
Wild Bitter Melon Leaf Extract Inhibits Porphyromonas gingivalis-Induced Inflammation: Identification of Active Compounds through Bioassay-Guided Isolation

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
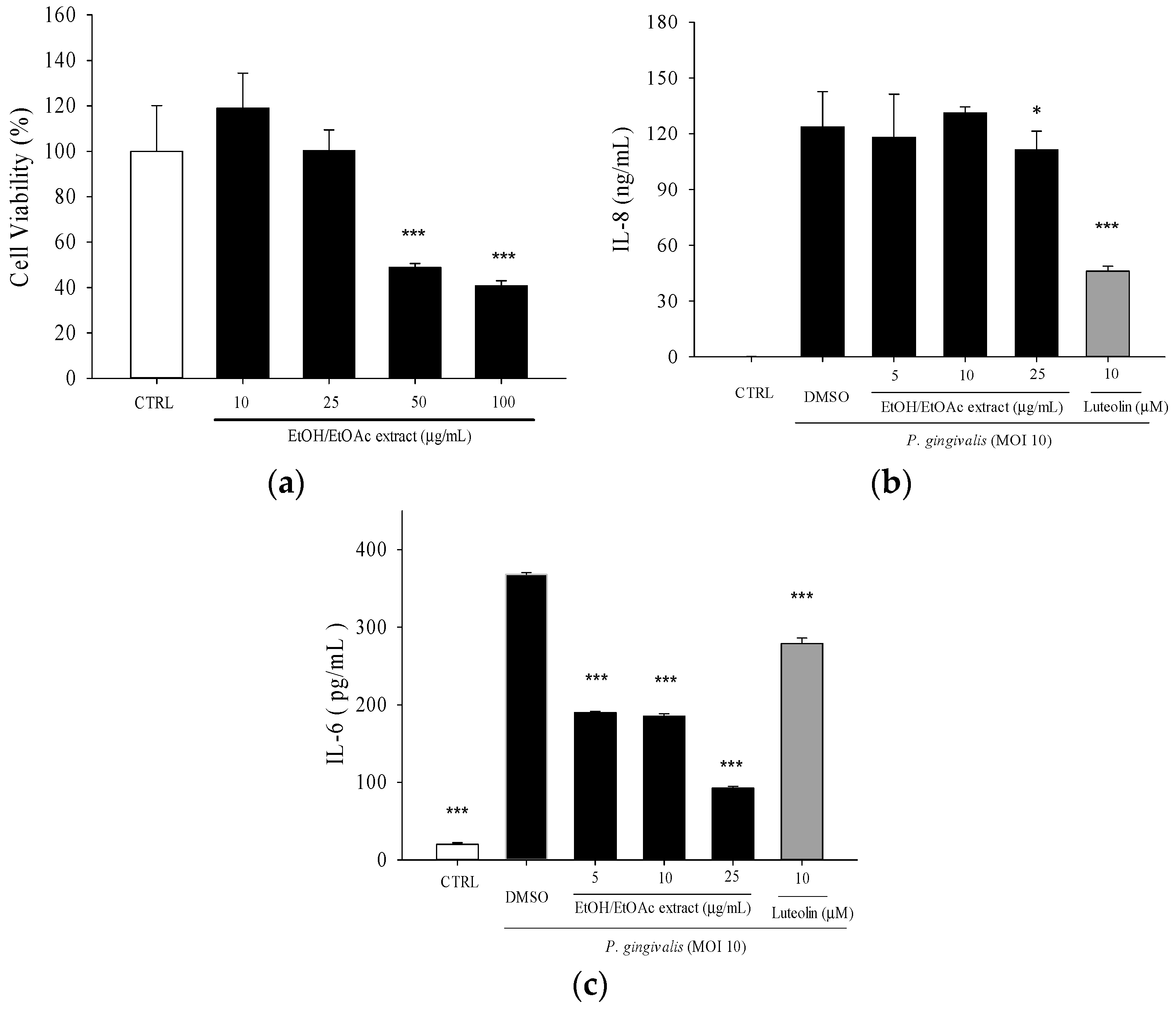
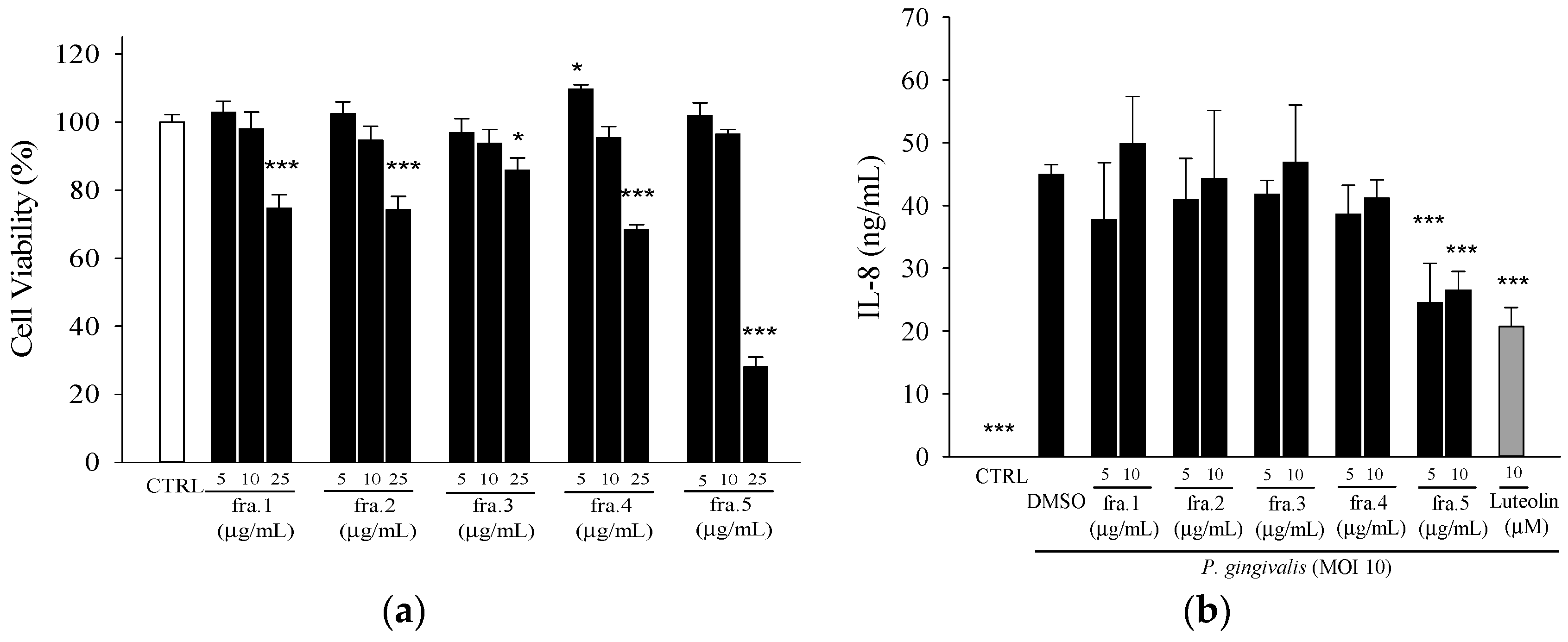
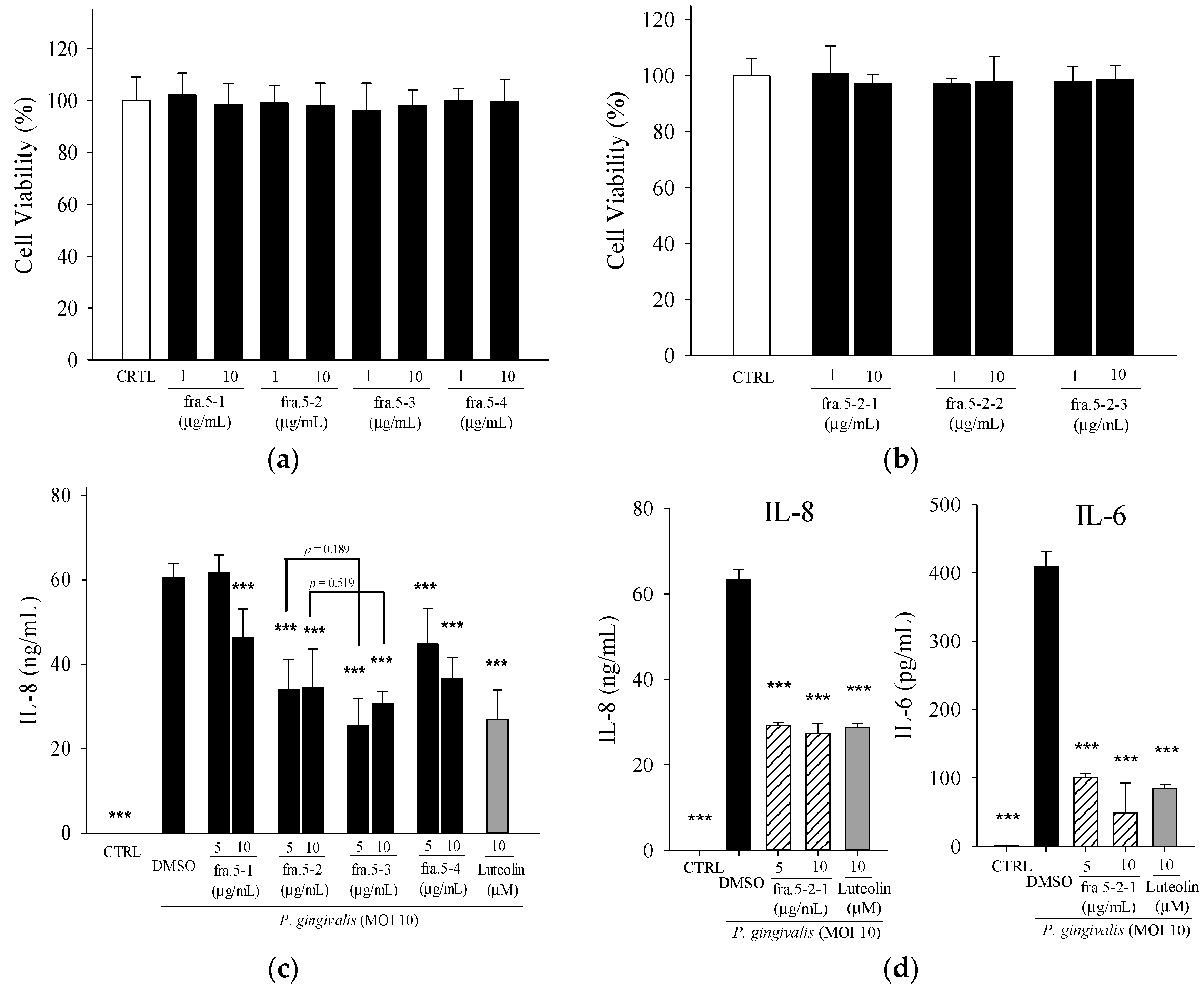
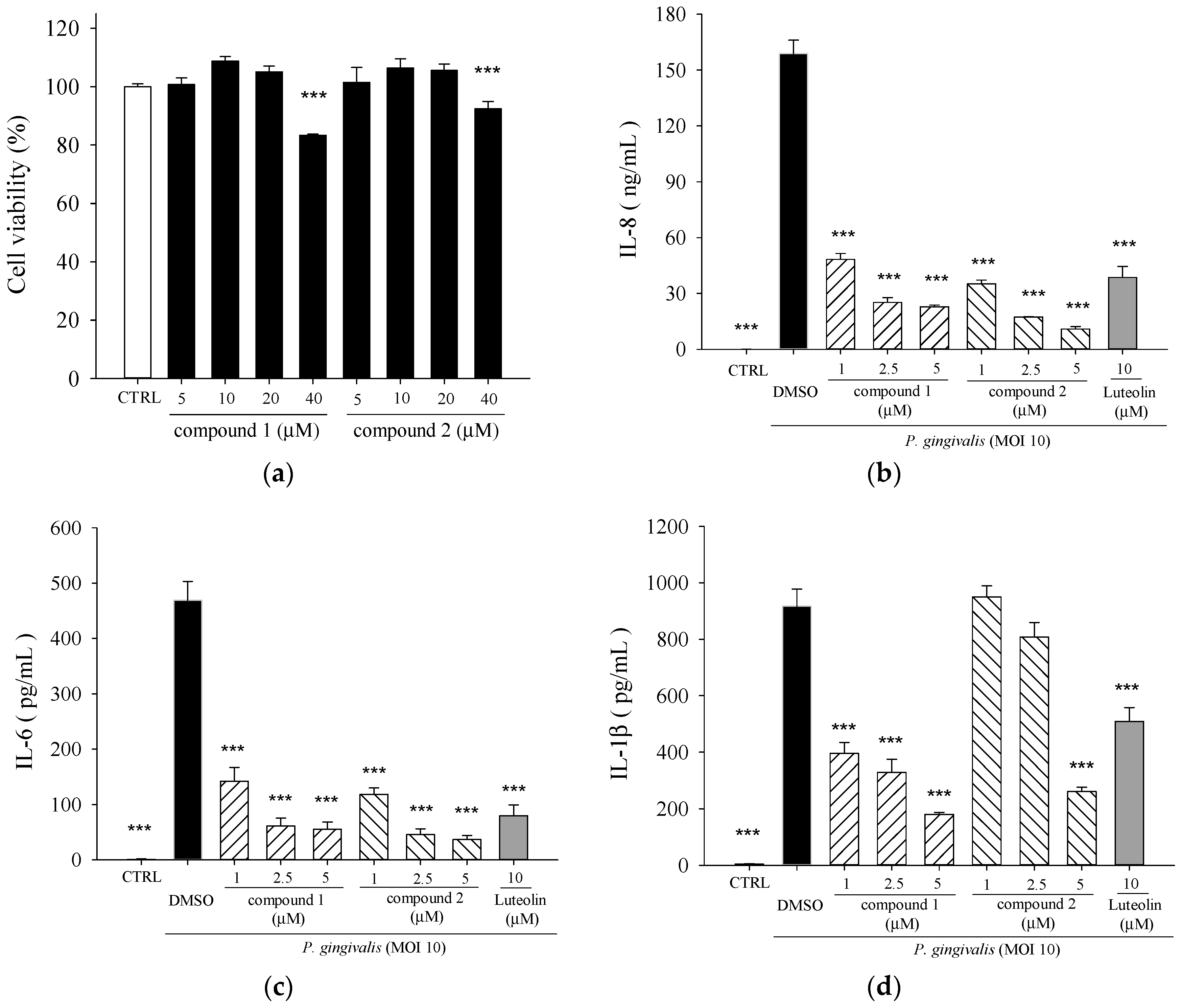
2. Results
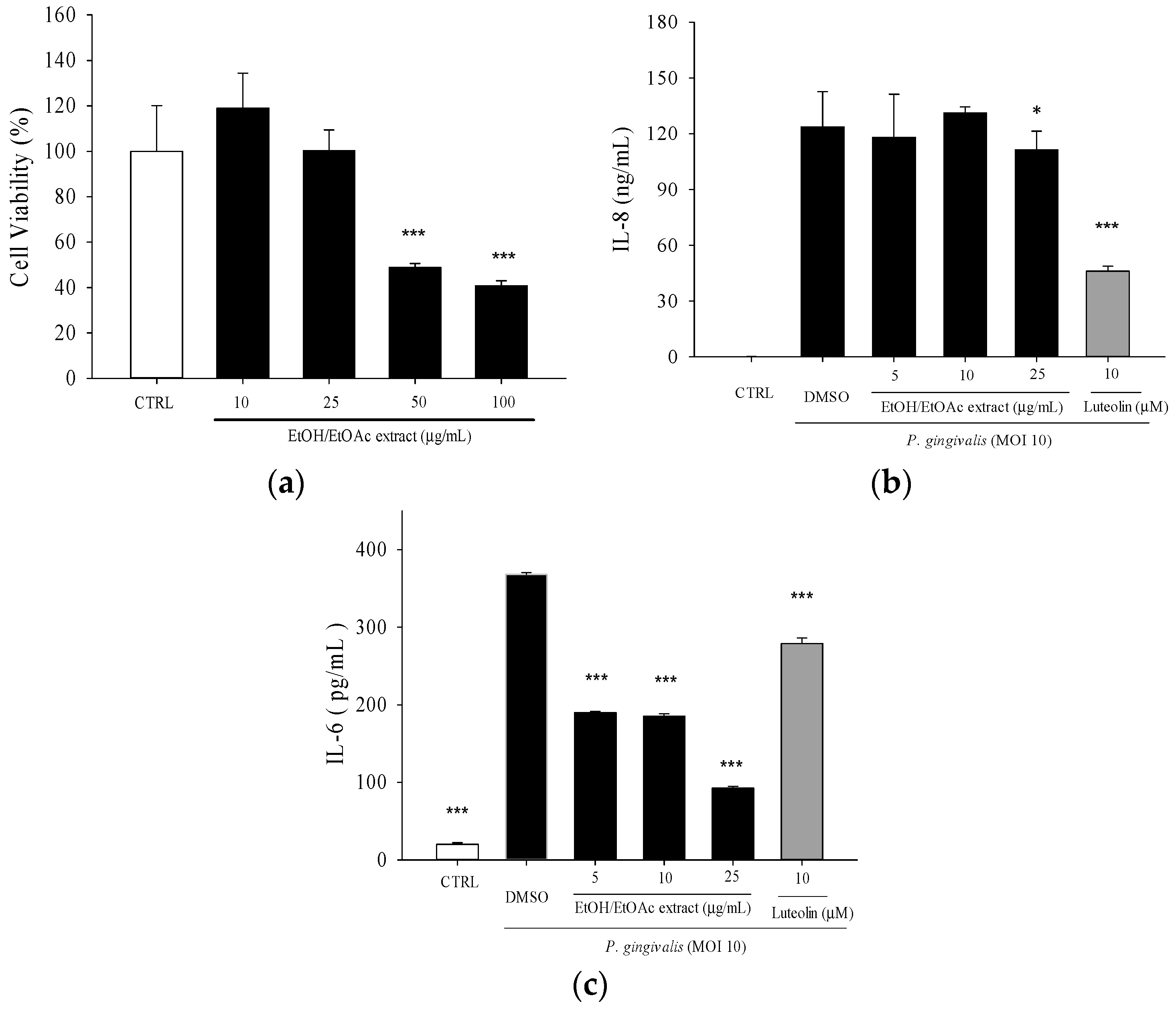
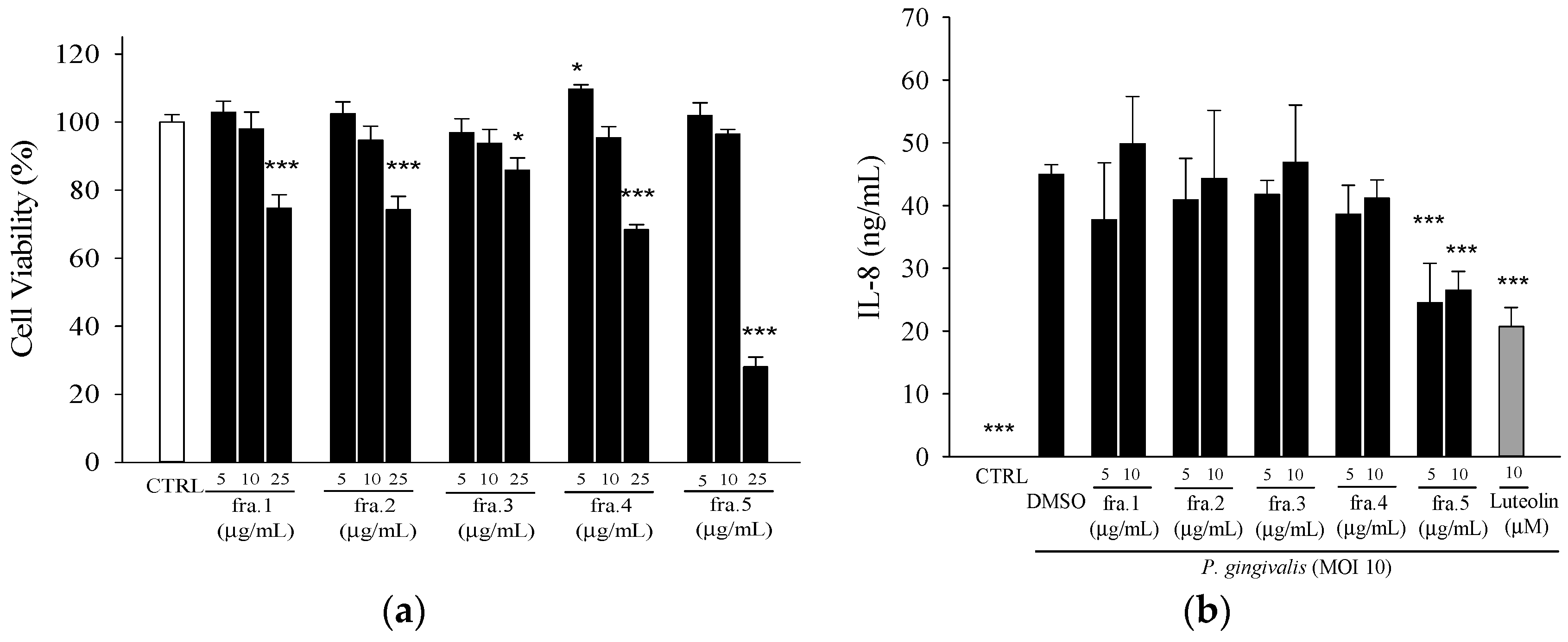
2.1. Effects of Leaf Extract of Wild Bitter Melon and Its Subfractions on P. gingivalis-Induced Cytokines in THP-Cells
