Hepatoprotective Effects of Kaempferol-3-O-α-l-Arabinopyranosyl-7-O-α-l-Rhamnopyranoside on d-Galactosamine and Lipopolysaccharide Caused Hepatic Failure in Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Plant Material
2.3. Extraction and Purification
2.4. Animals and Treatment
2.5. Histological Analysis and Detection of Apoptotic Cells (TUNEL Assay)
2.6. Serum ALT/AST Analysis
2.7. ELISA Assay
2.8. Western Blot Assay
2.9. Statistical Analysis
3. Results
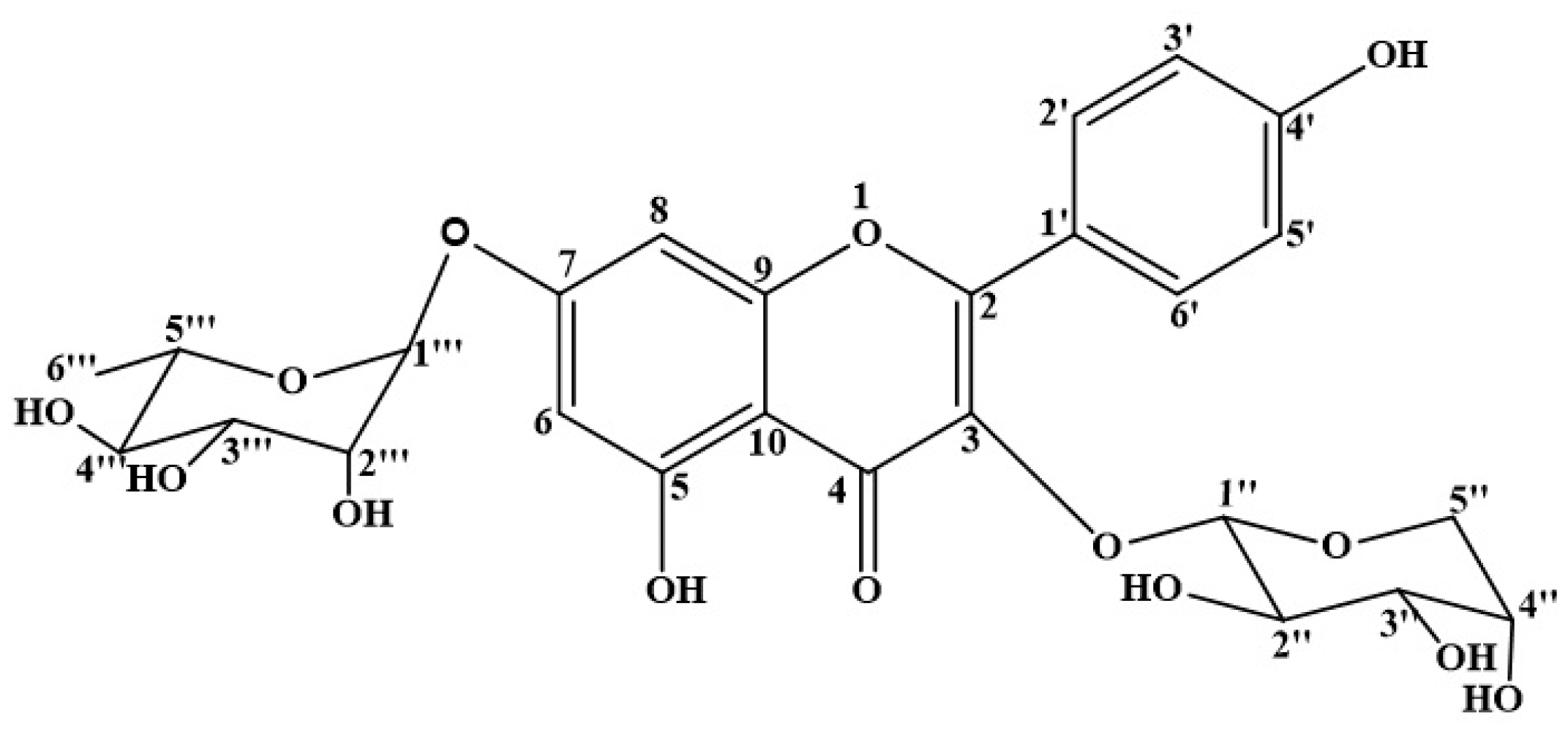
3.1. Chemical Structure of KAR
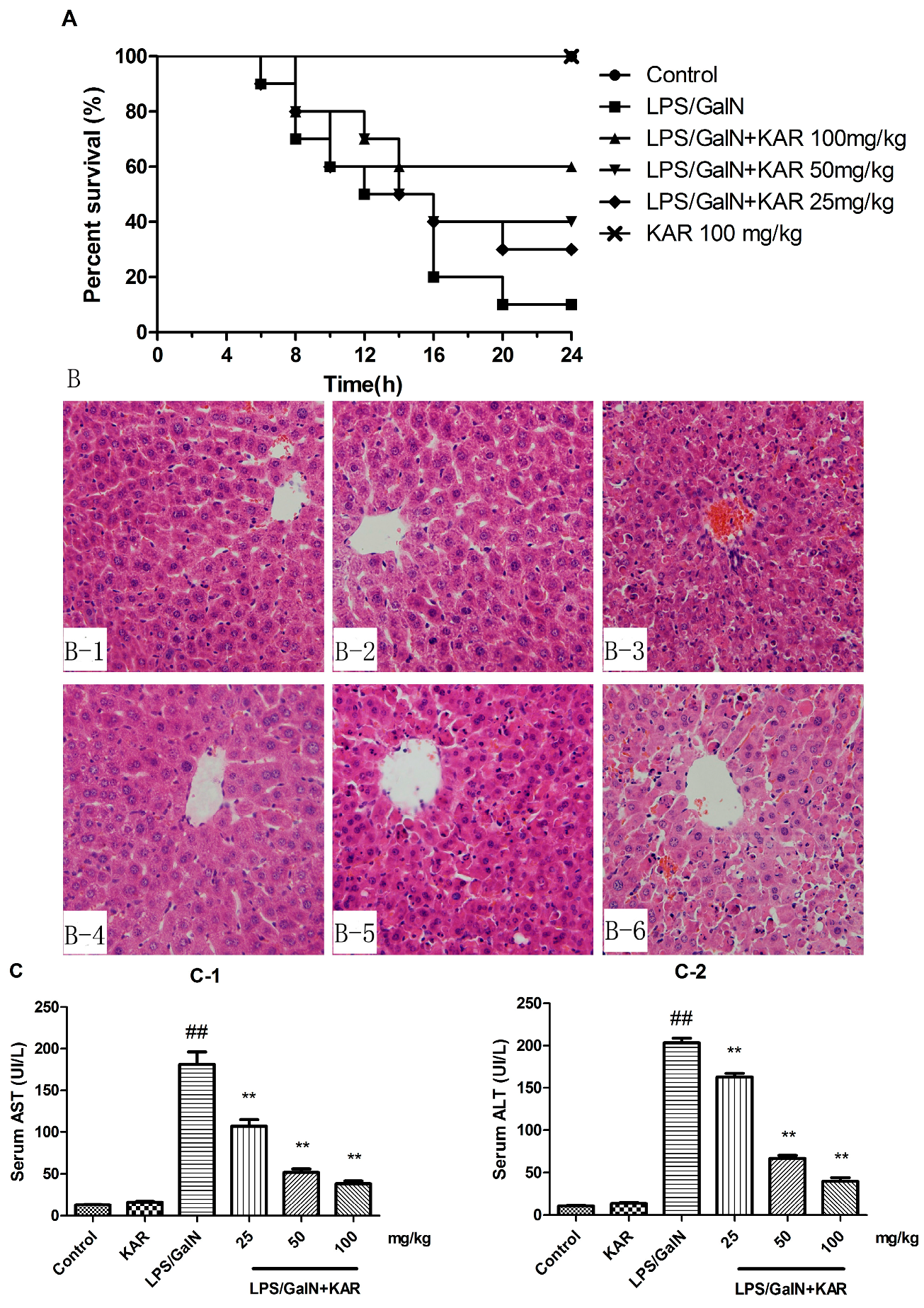
3.2. KAR Reduces GalN/LPS-Induced Mortality
3.3. KAR Protects the Normal Liver Structure and Decreases the Levels of Serum AST and ALT
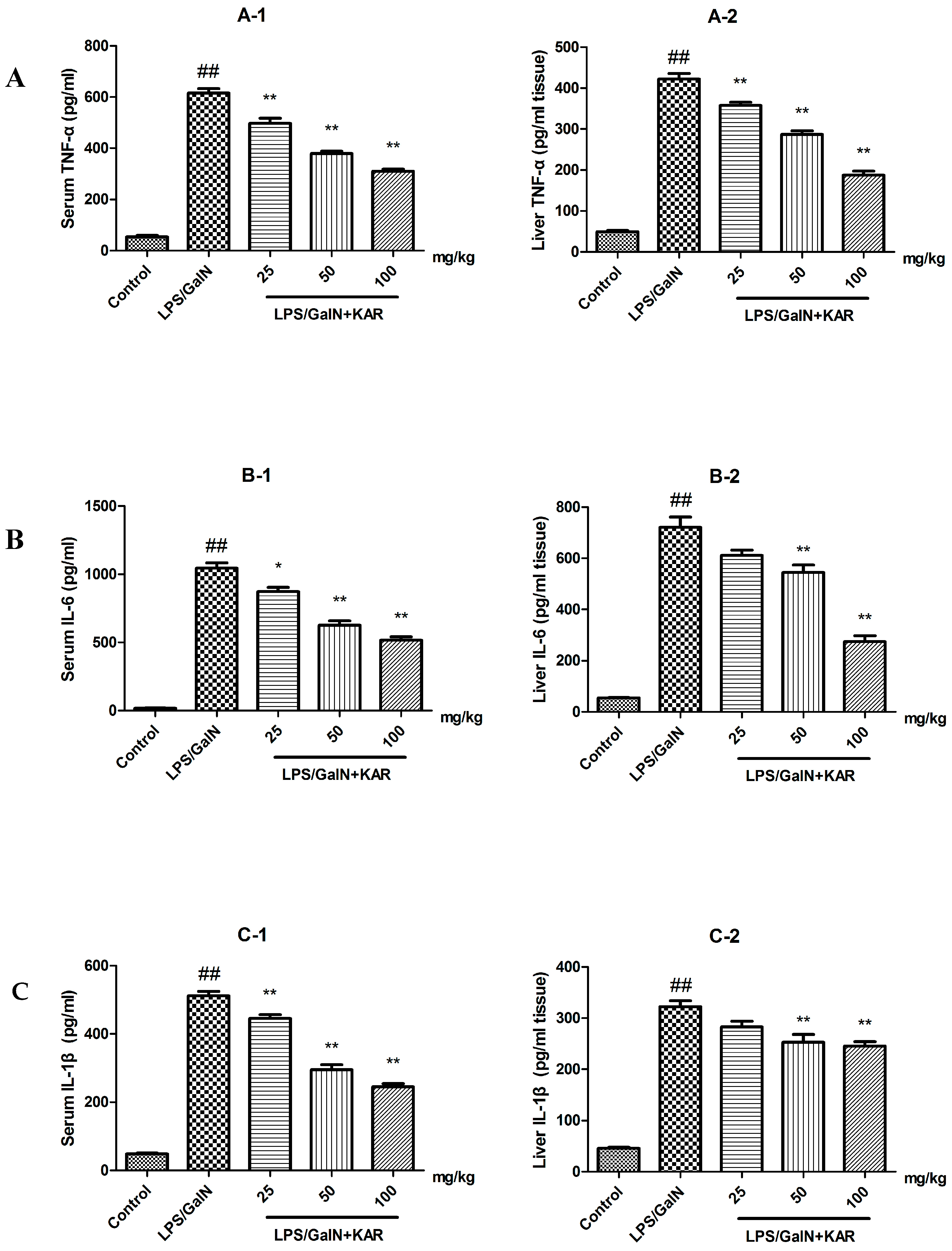
3.4. KAR Decreases the Production of Peroxidation Products and Inflammatory Cytokines
3.5. KAR Inhibits GalN/LPS-Induced Apoptosis of Hepatocytes
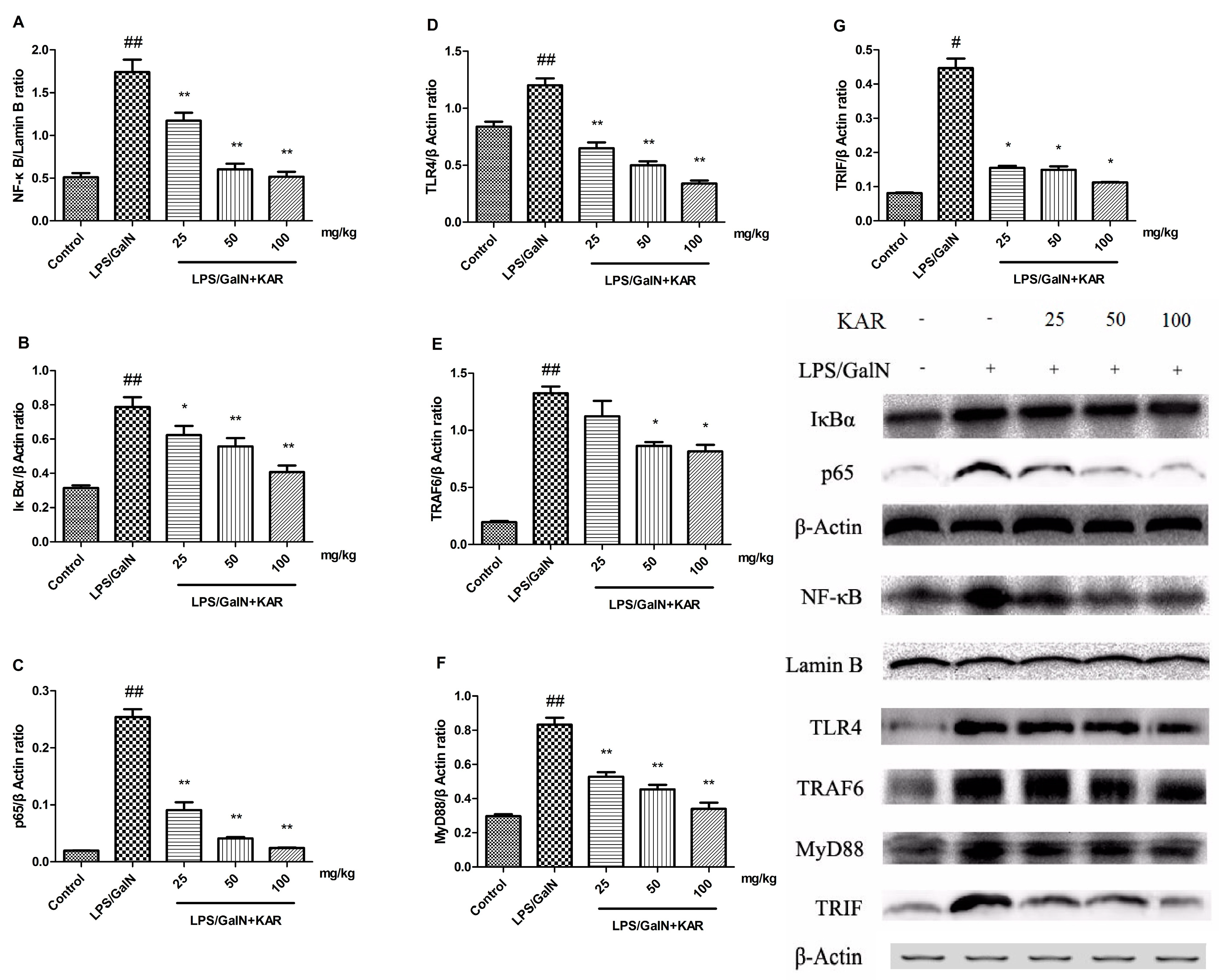
3.6. KAR Reduces the Expression of TLR4 and NF-κB Signaling Pathway Related Proteins
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Whitehouse, T.; Wendon, J. Acute liver failure. Best Pract. Res. Clini. Gastroenterol. 2013, 27, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Jiang, S.; Wu, Y.-L.; Bai, T.; Yang, Y.; Jin, X.; Lian, L.-H.; Nan, J.-X. Hepatoprotective effect of cryptotanshinone from Salvia miltiorrhiza in d-galactosamine/lipopolysaccharide-induced fulminant hepatic failure. Phytomedicine 2014, 21, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Ai, G.; Huang, Z.M.; Liu, Q.C.; Han, Y.Q.; Chen, X. The protective effect of total phenolics from Oenanthe Javanica on acute liver failure induced by D-galactosamine. J. Ethnopharmacol. 