Immobilization of Moniliella spathulata R25L270 Lipase on Ionic, Hydrophobic and Covalent Supports: Functional Properties and Hydrolysis of Sardine Oil

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Immobilization of M. spathulata R25L270 Lipaseon Different Supports
2.2. Characterization of M. spathulata R25L270 Lipase Immobilized Derivatives
2.2.1. Apparent Kinetic Parameters (Vmax and KM)
2.2.2. pH Stability
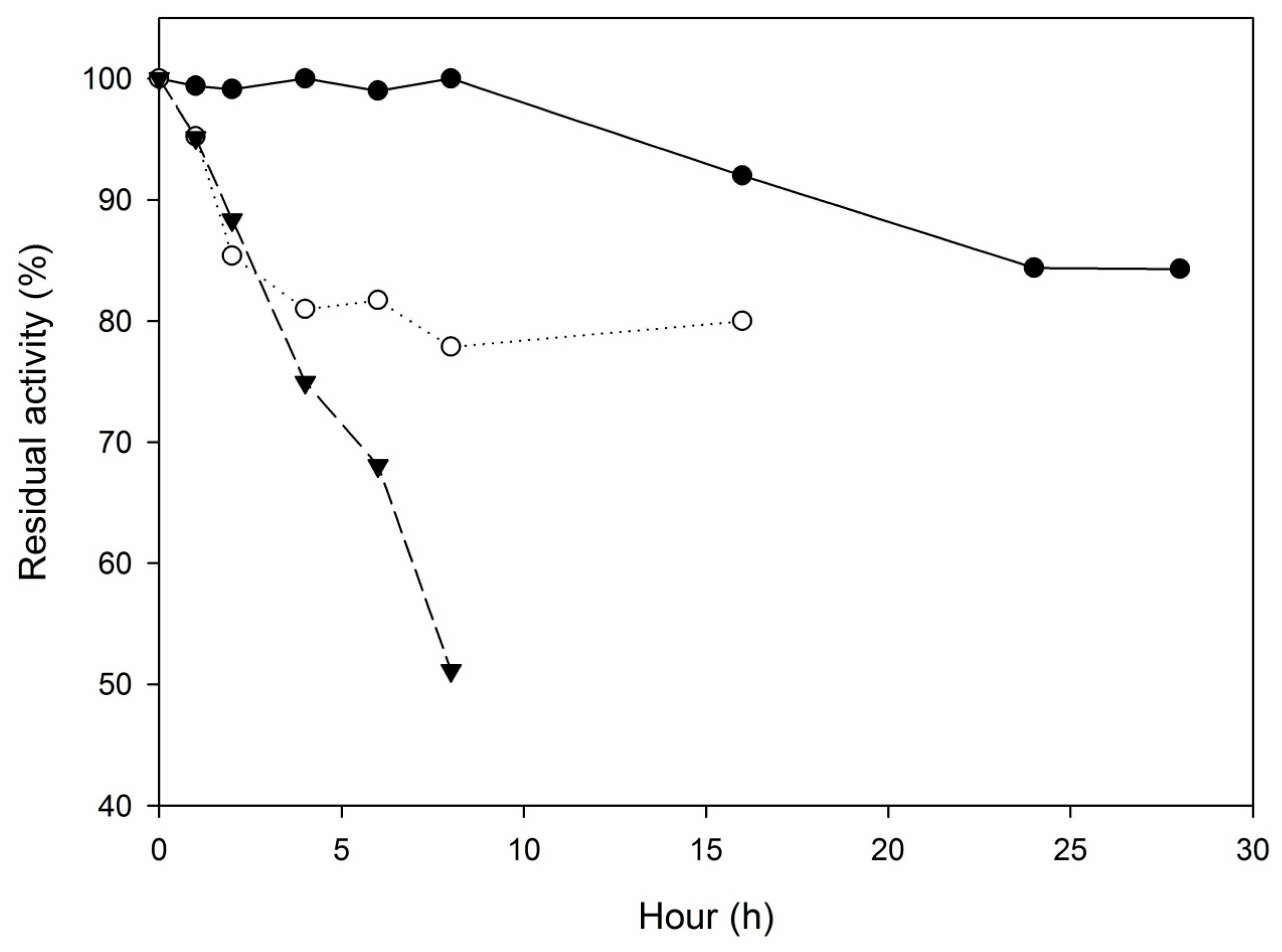
2.2.3. Thermostability
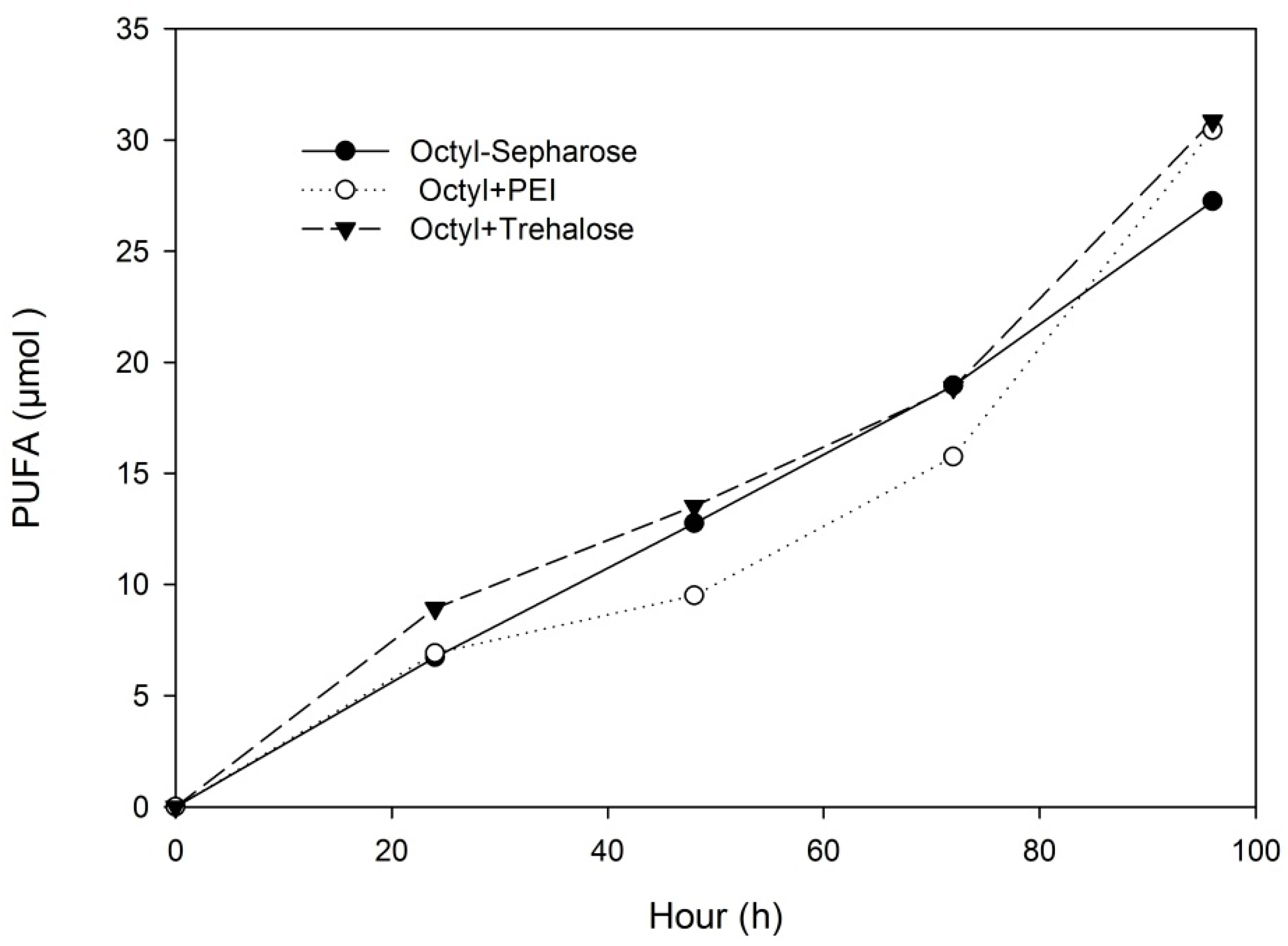
2.3. Hydrolysis of Sardine Oil by M. spathulata R25L270 Lipase Immobilized on Octyl-Sepharose
3. Materials and Methods
3.1. Materials
3.2. Lipase Production using Macaúba Cake as Substrate
3.3. Lipase Activity Assay
3.4. Immobilization of M. spathulata R25L270 Lipase by Adsorption on Hydrophobic and Ionic Supports
3.5. Immobilization of M. spathulata R25l270Lipase by Covalent Attachment on CNBr-Activated Sepharose
3.6. Determination of Immobilization Kinetic Parameters
3.7. Biochemical Characterization of M. Spathulata R25L270 Lipase Immobilized on Different Supports
3.7.1. Kinetic Parameters Estimation
3.7.2. pH Stability
3.7.3. Thermostability
3.8. Sardine Oil Hydrolysis in Biphasic System
4. Conclusions
Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| SF | Stabilization factors |
| EPA | eicosapentaenoic acid |
| PUFA | polyunsaturated fatty acids |
| DHA | docosahexaenoic acid |
| pNPP | p-nitrophenylpalmitate |
| pNP | p-nitrophenol |
| PEI | polyethyleneimine |
| CNBr | Cyanogen bromide-activated Sepharose |
| Y | Immobilization yield |
| RA | recovered activity |
| Kd | deactivation rate constant |
| t1/2 | half-life time |
| MANAE | monoaminoethyl-N-aminoethyl |
| DEAE | diethylaminoethyl |
| SP | sulfopropyl |
References
- Souza, L.T.A.; Oliveira, J.S.; Rodrigues, M.Q.R.B.; Dos Santos, V.L.; Pessela, B.C.; Resende, R.R. Macaúba (Acrocomia aculeata) cake from biodiesel processing: A low-cost substrate to produce lipases from Moniliella spathulata R25L270 with potential application in the oleochemical industry. Microb. Cell Fact. 2015, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Adlercreutz, P. Immobilisation and application of lipases in organic media. Chem. Soc. Rev. 2013, 42, 6406–6436. [Google Scholar] [CrossRef] [PubMed]