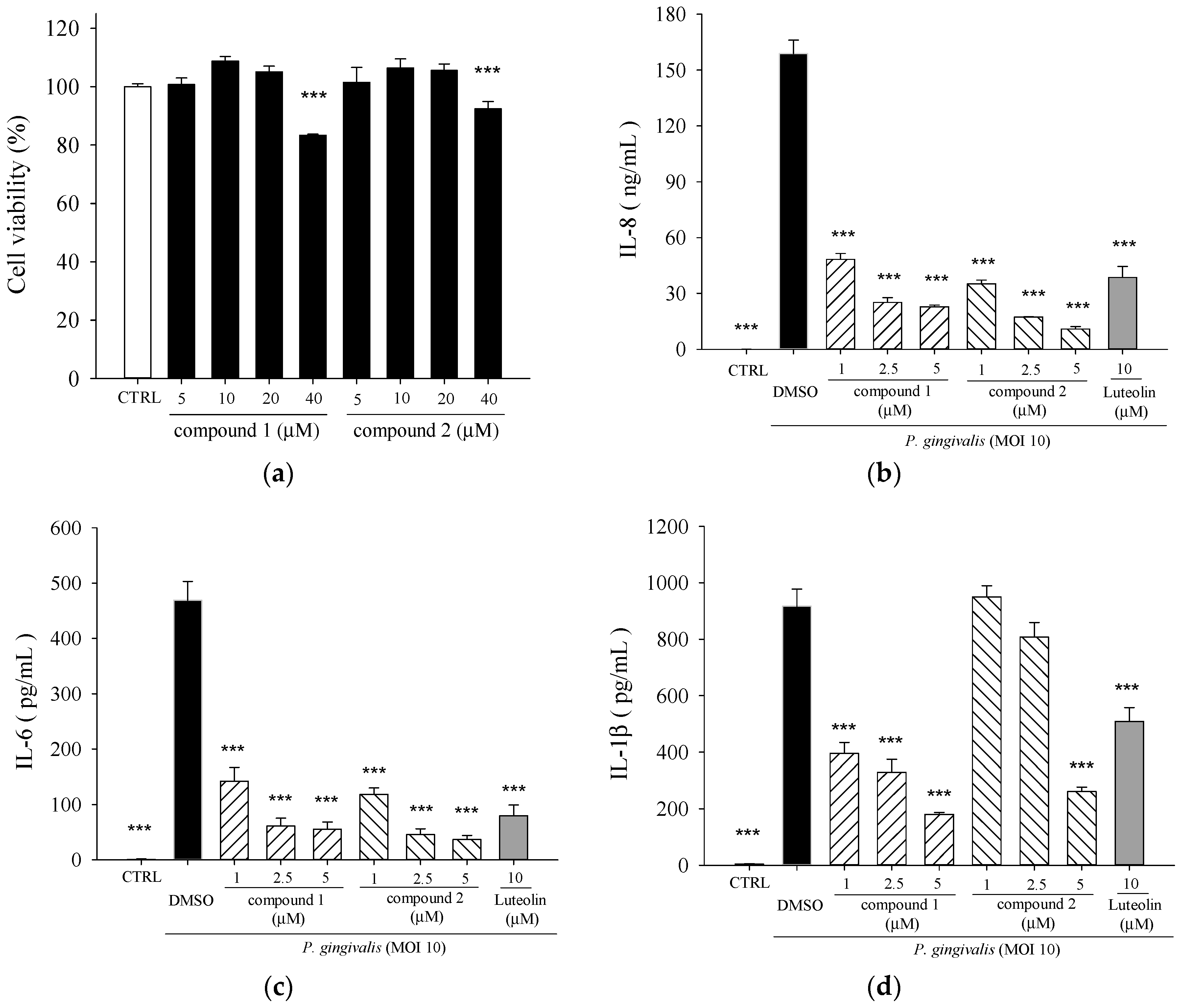
2.2. Effects of 5β,19-Epoxycucurbita-6,23-diene-3β,19,25-triol (1) and 3β,7β,25-Trihydroxycucurbita-5,23-dien-19-al (2) on P. gingivalis-Induced Cytokines in THP-Cells
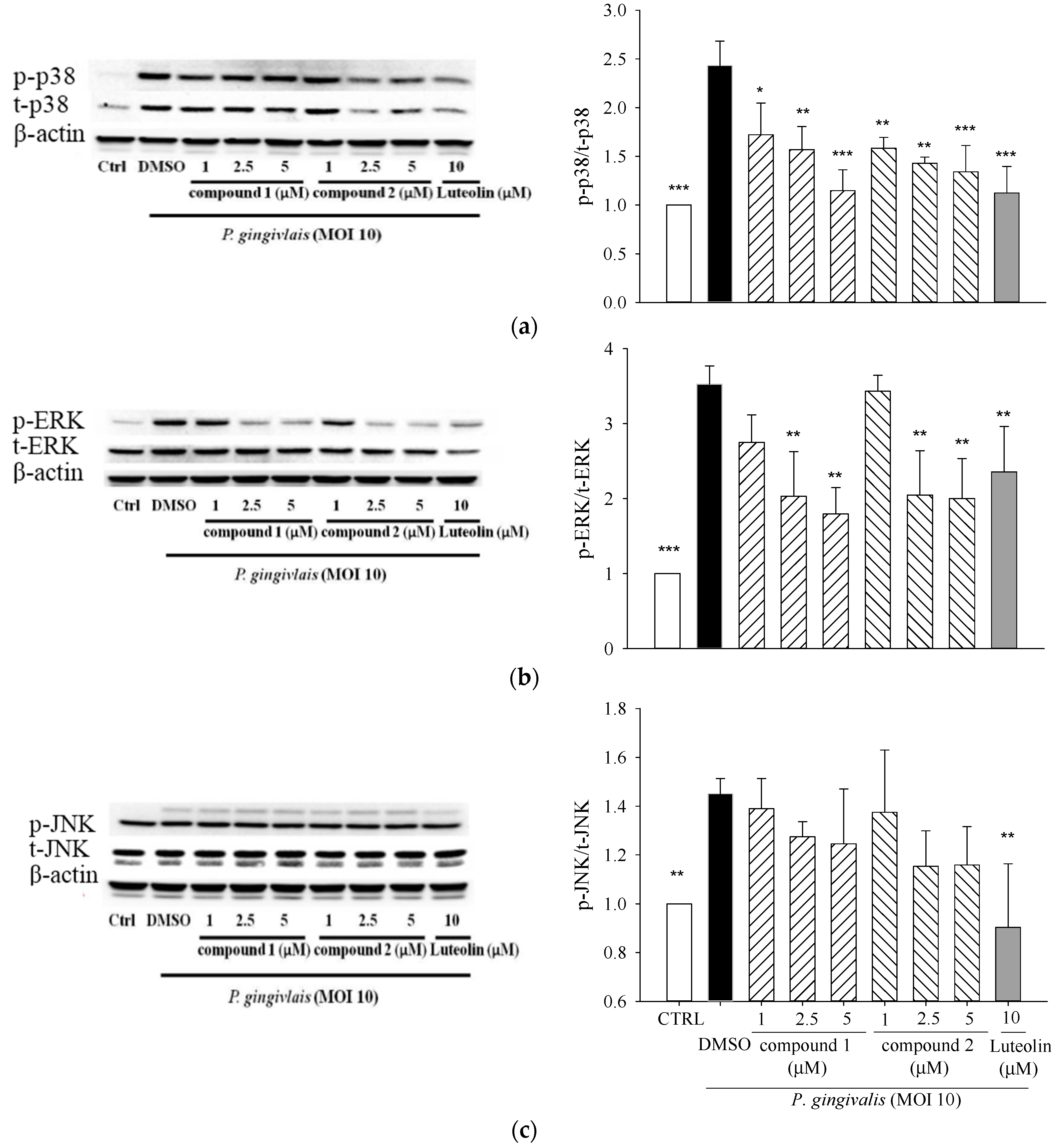
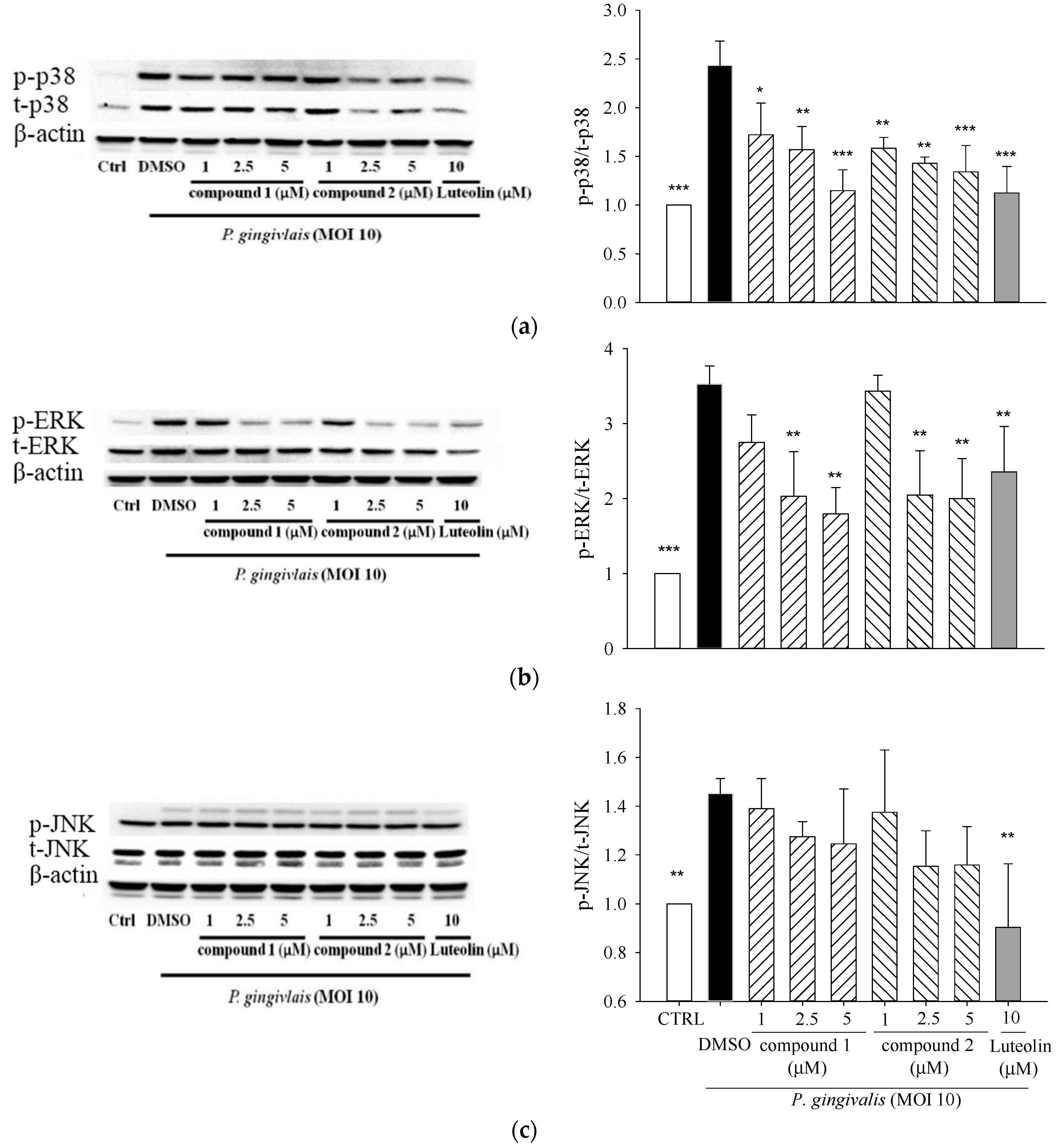
2.3. Effects of 5β,19-Epoxycucurbita-6,23-diene-3β,19,25-triol (1) and 3β,7β,25-Trihydroxycucurbita-5,23-dien-19-al (2) on MAPK Activation in P. gingivalis-Stimulated THP-1 Cells
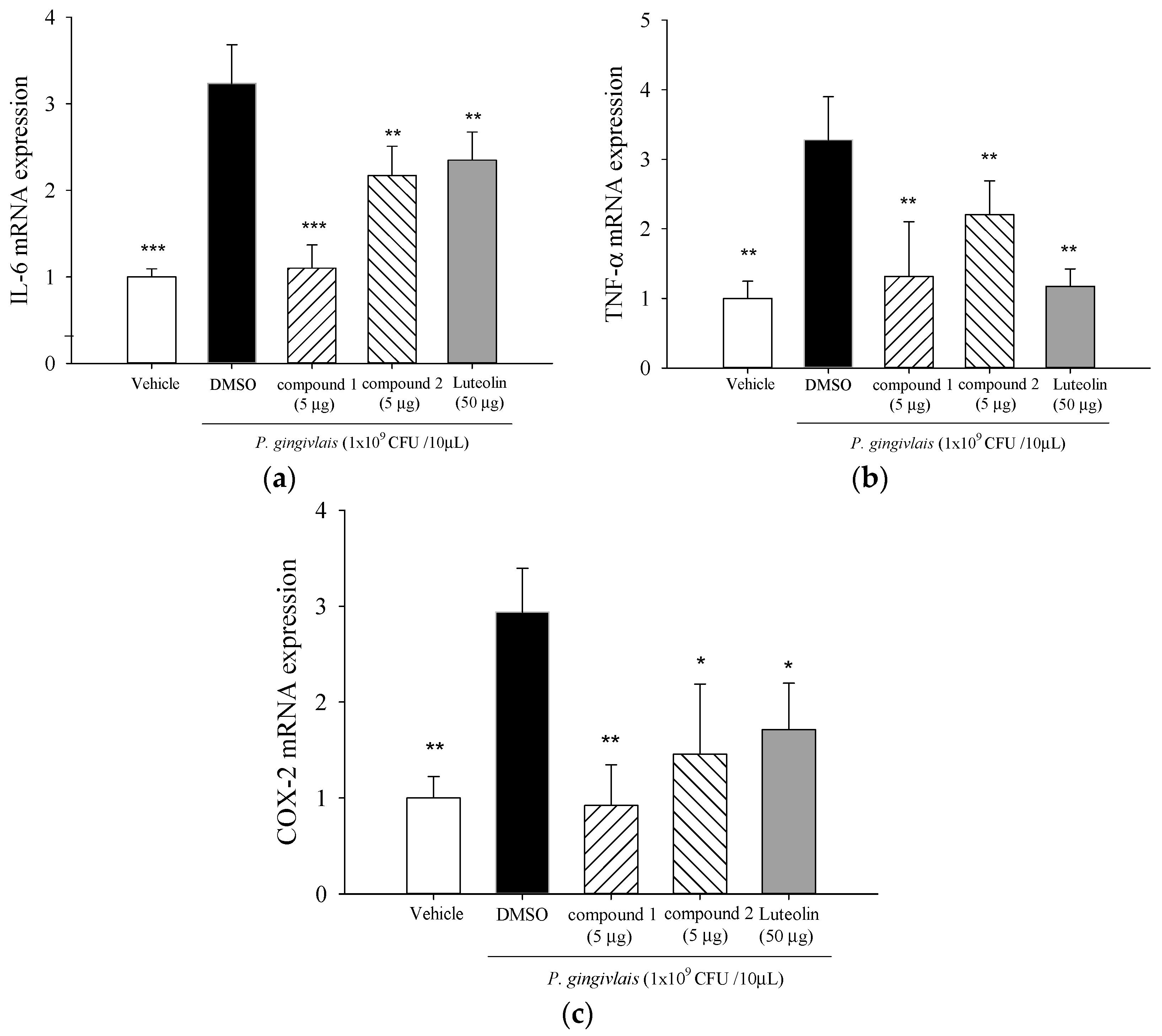
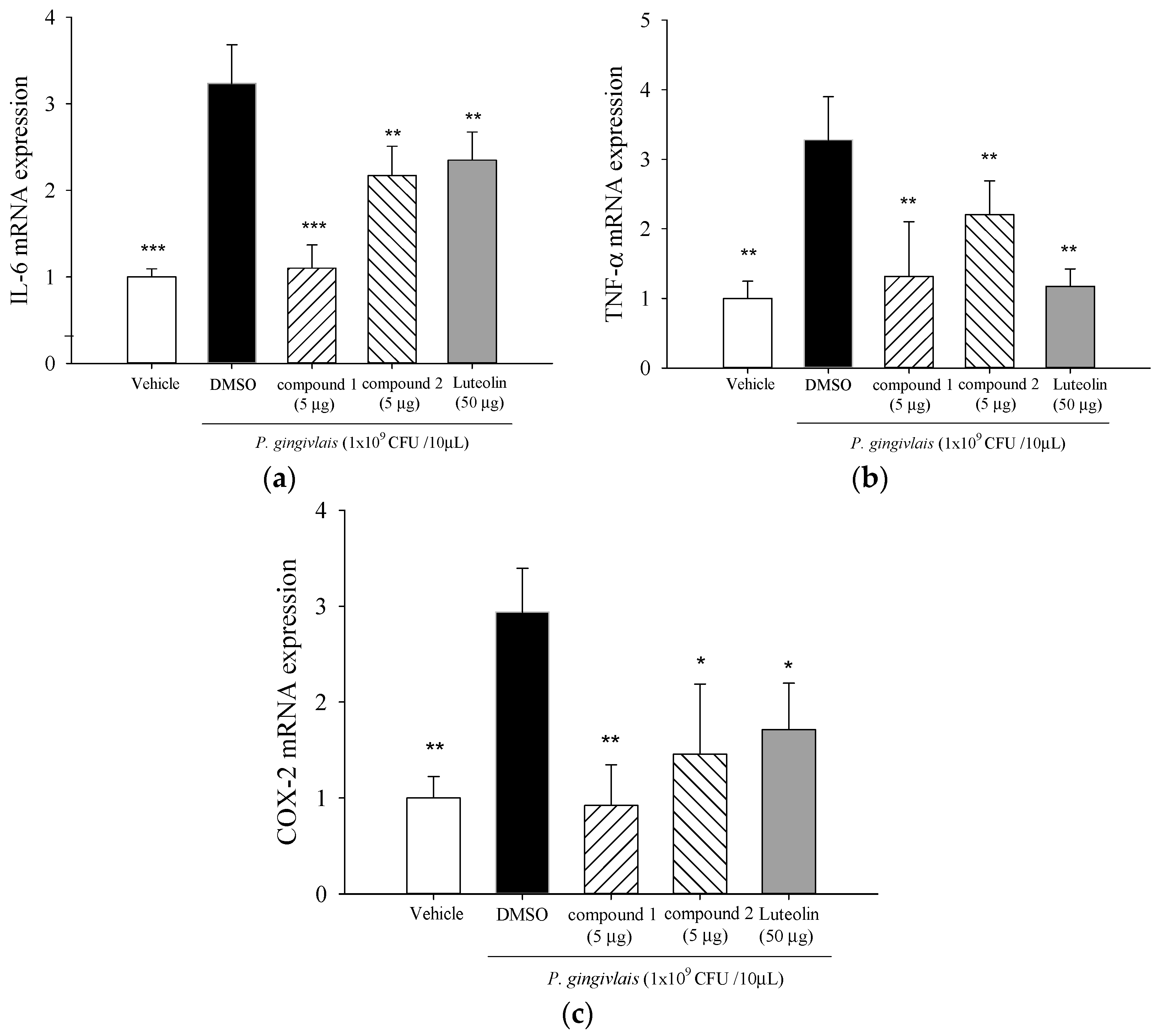