2016, 186, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Frau, R.; Bini, V.; Soggiu, A.; Scheggi, S.; Pardu, A.; Fanni, S.; Roncada, P.; Puligheddu, M.; Marrosu, F.; Caruso, D.; et al. The Neurosteroidogenic Enzyme 5alpha-Reductase Mediates Psychotic-Like Complications of Sleep Deprivation. Neuropsychopharmacology 2017, 42, 2196–2205. [Google Scholar] [CrossRef] [PubMed]
- Ganai, A.A.; Husain, M. Genistein Alleviates Neuroinflammation and Restores Cognitive Function in Rat Model of Hepatic Encephalopathy: Underlying Mechanisms. Mol. Neurobiol. 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Yang, Y.; Huang, L.; Zhang, Q.; Wan, R.Z.; Zhang, P.; Zhang, B. Antioxidation, anti-inflammation and anti-apoptosis by paeonol in LPS/d-GalN-induced acute liver failure in mice. Int. Immunopharmacol. 2017, 46, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, S.; You, S.; Liu, T.; Xu, F.; Ji, T.; Gu, Z. Hepatoprotective Effects of Nicotiflorin from Nymphaea candida against Concanavalin A-Induced and D-Galactosamine-Induced Liver Injury in Mice. Int. J. Mol. Sci. 2017, 18, 587. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Yang, R.; Dong, C.; Yang, Q. Physicochemical Properties and Fatty Acid Profiles of Elaeagnus mollis Diels Nut Oils. J. Oleo Sci. 2015, 64, 1267–1272. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qin, Y.; Du, Z.; Yan, G. Genetic diversity and differentiation of the endangered tree Elaeagnus mollis Diels (Elaeagnus L.) as revealed by Simple Sequence Repeat (SSR) Markers. Biochem. Syst. Ecol. 2012, 40, 25–33. [Google Scholar] [CrossRef]
- Liu, Y.J.; Su, J.X.; Du, X.L.; Zhang, P.; Wei, X.Z. Megasporogenesis, Microsporogenesis and development of Gametophytes of Elaeagnus mollis. Sci. Silvae Sin. 2008, 44, 39–44. [Google Scholar]