- Zinjarde, S.S. Food-related applications of Yarrowia lipolytica. Food Chem. 2014, 152, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.; Lu, J.; Nie, K.; Deng, L.; Wang, F. Biodiesel production with immobilized lipase: A review. Biotechnol. Adv. 2010, 28, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Murty, V.R.; Bhat, J.; Muniswaran, P.K. Hydrolysis of oils by using immobilized lipase enzyme: A review. Biotechnol. Bioprocess Eng. 2002, 7, 57–66. [Google Scholar] [CrossRef]
- Kapoor, M.; Gupta, M.N. Lipase promiscuity and its biochemical applications. Process Biochem. 2012, 47, 555–569. [Google Scholar] [CrossRef]
- Jaeger, K.E.; Reetz, M.T. Microbial lipases form versatile tools for biotechnology. Trends Biotechnol. 1998, 16, 396–403. [Google Scholar] [CrossRef]
- Moreno-Pérez, S.; Guisan, J.M.; Fernandez-Lorente, G. Selective ethanolysis of fish oil catalyzed by immobilized lipases. J. Am. Oil Chem. Soc. 2014, 91, 63–69. [Google Scholar] [CrossRef]
- Mohammadi, M.; Habibi, Z.; Dezvarei, S.; Yousefi, M.; Ashjari, M. Selective enrichment of polyunsaturated fatty acids by hydrolysis of fish oil using immobilized and stabilized Rhizomucor miehei lipase preparations. Food Bioprod. Process. 2014, 94, 4–11. [Google Scholar] [CrossRef]
- Fernández-Lorente, G.; Betancor, L.; Carrascosa, A.V.; Palomo, J.M.; Guisan, J.M. Modulation of the selectivity of immobilized lipases by chemical and physical modifications: Release of omega-3 fatty acids from fish oil. J. Am. Oil Chem. Soc. 2012, 89, 97–102. [Google Scholar] [CrossRef]
- Fernández-Lorente, G.; Pizarro, C.; López-Vela, D.; Betancor, L.; Carrascosa, A.V.; Pessela, B.; Guisan, J.M. Hydrolysis of Fish Oil by Lipases Immobilized Inside Porous Supports. J. Am. Oil Chem. Soc. 2010, 88, 819–826. [Google Scholar] [CrossRef] [Green Version]
- Pizarro, C.; Brañes, M.C.; Markovits, A.; Fernandez-Lorente, G.; Guisán, J.M.; Chamy, R.; Wilson, L. Influence of different immobilization techniques for Candida cylindracea lipase on its stability and fish oil hydrolysis. J. Mol. Catal. B Enzym. 2012, 78, 111–118. [Google Scholar] [CrossRef]
- Byun, H.G.; Eom, T.K.; Jung, W.K.; Kim, S.K. Lipase-catalyzed hydrolysis of fish oil in an optimum emulsion system. Biotechnol. Bioprocess Eng. 2007, 12, 484–490. [Google Scholar] [CrossRef]