2.4. Effect of 5β,19-Epoxycucurbita-6,23-diene-3β,19,25-triol (1) and 3β,7β,25-Trihydroxycucurbita-5,23-dien-19-al (2) on mRNA Levels of IL-6, TNF-α, and cyclooxygenase (COX)-2 in P. gingivalis-Stimulated Gingival Tissue of Mice
3. Discussion
4. Materials and Methods
4.1. Plant Material
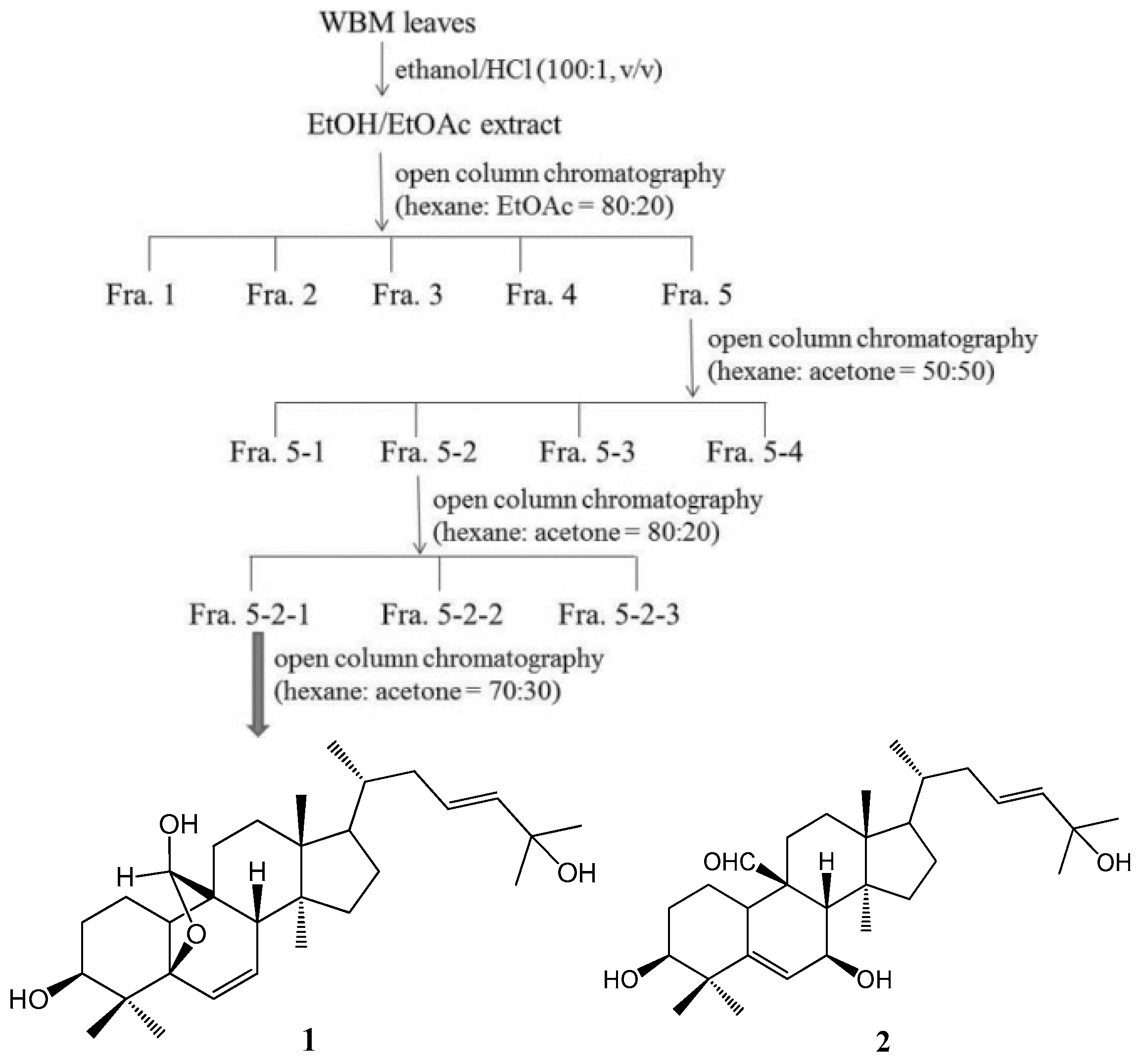
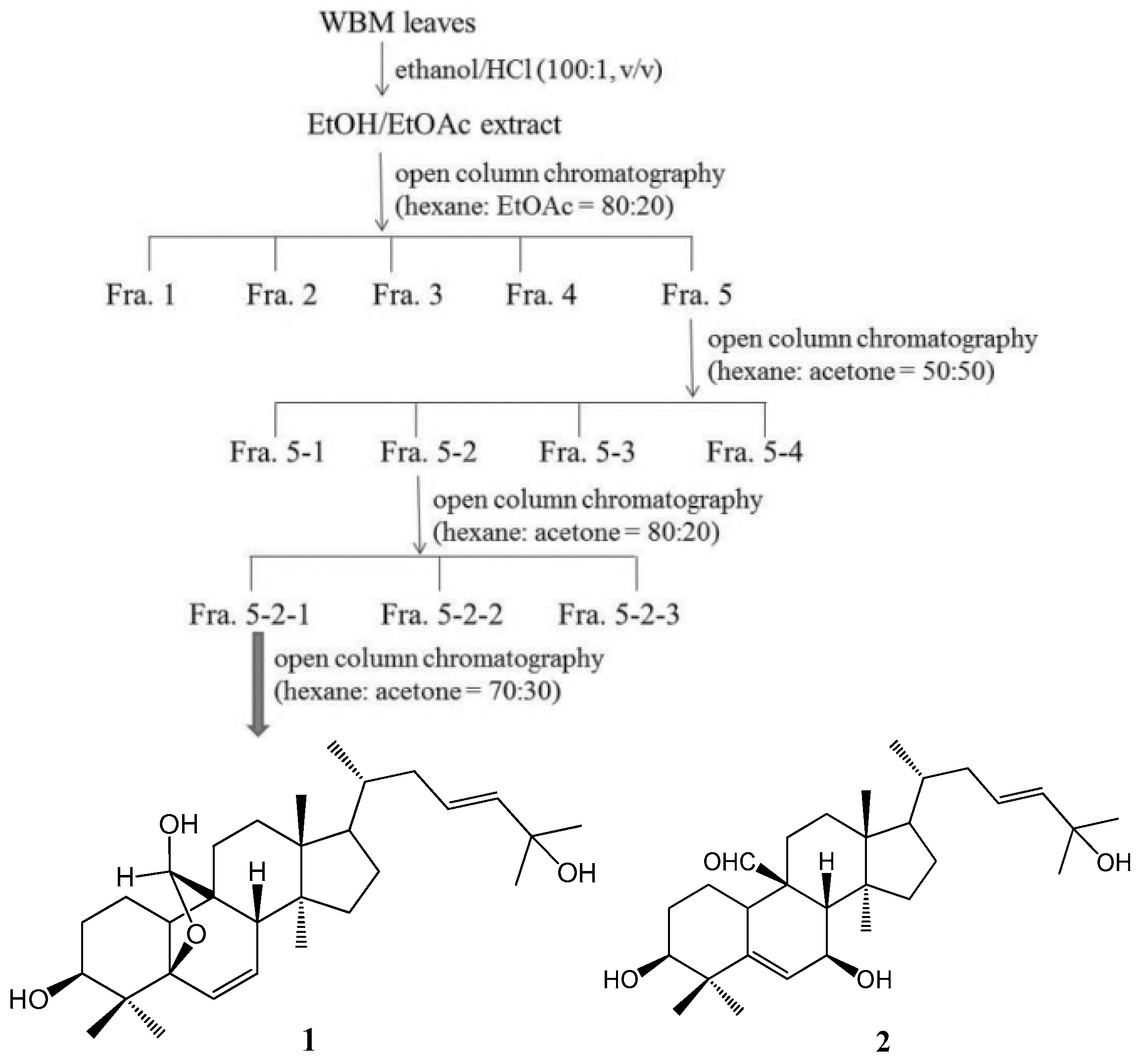
4.2. Isolation and Determination of the Active Compounds
4.3. Preparation of Heat-Inactivated Porphyromonas Gingivalis
4.4. Cell Culture and Determination of the Viability of THP-1 Cells
4.5. Measurement of Cytokine Production in Human Monocytic THP-1 Cells
4.6. Detection of MAPK Phosphorylation by Western Blot Analysis