- Chen, M.; Wang, T.; Jiang, Z.Z.; Shan, C.; Wang, H.; Wu, M.J.; Zhang, S.; Zhang, Y.; Zhang, L.Y. Anti-inflammatory and hepatoprotective effects of total flavonoid C-glycosides from Abrus mollis extracts. Chin. J. Nat. Med. 2014, 12, 590–598. [Google Scholar] [CrossRef]
- Cho, H.I.; Park, J.H.; Choi, H.S.; Kwak, J.H.; Lee, D.U.; Lee, S.K.; Lee, S.M. Protective mechanisms of acacetin against D-galactosamine and lipopolysaccharide-induced fulminant hepatic failure in mice. J. Nat. Prod. 2014, 77, 2497–2503. [Google Scholar] [CrossRef] [PubMed]
- Gevrenova, R.; Kondeva-Burdina, M.; Denkov, N.; Zheleva-Dimitrova, D. Flavonoid profiles of three Bupleurum species and in vitro hepatoprotective of activity Bupleurum flavum Forsk. Pharmacogn. Mag. 2015, 11, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Yao, X.; Zeng, H.; Zhou, G.; Chen, Y.; Ma, B.; Wang, Y. Hepatoprotective activity of flavonoids from Cichorium glandulosum seeds in vitro and in vivo carbon tetrachloride-induced hepatotoxicity. J. Ethnopharmacol. 2015, 174, 355–363. [Google Scholar] [CrossRef] [PubMed]
- AHashem, N.; Soliman, M.S.; Hamed, M.A.; Swilam, N.F.; Lindequist, U.; Nawwar, M.A. Beta vulgaris subspecies cicla var. flavescens (Swiss chard): Flavonoids, hepatoprotective and hypolipidemic activities. Pharmazie 2016, 71, 227–232. [Google Scholar]
- Kim, T.W.; Lee, D.R.; Choi, B.K.; Kang, H.K.; Jung, J.Y.; Lim, S.W.; Yang, S.H.; Suh, J.W. Hepatoprotective effects of polymethoxyflavones against acute and chronic carbon tetrachloride intoxication. Food Chem. Toxicol. 