
The Effect of Capsaicin on Salivary Gland Dysfunction


{kind=link}
Abstract
:1. Introduction
2. Modulation of Salivary Secretion by Capsaicin
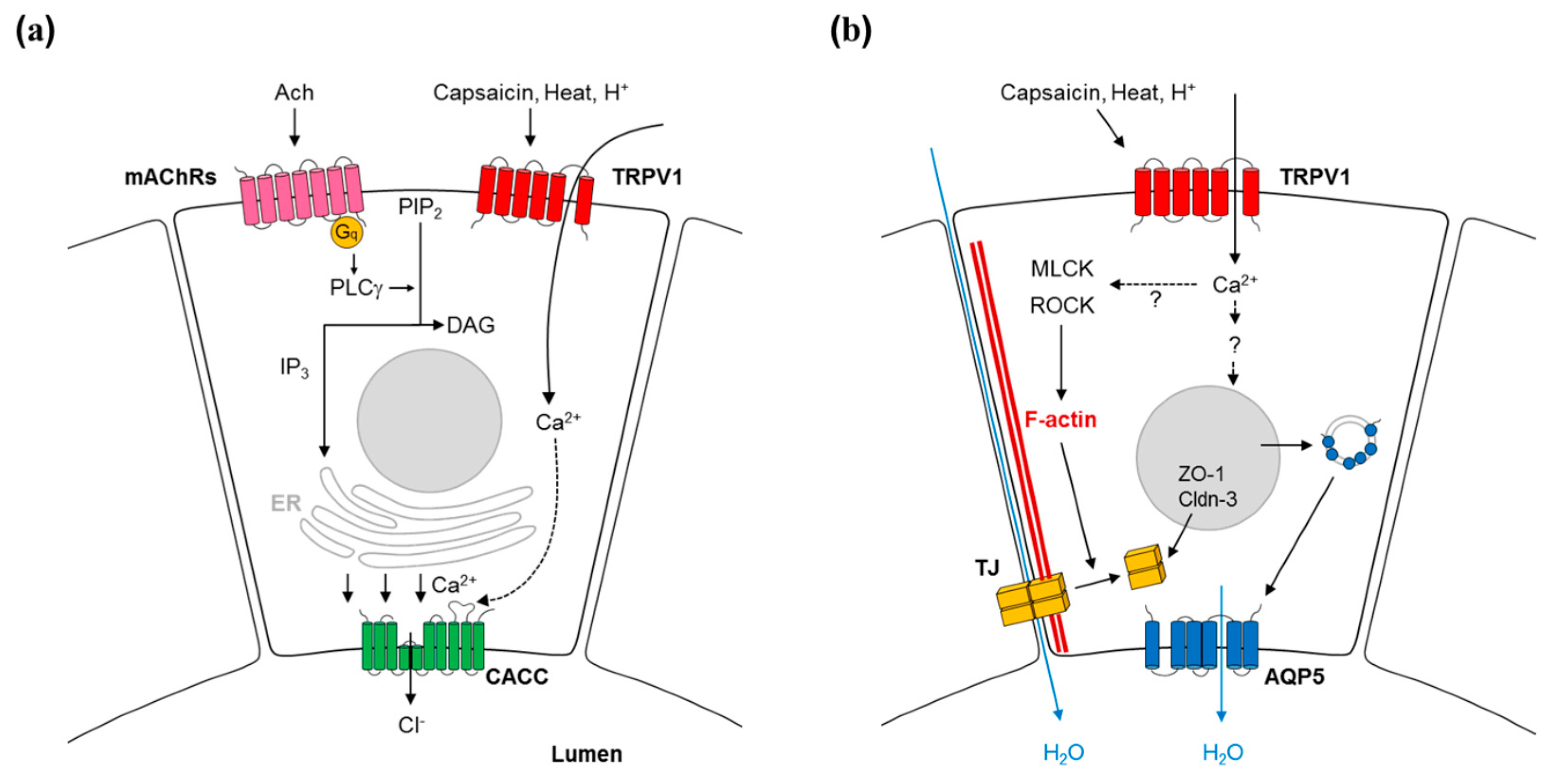
3. Mechanism of Capsaicin Action in Salivary Secretion
4. Modulation of Inflammation by Capsaicin
5. Pharmacokinetics and Therapeutic Potential of Capsaicin
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jensen, J.L.; Barkvoll, P. Clinical implications of the dry mouth. Oral mucosal diseases. Ann. N. Y. Acad. Sci. 1998, 842, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Narhi, T.O.; Meurman, J.H.; Ainamo, A. Xerostomia and hyposalivation: Causes, consequences and treatment in the elderly. Drugs Aging 1999, 15, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Strahl, R.C.; Welsh, S.; Streckfus, C.F. Salivary flow rates: A diagnostic aid in treatment planning geriatric patients. Clin. Prev. Dent 1990, 12, 10–12. [Google Scholar] [PubMed]
- Valdez, I.H.; Fox, P.C. Diagnosis and management of salivary dysfunction. Crit. Rev. Oral Biol. Med. 1993, 4, 271–277. [Google Scholar] [PubMed]
- Atkinson, J.C.; Grisius, M.; Massey, W. Salivary hypofunction and xerostomia: Diagnosis and treatment. Dent Clin. N. Am. 2005, 49, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Grisius, M.M. Salivary gland dysfunction: A review of systemic therapies. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2001, 92, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Benchabane, S.; Boudjelida, A.; Toumi, R.; Belguendouz, H.; Youinou, P.; Touil-Boukoffa, C. A case for IL-6, IL-17A, and nitric oxide in the pathophysiology of Sjogren′s syndrome. Int. J. Immunopathol. Pharmacol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Vij, A.S.; Sharma, M. Mechanisms and clinical uses of capsaicin. Eur. J. Pharmacol. 2013, 720, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Gunthorpe, M.J.; Szallasi, A. Peripheral TRPV1 receptors as targets for drug development: New molecules and mechanisms. Curr. Pharm. Des. 2008, 14, 32–41. [Google Scholar] [PubMed]
- Vadivelu, N.; Mitra, S.; Narayan, D. Recent advances in postoperative pain management. Yale J. Biol. Med. 2010, 83, 11–25. [Google Scholar] [PubMed]
- Choi, S.; Shin, Y.H.; Namkoong, E.; Hwang, S.M.; Cong, X.; Yu, G.; Park, K. TRPV1 in Salivary Gland Epithelial Cells Is Not Involved in Salivary Secretion via Transcellular Pathway. Korean J. Physiol. Pharmacol. 2014, 18, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Oyagbemi, A.A.; Saba, A.B.; Azeez, O.I. Capsaicin: A novel chemopreventive molecule and its underlying molecular mechanisms of action. Indian J. Cancer 2010, 47, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.H.; Namkoong, E.; Choi, S.; Bae, J.S.; Jin, M.; Hwang, S.M.; Arote, R.; Choi, S.Y.; Park, K. Capsaicin regulates the NF-κB pathway in salivary gland inflammation. J. Dent Res. 2013, 92, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Szallasi, A. Vanilloid receptor ligands: Hopes and realities for the future. Drugs Aging 2001, 18, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [PubMed]