4.7. Induction of Experimental Periodontitis by Injection of P. gingivalis
4.8. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (PCR)
4.9. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Sojar, H.; Genco, R.J.; DeNardin, E. Intracellular signaling and cytokine induction upon interactions of Porphyromonas gingivalis fimbriae with pattern-recognition receptors. Immunol. Investig. 2004, 33, 157–172. [Google Scholar] [CrossRef]
- Hiramine, H.; Watanabe, K.; Hamada, N.; Umemoto, T. Porphyromonas gingivalis 67-kDa fimbriae induced cytokine production and osteoclast differentiation utilizing TLR2. FEMS Microbiol. Lett. 2003, 229, 49–55. [Google Scholar] [CrossRef]
- Imatani, T.; Kato, T.; Okuda, K. Production of inflammatory cytokines by human gingival fibroblasts stimulated by cell-surface preparations of Porphyromonas gingivalis. Oral Microbiol. Immunol. 2001, 16, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Hagiwara, M.; Ishihara, Y.; Isoda, R.; Sugiura, S.; Komatsu, T.; Ishida, N.; Noguchi, T.; Matsushita, K. TNF-α augmented Porphyromonas gingivalis invasion in human gingival epithelial cells through Rab5 and ICAM-1. BMC Microbiol. 2014, 14, 229. [Google Scholar] [CrossRef] [PubMed]