2016, 91, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Yusifova, D.Y.; Movsumov, I.S.; Garaev, E.A.; Mahiou-Leddet, V.; Mabrouki, F.; Herbette, G.; Baghdikian, B.; Ollivier, E. Biologically Active Compounds from Lepidium campestre and Pulp from Lemon-Juice Production. Chem. Nat. Compd. 2015, 51, 964–965. [Google Scholar] [CrossRef]
- Galanos, C.; Freudenberg, M.A.; Reutter, W. Galactosamine-induced sensitization to the lethal effects of endotoxin. Proc. Natl. Acad. Sci. USA 1979, 76, 5939–5943. [Google Scholar] [CrossRef] [PubMed]
- Jaeschke, H. Inflammation in response to hepatocellular apoptosis. Hepatology 2002, 35, 964–966. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.-W.; Zhou, G.-Y.; Chen, W.-L.; Zhuge, L.; Jin, L.-X.; Zheng, Y.; Lin, W.; Pan, Z.-Z. Protective effect of forsythiaside A on lipopolysaccharide/d-galactosamine-induced liver injury. Int. Immunopharmacol. 2015, 26, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.-I.; Hong, J.-M.; Choi, J.-W.; Choi, H.-S.; Hwan Kwak, J.; Lee, D.-U.; Kook Lee, S.; Lee, S.-M. β-Caryophyllene alleviates d-galactosamine and lipopolysaccharide-induced hepatic injury through suppression of the TLR4 and RAGE signaling pathways. Eur. J. Pharmacol. 2015, 764, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Liaskou, E.; Wilson, D.V.; Oo, Y.H. Innate immune cells in liver inflammation. Mediators Inflamm. 2012, 2012, 949157. [Google Scholar] [CrossRef] [PubMed]
- Fiuza, C.; Suffredini, A.F. Human models of innate immunity: Local and systemic inflammatory responses. J. Endotoxin Res. 2001, 7, 385–388. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Arras, D.; Rose-John, S. IL-6 pathway in the liver: From physiopathology to therapy. J. Hepatol. 