- Cong, X.; Zhang, Y.; Shi, L.; Yang, N.Y.; Ding, C.; Li, J.; Ding, Q.W.; Su, Y.C.; Xiang, R.L.; Wu, L.L.; et al. Activation of transient receptor potential vanilloid subtype 1 increases expression and permeability of tight junction in normal and hyposecretory submandibular gland. Lab. Investig. 2012, 92, 753–768. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.W.; Zhang, Y.; Wang, Y.; Wang, Y.N.; Zhang, L.; Ding, C.; Wu, L.L.; Yu, G.Y. Functional vanilloid receptor-1 in human submandibular glands. J. Dent. Res. 2010, 89, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.; Aeschlimann, A.; Simmen, B.R.; Michel, B.A.; Gay, R.E.; Gay, S.; Sprott, H. Expression of transient receptor potential vanilloid 1 (TRPV1) in synovial fibroblasts from patients with osteoarthritis and rheumatoid arthritis. Biochem. Biophys. Res. Commun. 2007, 359, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Koizumi, S.; Fuziwara, S.; Denda, S.; Inoue, K.; Denda, M. Functional vanilloid receptors in cultured normal human epidermal keratinocytes. Biochem. Biophys. Res. Commun. 2002, 291, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, B.; Carter, J.D.; Devlin, R.B.; Simon, S.A.; Oortgiesen, M. Neuropeptides and capsaicin stimulate the release of inflammatory cytokines in a human bronchial epithelial cell line. Neuropeptides 1999, 33, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xiang, B.; Li, Y.M.; Wang, Y.; Wang, X.; Wang, Y.N.; Wu, L.L.; Yu, G.Y. Expression and characteristics of vanilloid receptor 1 in the rabbit submandibular gland. Biochem. Biophys. Res. Commun. 2006, 345, 467–473. [Google Scholar] [CrossRef] [PubMed]
- Melvin, J.E.; Yule, D.; Shuttleworth, T.; Begenisich, T. Regulation of fluid and electrolyte secretion in salivary gland acinar cells. Annu. Rev. Physiol. 2005, 67, 445–469. [Google Scholar] [CrossRef] [PubMed]
- Duner-Engstrom, M.; Fredholm, B.B.; Larsson, O.; Lundberg, J.M.; Saria, A. Autonomic mechanisms underlying capsaicin induced oral sensations and salivation in man. J. Physiol. 1986, 373, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Kanehira, T.; Hongou, H.; Asano, K.; Morita, M.; Maeshima, E.; Matsuda, A.; Sakamoto, W. A simple test for salivary gland function measuring resting and stimulated submandibular and sublingual secretions. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. 2014, 117, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Kanehira, T.; Yamaguchi, T.; Asano, K.; Morita, M.; Maeshima, E.; Matsuda, A.; Fujii, Y.; Sakamoto, W. A screening test for capsaicin-stimulated salivary flow using filter paper: A study for diagnosis of hyposalivation with a complaint of dry mouth. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2011, 112, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Nagumo, Y.; Han, J.; Bellila, A.; Isoda, H.; Tanaka, T. Cofilin mediates tight-junction opening by redistributing actin and tight-junction proteins. Biochem. Biophys. Res. Commun. 2008, 377, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cong, X.; Shi, L.; Xiang, B.; Li, Y.M.; Ding, Q.W.; Ding, C.; Wu, L.L.; Yu, G.Y. Activation of transient receptor potential vanilloid subtype 1 increases secretion of the hypofunctional, transplanted submandibular gland. Am. J. Physiol Gastrointest Liver Physiol 2010, 299, G54–G62. [Google Scholar] [CrossRef] [PubMed]