- Phillips, M.A. Omega-3 fatty acids, cognitive impairment and Alzheimer’s disease. Rev. Clin. Gerontol. 2010, 20, 219–238. [Google Scholar] [CrossRef]
- Manerba, A.; Vizzardi, E.; Metra, M.; Dei Cas, L. n-3 PUFAs and cardiovascular disease prevention. Future Cardiol. 2010, 6, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Fernández-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.T.A.; Mendes, A.A.; de Castro, H.F. Selection of lipases for the synthesis of biodiesel from Jatropha Oil and the potential of microwave irradiation to enhance the reaction rate. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Cipolatti, E.P.; Moreno-Pérez, S.; Souza, L.T.A.; Valério, A.; Guisán, J.M.; Araújo, P.H.H.; Sayer, C.; Ninow, J.L.; Oliveira, D.; Pessela, B.C. Synthesis and modification of polyurethane for immobilization of Thermomyces lanuginosus (TLL) lipase for ethanolysis of fish oil in solvent free system. J. Mol. Catal. B Enzym. 2015, 122, 163–169. [Google Scholar] [CrossRef]
- Marciello, M.; Bolivar, J.M.; Filice, M.; Mateo, C.; Guisan, J.M. Preparation of lipase-coated, stabilized, hydrophobic magnetic particles for reversible conjugation of biomacromolecules. Biomacromolecules 2013, 14, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Martins, A.B.; Friedrich, J.L.R.; Cavalheiro, J.C.; Garcia-Galan, C.; Barbosa, O.; Ayub, M.Z.; Fernandez-Lafuente, R.; Rodrigues, R.C. Improved production of butyl butyrate with lipase from Thermomyces lanuginosus immobilized on styrene-divinylbenzene beads. Bioresour. Technol. 2013, 134, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Chiou, S.-H.; Wu, W.-T. Immobilization of Candida rugosa lipase on chitosan with activation of the hydroxyl groups. Biomaterials 2004, 25, 197–204. [Google Scholar] [CrossRef]
- Kumar, D.; Nagar, S.; Bhushan, I.; Kumar, L.; Parshad, R.; Gupta, V.K. Covalent immobilization of organic solvent tolerant lipase on aluminum oxide pellets and its potential application in esterification reaction. J. Mol. Catal. B Enzym. 2013, 87, 51–61. [Google Scholar] [CrossRef]
- Pereira, M.G.; Facchini, F.D.A.; Filó, L.E.C.; Polizeli, A.M.; Vici, A.C.; Jorge, J.A.; Pessela, B.C.; Guisan, J.M.; de Moraes Polizeli, M.d.L. Immobilized lipase from Hypocrea pseudokoningii on hydrophobic and ionic supports: Determination of thermal and organic solvent stabilities for applications in the oleochemical industry. Process Biochem. 2015, 50, 1–10. [Google Scholar] [CrossRef]