2016, 64, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- Yano, A.; Higuchi, S.; Tsuneyama, K.; Fukami, T.; Nakajima, M.; Yokoi, T. Involvement of immune-related factors in diclofenac-induced acute liver injury in mice. Toxicology 2012, 293, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Bataller, R.; Rombouts, K.; Altamirano, J.; Marra, F. Fibrosis in alcoholic and nonalcoholic steatohepatitis. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yu, S.; Jiang, Z.; Liang, C.; Yu, W.; Li, J.; Du, X.; Wang, H.; Gao, X.; Wang, X. N-acetyl-serotonin protects HepG2 cells from oxidative stress injury induced by hydrogen peroxide. Oxid. Med. Cell. Longev. 2014, 2014, 310504. [Google Scholar] [CrossRef] [PubMed]
- Guesmi, F.; Tyagi, A.K.; Bellamine, H.; Landoulsi, A. Antioxidant Machinery Related to Decreased MDA Generation by Thymus Algeriensis Essential Oil-induced Liver and Kidney Regeneration. Biomed. Environ. Sci. 2016, 29, 639–649. [Google Scholar] [PubMed]
- Liang, Q.; Sheng, Y.; Jiang, P.; Ji, L.; Xia, Y.; Min, Y.; Wang, Z. The gender-dependent difference of liver GSH antioxidant system in mice and its influence on isoline-induced liver injury. Toxicology 2011, 280, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Yang, X. Proteases for cell suicide: Functions and regulation of caspases. Microbiol. Mol. Biol. Rev. 2000, 64, 821–846. [Google Scholar] [CrossRef] [PubMed]
- Johnstone, R.W.; Ruefli, A.A.; Lowe, S.W. Apoptosis: A link between cancer genetics and chemotherapy. Cell 2002, 108, 153–164. [Google Scholar] [CrossRef]