- Proctor, G.B.; Carpenter, G.H. Regulation of salivary gland function by autonomic nerves. Auton Neurosci. 2007, 133, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Abrams, P.; Andersson, K.E.; Buccafusco, J.J.; Chapple, C.; de Groat, W.C.; Fryer, A.D.; Kay, G.; Laties, A.; Nathanson, N.M.; Pasricha, P.J.; et al. Muscarinic receptors: Their distribution and function in body systems, and the implications for treating overactive bladder. Br. J. Pharmacol. 2006, 148, 565–578. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.; Tourwe, D.; Delaet, N.; Kramer, T.H.; Burks, T.F. In vitro evaluation of a casomorphin related pentapeptide, a competitive mu opioid receptor antagonist. Proc. West Pharmacol. Soc. 1991, 34, 429–432. [Google Scholar] [PubMed]
- Gautam, D.; Heard, T.S.; Cui, Y.; Miller, G.; Bloodworth, L.; Wess, J. Cholinergic stimulation of salivary secretion studied with M1 and M3 muscarinic receptor single- and double-knockout mice. Mol. Pharmacol. 2004, 66, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Proctor, G.B.; Carpenter, G.H. Salivary secretion: Mechanism and neural regulation. Monogr. Oral Sci. 2014, 24, 14–29. [Google Scholar] [PubMed]
- Romanenko, V.G.; Catalan, M.A.; Brown, D.A.; Putzier, I.; Hartzell, H.C.; Marmorstein, A.D.; Gonzalez-Begne, M.; Rock, J.R.; Harfe, B.D.; Melvin, J.E. Tmem16A encodes the Ca2+-activated Cl- channel in mouse submandibular salivary gland acinar cells. J. Biol. Chem. 2010, 285, 12990–13001. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.D.; Cho, H.; Koo, J.Y.; Tak, M.H.; Cho, Y.; Shim, W.S.; Park, S.P.; Lee, J.; Lee, B.; Kim, B.M.; et al. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 2008, 455, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Hayes, P.; Meadows, H.J.; Gunthorpe, M.J.; Harries, M.H.; Duckworth, D.M.; Cairns, W.; Harrison, D.C.; Clarke, C.E.; Ellington, K.; Prinjha, R.K.; et al. Cloning and functional expression of a human orthologue of rat vanilloid receptor-1. Pain 2000, 88, 205–215. [Google Scholar] [CrossRef]
- Tominaga, M.; Tominaga, T. Structure and function of TRPV1. Pflugers Arch. 2005, 451, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Ambudkar, I.S. Regulation of calcium in salivary gland secretion. Crit. Rev. Oral Biol. Med. 2000, 11, 4–25. [Google Scholar] [CrossRef] [PubMed]
- Takayama, Y.; Shibasaki, K.; Suzuki, Y.; Yamanaka, A.; Tominaga, M. Modulation of water efflux through functional interaction between TRPV4 and TMEM16A/anoctamin 1. FASEB J. 2014, 28, 2238–2248. [Google Scholar] [CrossRef] [PubMed]
- Takayama, Y.; Uta, D.; Furue, H.; Tominaga, M. Pain-enhancing mechanism through interaction between TRPV1 and anoctamin 1 in sensory neurons. Proc. Natl. Acad. Sci. USA 2015, 112, 5213–5218. [Google Scholar] [CrossRef] [PubMed]