- Volpato, G.; Filice, M.; Ayub, M.A.Z.; Guisan, J.M.; Palomo, J.M. Single-step purification of different lipases from Staphylococcus warneri. J. Chromatogr. A 2010, 1217, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Cunha, A.G.; Fernández-Lorente, G.; Gutarra, M.L.E.; Bevilaqua, J.V.; Almeida, R.V.; Paiva, L.M.C.; Fernández-Lafuente, R.; Guisán, J.M.; Freire, D.M.G. Separation and immobilization of lipase from Penicillium simplicissimum by selective adsorption on hydrophobic supports. Appl. Biochem. Biotechnol. 2009, 156, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Lorente, G.; Ortiz, C.; Segura, R.L.; Fernández-Lafuente, R.; Guisán, J.M.; Palomo, J.M. Purification of different lipases from Aspergillus niger by using a highly selective adsorption on hydrophobic supports. Biotechnol. Bioeng. 2005, 92, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Bastida, A.; Sabuquillo, P.; Armisen, P.; Huguet, J.; Guisa, M. A single step purification, immobilization, and hyperactivation of lipases via interfacial adsorption on strongly hydrophobic supports. Biotechnol. Bioeng. 1998, 58, 486–493. [Google Scholar] [CrossRef]
- Manoel, E.; dos Santos, J.C.S.; Freire, D.M.G.; Rueda, N.; Fernandez-Lafuente, R. Immobilization of lipases on hydrophobic supports involves the open form of the enzyme. Enzyme Microb. Technol. 2015, 71, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Abian, O.; Bernedo, M.; Cuenca, E.; Fuentes, M.; Fernandez-Lorente, G.; Palomo, J.M.; Grazu, V.; Pessela, B.C.C.; Giacomini, C.; et al. Some special features of glyoxyl supports to immobilize proteins. Enzyme Microb. Technol. 2005, 37, 456–462. [Google Scholar] [CrossRef]
- Dandavate, V.; Keharia, H.; Madamwar, D. Ethyl isovalerate synthesis using Candida rugosa lipase immobilized on silica nanoparticles prepared in nonionic reverse micelles. Process Biochem. 2009, 44, 349–352. [Google Scholar] [CrossRef]
- Izrael Živković, L.T.; Živković, L.S.; Babić, B.M.; Kokunešoski, M.J.; Jokić, B.M.; Karadžić, I.M. Immobilization of Candida rugosa lipase by adsorption onto biosafe meso/macroporous silica and zirconia. Biochem. Eng. J. 2015, 93, 73–83. [Google Scholar] [CrossRef]