- Leist, M.; Jäättelä, M. Four deaths and a funeral: From caspases to alternative mechanisms. Nat. Rev. Mol. Cell Biol. 2001, 2, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Lockshin, R.A.; Zakeri, Z. Programmed cell death and apoptosis: Origins of the theory. Nat. Rev. Mol. Cell Biol. 2001, 2, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Meier, P.; Finch, A.; Evan, G. Apoptosis in development. Nature 2000, 407, 796–801. [Google Scholar] [CrossRef] [PubMed]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Harrington, H.A.; Ho, K.L.; Ghosh, S.; Tung, K.C. Construction and analysis of a modular model of caspase activation in apoptosis. Theor. Biol. Med. Model. 2008, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Budihardjo, I.; Lutter, M.; Luo, X.; Wang, X. Biochemical pathways of caspase activation during apoptosis. Annu. Rev. Cell Dev. Biol. 1999, 15, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; May, M.J.; Kopp, E.B. NF-kappa B and Rel proteins: Evolutionarily conserved mediators of immune responses. Annu. Rev. Immunol. 1998, 16, 225–260. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Su, C.; Fu, J.; Zhang, P.; Jiang, X.; Xu, D.; Hu, L.; Song, E.; Song, Y. Role of α-lipoic acid in LPS/d-GalN induced fulminant hepatic failure in mice: Studies on oxidative stress, inflammation and apoptosis. Int. Immunopharmacol. 2014, 22, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Zhang, L.; Jiang, R.; Wang, C.D.; Yin, X.R.; Wan, J.Y. Hepatoprotective effects of syringin on fulminant hepatic failure induced by D-galactosamine and lipopolysaccharide in mice. J. Appl. Toxicol. 2014, 34, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Takeda, K.; Kaisho, T.; Akira, S. Toll-like receptors. Annu. Rev. Immunol. 2003, 21, 335–376. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sun, R. Toll-like receptors in acute liver injury and regeneration. Int. Immunopharmacol. 2011, 11, 1433–1441. [Google Scholar] [CrossRef] [PubMed]
