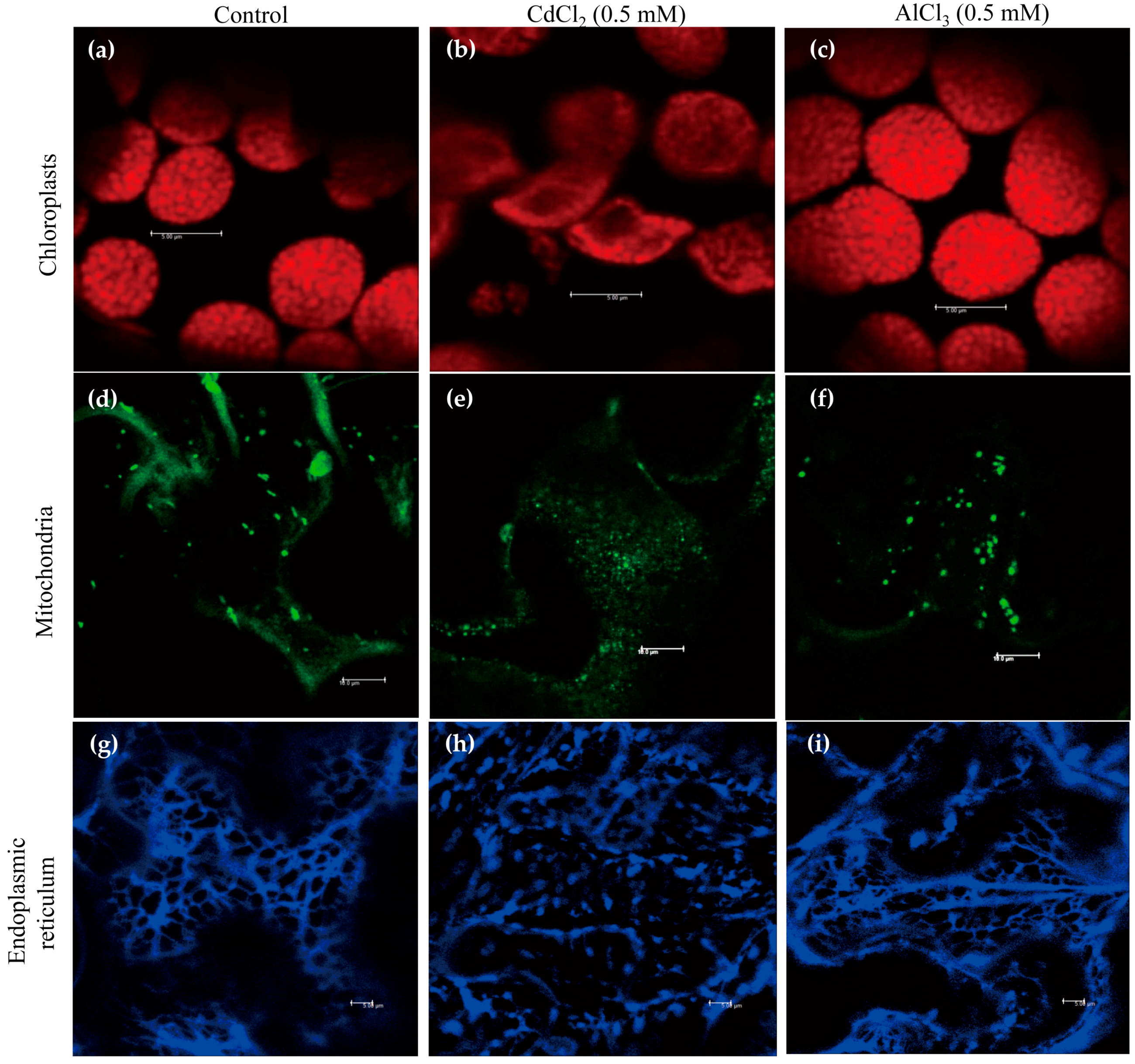
Figure 2.
Induction of melatonin in response to cadmium and aluminum treatments in seven-day-old rice seedlings. (a) Phenotypes of rice seedlings upon cadmium and aluminum treatments; (b) Quantification of melatonin by HPLC. Data represent mean ± standard error of the mean, n = 3. Rice seedlings grown for seven days were challenged with cadmium (0.5 mM) or aluminum of varying concentrations for three days under continuous light. FW, fresh weight; t denotes 1 ng/g FW.
Figure 2.
Induction of melatonin in response to cadmium and aluminum treatments in seven-day-old rice seedlings. (a) Phenotypes of rice seedlings upon cadmium and aluminum treatments; (b) Quantification of melatonin by HPLC. Data represent mean ± standard error of the mean, n = 3. Rice seedlings grown for seven days were challenged with cadmium (0.5 mM) or aluminum of varying concentrations for three days under continuous light. FW, fresh weight; t denotes 1 ng/g FW.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, H.-Y.; Back, K. Cadmium Disrupts Subcellular Organelles, Including Chloroplasts, Resulting in Melatonin Induction in Plants. Molecules 2017, 22, 1791. https://doi.org/10.3390/molecules22101791
AMA Style
Lee H-Y, Back K. Cadmium Disrupts Subcellular Organelles, Including Chloroplasts, Resulting in Melatonin Induction in Plants. Molecules. 2017; 22(10):1791. https://doi.org/10.3390/molecules22101791
Chicago/Turabian StyleLee, Hyoung-Yool, and Kyoungwhan Back. 2017. "Cadmium Disrupts Subcellular Organelles, Including Chloroplasts, Resulting in Melatonin Induction in Plants" Molecules 22, no. 10: 1791. https://doi.org/10.3390/molecules22101791