- Zhang, W.; Qing, W.; Ren, Z.; Li, W.; Chen, J. Lipase immobilized catalytically active membrane for synthesis of lauryl stearate in a pervaporation membrane reactor. Bioresour. Technol. 2014, 172C, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Urioste, D.; Andrade Souza, L.T.; Mendes, A.; de Castro, H.F. Assessment of the Morphological, Biochemical, and Kinetic Properties for Candida rugosa lipase immobilized on hydrous Niobium oxide to be used in the biodiesel synthesis. Enzyme Res. 2011, 2011, 216453. [Google Scholar] [CrossRef] [PubMed]
- Sadana, A.; Henley, J.P. Single-step unimolecular non-first-order enzyme deactivation kinetics. Biotechnol. Bioeng. 1987, 30, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Abian, O.; Fernandez-Lafuente, R.; Guisan, J.M. Increase in conformational stability of enzymes immobilized on epoxy-activated supports by favoring additional multipoint covalent attachment. Enzyme Microb. Technol. 2000, 26, 509–515. [Google Scholar] [CrossRef]
- Souza, L.; Oliveira, J.S.; Santos, V.L.; Regis, W.C.B.; Santoro, M.M.; Resende, R.R. Lipolytic potential of Aspergillus japonicus LAB01: Production, partial purification, and characterisation of an extracellular lipase. BioMed Res. Int. 2014, 2014, 1080913. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |

{kind=link}
{kind=link}
| Immobilization Method | Derivative | Hydrolytic Activity (U/g) | Immobilization Yield (%) | Recovered Activity (%) |
|---|---|---|---|---|
| Hydrophobic Adsorption | Butyl-Sepharose | 5.82 ± 0.11 | 64.80 | 95.80 |
| Phenyl-Sepharose | 5.33 ± 0.06 | 52.30 | 87.80 | |
| Octyl-Sepharose | 3.87 ± 0.02 | 63.98 | 63.80 | |
| Octyl + polyethyleneimine (PEI) | 4.41± 0.08 | 63.98 | 72.55 | |
| Octyl + Trehalose | 3.94 ± 0.05 | 63.98 | 64.92 | |
| Ionic Adsorption | MANAE (Monoaminoethyl-N-aminoethyl)-Agarose | 1.32 ± 0.23 | 63.97 | 21.80 |
| DEAE-(Diethylaminoethyl)-Agarose | 2.46 ± 0.38 | 75.20 | 40.51 | |
| SP-(Sulfopropyl)-Sepharose | 1.10 ± 0.04 | 22.29 | 18.19 | |
| Covalent Attachment | CNBr-(Cyanogenogen Bromide)-activated Sepharose | 1.38 ± 0.02 | 36.27 | 22.73 |
| Derivative | Michaelis Constant KM (mM) | Maximum Velocity Vmax (µmoles/min) | R2 |
|---|---|---|---|
| Free | 1.92 | 3.15 | 0.98 |
| Octyl-Sepharose | 1.35 | 2.48 | 0.99 |