- Kitazawa, T.; Tsujimoto, T.; Kawaratani, H.; Fujimoto, M.; Fukui, H. Expression of Toll-like receptor 4 in various organs in rats with D-galactosamine-induced acute hepatic failure. J. Gastroenterol. Hepatol. 2008, 23, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Broering, R.; Lu, M.; Schlaak, J.F. Role of Toll-like receptors in liver health and disease. Clin. Sci. 2011, 121, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Hritz, I.; Mandrekar, P.; Velayudham, A.; Catalano, D.; Dolganiuc, A.; Kodys, K.; KurtJones, E.; Szabo, G. The critical role of toll-like receptor (TLR) 4 in alcoholic liver disease is independent of the common TLR adapter MyD88. Hepatology 2008, 48, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Ben Ari, Z.; Avlas, O.; Pappo, O.; Zilbermints, V.; Cheporko, Y.; Bachmetov, L.; Zemel, R.; Shainberg, A.; Sharon, E.; Grief, F.; et al. Reduced Hepatic Injury in Toll-Like Receptor 4-Deficient Mice Following D-Galactosamine/Lipopolysaccharide-Induced Fulminant Hepatic Failure. Cell. Physiol. Biochem. 2012, 29, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.W.; Kim, D.W.; Choi, J.S.; Kim, Y.S.; Lee, S.M. Scoparone attenuates D-galactosamine/lipopolysaccharide-induced fulminant hepatic failure through inhibition of toll-like receptor 4 signaling in mice. Food Chem. Toxicol. 2013, 57, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.N.; Piseteky, D.S.; Schwartz, D.A. Toll-like receptors in the pathogenesis of human disease. Nat. Immunol. 2004, 5, 975–979. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, M.D.; Yamahina, S.; Froh, M.; Rusyn, I.; Thurman, R.G. Adenoviral gene delivery can inactivate Kupffer cells: Role of oxidants in NF-kappaB activation and cytokine production. J. Leukoc. Biol. 2001, 69, 622–630. [Google Scholar] [PubMed]