- Tsukita, S.; Furuse, M.; Itoh, M. Multifunctional strands in tight junctions. Nat. Rev. Mol. Cell Biol. 2001, 2, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Cong, X.; Zhang, Y.; Yang, N.Y.; Li, J.; Ding, C.; Ding, Q.W.; Su, Y.C.; Mei, M.; Guo, X.H.; Wu, L.L.; et al. Occludin is required for TRPV1-modulated paracellular permeability in the submandibular gland. J. Cell Sci. 2013, 126, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cong, X.; Zhang, Y.; Xiang, R.L.; Mei, M.; Yang, N.Y.; Su, Y.C.; Choi, S.; Park, K.; Zhang, L.W.; et al. ZO-1 and -2 Are Required for TRPV1-Modulated Paracellular Permeability. J. Dent Res. 2015, 94, 1748–1756. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Song, Y.; Gillespie, A.; Carlson, E.J.; Epstein, C.J.; Verkman, A.S. Defective secretion of saliva in transgenic mice lacking aquaporin-5 water channels. J. Biol. Chem. 1999, 274, 20071–20074. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, G.G.; Garcia de la Rubia, P.; Gallar, J.; Belmonte, C. Reduction of capsaicin-induced ocular pain and neurogenic inflammation by calcium antagonists. Investig. Ophthalmol. Vis Sci. 1993, 34, 3329–3335. [Google Scholar] [PubMed]
- Joe, B.; Rao, U.J.; Lokesh, B.R. Presence of an acidic glycoprotein in the serum of arthritic rats: Modulation by capsaicin and curcumin. Mol. Cell Biochem. 1997, 169, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Planells-Cases, R.; Aracil, A.; Merino, J.M.; Gallar, J.; Perez-Paya, E.; Belmonte, C.; Gonzalez-Ros, J.M.; Ferrer-Montiel, A.V. Arginine-rich peptides are blockers of VR-1 channels with analgesic activity. FEBS Lett. 2000, 481, 131–136. [Google Scholar] [CrossRef]
- Lee, I.O.; Lee, K.H.; Pyo, J.H.; Kim, J.H.; Choi, Y.J.; Lee, Y.C. Anti-inflammatory effect of capsaicin in Helicobacter pylori-infected gastric epithelial cells. Helicobacter 2007, 12, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Han, S.S.; Keum, Y.S.; Chun, K.S.; Surh, Y.J. Suppression of phorbol ester-induced NF-κB activation by capsaicin in cultured human promyelocytic leukemia cells. Arch. Pharm. Res. 2002, 25, 475–479. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Kawada, T.; Kim, B.S.; Han, I.S.; Choe, S.Y.; Kurata, T.; Yu, R. Capsaicin exhibits anti-inflammatory property by inhibiting IκB-a degradation in LPS-stimulated peritoneal macrophages. Cell Signal 2003, 15, 299–306. [Google Scholar] [CrossRef]
- Singh, S.; Natarajan, K.; Aggarwal, B.B. Capsaicin (8-methyl-N-vanillyl-6-nonenamide) is a potent inhibitor of nuclear transcription factor-κB activation by diverse agents. J. Immunol. 1996, 157, 4412–4420. [Google Scholar] [PubMed]
- Lisi, S.; Sisto, M.; Lofrumento, D.D.; D′Amore, M. Altered IkappaBalpha expression promotes NF-κB activation in monocytes from primary Sjogren′s syndrome patients. Pathology 2012, 44, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, H.; Kawakami, A.; Ida, H.; Koji, T.; Eguchi, K. EGF activates PI3K-Akt and NF-κB via distinct pathways in salivary epithelial cells in Sjogren′s syndrome. Rheumatol. Int. 2007, 28, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Ping, L.; Ogawa, N.; Zhang, Y.; Sugai, S.; Masaki, Y.; Weiguo, X. p38 mitogen-activated protein kinase and nuclear factor-κB facilitate CD40-mediated salivary epithelial cell death. J. Rheumatol. 2012, 39, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Sisto, M.; Lisi, S.; Lofrumento, D.D.; D′Amore, M.; Frassanito, M.A.; Ribatti, D. Sjogren′s syndrome pathological neovascularization is regulated by VEGF-A-stimulated TACE-dependent crosstalk between VEGFR2 and NF-κB. Genes Immun. 