| Octyl + PEI | 1.38 | 2.38 | 0.97 |
| Octyl + Trehalose | 1.66 | 2.79 | 0.99 |
| DEAE-Agarose | 0.40 | 2.01 | 0.98 |
| CNBr-activated Sepharose | 0.10 | 1.03 | 0.99 |
| Residual Activity (%) | |||
|---|---|---|---|
| Immobilized Derivative | pH 5.0 | pH 7.0 | pH 9.0 |
| Free | 62 ± 1 | 102 ± 2 | 105 ± 1 |
| Butyl-Sepharose | 0 | 96 ± 13 | 99 ± 14 |
| Phenyl-Sepharose | 85 ± 4 | 91 ± 11 | 106 ± 1 |
| Octyl-Sepharose | 100 ± 8 | 120 ± 4 | 113 ± 10 |
| Octyl + PEI | 96 ± 1 | 108 ± 5 | 112 ± 7 |
| Octyl + Trehalose | 89 ± 15 | 128 ± 18 | 105 ± 1 |
| DEAE-Agarose | 0 | 76 ± 1 | 104 ± 5 |
| CNBr-activated Sepharose | 0 | 122 ± 16 | 121 ± 15 |
| Derivative | Kd (h−1) | t1/2 (h) | Stabilization Factor (SF) | R2 |
|---|---|---|---|---|
| Free | 1.72 | 0.40 | - | 0.99 |
| Octyl-Sepharose | 0.17 | 4.15 | 10.30 | 0.99 |
| Butyl-Sepharose | 0.65 | 1.06 | 2.63 | 0.98 |
| Phenyl-Sepharose | 0.29 | 2.39 | 5.92 | 0.98 |
| DEAE-Agarose | 2.21 | 0.31 | 0.77 | 1.00 |
| Selectivity a | |||
|---|---|---|---|
| Derivative | pH 5.0 | pH 7.0 | pH 9.0 |
| Octyl-Sepharose | 7.53 | 3.33 | 3.03 |
| Octyl + PEI | 6.05 | 4.14 | 2.39 |
| Octyl + Trehalose | 4.34 | 4.23 | 2.83 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, L.T.d.A.; Moreno-Perez, S.; Fernández Lorente, G.; Cipolatti, E.P.; De Oliveira, D.; Resende, R.R.; Pessela, B.C. Immobilization of Moniliella spathulata R25L270 Lipase on Ionic, Hydrophobic and Covalent Supports: Functional Properties and Hydrolysis of Sardine Oil. Molecules 2017, 22, 1508. https://doi.org/10.3390/molecules22101508
Souza LTdA, Moreno-Perez S, Fernández Lorente G, Cipolatti EP, De Oliveira D, Resende RR, Pessela BC. Immobilization of Moniliella spathulata R25L270 Lipase on Ionic, Hydrophobic and Covalent Supports: Functional Properties and Hydrolysis of Sardine Oil. Molecules. 2017; 22(10):1508. https://doi.org/10.3390/molecules22101508
Chicago/Turabian StyleSouza, Lívia T. de A., Sonia Moreno-Perez, Gloria Fernández Lorente, Eliane P. Cipolatti, Débora De Oliveira, Rodrigo R. Resende, and Benevides C. Pessela. 2017. "Immobilization of Moniliella spathulata R25L270 Lipase on Ionic, Hydrophobic and Covalent Supports: Functional Properties and Hydrolysis of Sardine Oil" Molecules 22, no. 10: 1508. https://doi.org/10.3390/molecules22101508