- Sato, S.; Sugiyama, M.; Yamamoto, M.; Watanabe, Y.; Kawai, T.; Takeda, K.; Akira, S. Toll/IL-1 receptor domain-containing adaptor inducing IFN-beta (TRIF) associates with TNF receptor-associated factor 6 and TANK-binding kinase 1, and activates two distinct transcription factors, NF-kappa B and IFN-regulatory factor-3, in the Toll-like receptor signaling. J. Immunol. 2003, 171, 4304–4310. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds Kaempferol-3-O-α-l-Arabinopyranosyl-7-O-α-l-Rhamnopyranoside are available from the authors. |



© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Yin, L.; Quan, H.; Chu, Y.; Lu, J. Hepatoprotective Effects of Kaempferol-3-O-α-l-Arabinopyranosyl-7-O-α-l-Rhamnopyranoside on d-Galactosamine and Lipopolysaccharide Caused Hepatic Failure in Mice. Molecules 2017, 22, 1755. https://doi.org/10.3390/molecules22101755
Dong L, Yin L, Quan H, Chu Y, Lu J. Hepatoprotective Effects of Kaempferol-3-O-α-l-Arabinopyranosyl-7-O-α-l-Rhamnopyranoside on d-Galactosamine and Lipopolysaccharide Caused Hepatic Failure in Mice. Molecules. 2017; 22(10):1755. https://doi.org/10.3390/molecules22101755
Chicago/Turabian StyleDong, Lin, Lei Yin, Hongfeng Quan, Yuankui Chu, and Jincai Lu. 2017. "Hepatoprotective Effects of Kaempferol-3-O-α-l-Arabinopyranosyl-7-O-α-l-Rhamnopyranoside on d-Galactosamine and Lipopolysaccharide Caused Hepatic Failure in Mice" Molecules 22, no. 10: 1755. https://doi.org/10.3390/molecules22101755