2012, 13, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Villalon, L.; Mamani, M.; Romanini, F.E.; Catalan Pellet, A.; Berra, A. Primary Sjogren′s syndrome: Expression of NF-κB in minor salivary glands). Reumatol. Clin. 2010, 6, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Takada, Y.; Boriek, A.M.; Aggarwal, B.B. Nuclear factor-κB: Its role in health and disease. J. Mol. Med. (Berl.) 2004, 82, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Hayden, M.S.; Ghosh, S. Signaling to NF-κB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [PubMed]
- Oeckinghaus, A.; Hayden, M.S.; Ghosh, S. Crosstalk in NF-κB signaling pathways. Nat. Immunol. 2011, 12, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Chaiyasit, K.; Khovidhunkit, W.; Wittayalertpanya, S. Pharmacokinetic and the effect of capsaicin in Capsicum frutescens on decreasing plasma glucose level. J. Med. Assoc. Thai. 2009, 92, 108–113. [Google Scholar] [PubMed]
- Pershing, L.K.; Reilly, C.A.; Corlett, J.L.; Crouch, D.J. Effects of vehicle on the uptake and elimination kinetics of capsaicinoids in human skin in vivo. Toxicol. Appl. Pharmacol. 2004, 200, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Babbar, S.; Chanda, S.; Bley, K. Inhibition and induction of human cytochrome P450 enzymes in vitro by capsaicin. Xenobiotica 2010, 40, 807–816. [Google Scholar] [CrossRef] [PubMed]
- Kulkantrakorn, K.; Lorsuwansiri, C.; Meesawatsom, P. 0.025% capsaicin gel for the treatment of painful diabetic neuropathy: A randomized, double-blind, crossover, placebo-controlled trial. Pain Pract. 2013, 13, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef] [PubMed]
- Durmuşlar, M.C.; Türer, A.; Ballı, U.; Yılmaz, Z.; Önger, M.E.; Çelik, H.H.; Vatansever, A. The effect of infliximab on bone healing in osteoporotic rats. Eur. J. Inflamm. 2016, 14, 54–60. [Google Scholar] [CrossRef]
- Wang, M.; Ji, P.; Wang, R.; Zhao, L.; Xia, Z. TRPV1 agonist capsaicin attenuates lung ischemia-reperfusion injury in rabbits. J. Surg. Res. 2012, 173, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Backonja, M.M.; Malan, T.P.; Vanhove, G.F.; Tobias, J.K. NGX-4010, a high-concentration capsaicin patch, for the treatment of postherpetic neuralgia: A randomized, double-blind, controlled study with an open-label extension. Pain Med. 2010, 11, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, S. Advances in the management of diabetic peripheral neuropathy. Curr. Opin. Support. Palliat. Care 2009, 3, 136–143. [Google Scholar] [CrossRef] [PubMed]

© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, Y.-H.; Kim, J.M.; Park, K. The Effect of Capsaicin on Salivary Gland Dysfunction. Molecules 2016, 21, 835. https://doi.org/10.3390/molecules21070835
Shin Y-H, Kim JM, Park K. The Effect of Capsaicin on Salivary Gland Dysfunction. Molecules. 2016; 21(7):835. https://doi.org/10.3390/molecules21070835
Chicago/Turabian StyleShin, Yong-Hwan, Jin Man Kim, and Kyungpyo Park. 2016. "The Effect of Capsaicin on Salivary Gland Dysfunction" Molecules 21, no. 7: 835. https://doi.org/10.3390/molecules21070835
APA StyleShin, Y.-H., Kim, J. M., & Park, K. (2016). The Effect of Capsaicin on Salivary Gland Dysfunction. Molecules, 21(7), 835. https://doi.org/10.3390/molecules21070835