- Mysak, J.; Podzimek, S.; Sommerova, P.; Lyuya-Mi, Y.; Bartova, J.; Janatova, T.; Prochazkova, J.; Duskova, J. Porphyromonas gingivalis: Major periodontopathic pathogen overview. J. Immunol. Res. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Birkedal-Hansen, H. Role of cytokines and inflammatory mediators in tissue destruction. J. Periodontal. Res. 1993, 28, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Bickel, M. The role of interleukin-8 in inflammation and mechanisms of regulation. J. Periodontol. 1993, 64 (Suppl. S5), 456–460. [Google Scholar] [PubMed]
- Okada, H.; Murakami, S. Cytokine expression in periodontal health and disease. Crit. Rev. Oral Biol. Med. 1998, 9, 248–266. [Google Scholar] [CrossRef] [PubMed]
- Azuma, M.M.; Samuel, R.O.; Gomes-Filho, J.E.; Dezan-Junior, E.; Cintra, L.T.A. The role of IL-6 on apical periodontitis: A systematic review. Int. Endod. J. 2014, 47, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, Z.; Goultschin, J.; Dean, D.D.; Boyan, B.D. Mechanisms of alveolar bone destruction in periodontitis. Periodontol. 2000 1997, 14, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Rossomando, E.F.; Kennedy, J.E.; Hadjimichael, J. Tumour necrosis factor alpha in gingival crevicular fluid as a possible indicator of periodontal disease in humans. Arch. Oral Biol. 1990, 35, 431–434. [Google Scholar] [CrossRef]
- Kato, H.; Taguchi, Y.; Tominaga, K.; Umeda, M.; Tanaka, A. Porphyromonas gingivalis LPS inhibits osteoblastic differentiation and promotes pro-inflammatory cytokine production in human periodontal ligament stem cells. Arch. Oral Biol. 2014, 59, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Noh, M.K.; Jung, M.; Kim, S.H.; Lee, S.; Park, K.H.; Kim, D.H.; Kim, H.H.; Park, Y.G. Assessment of IL-6, IL-8 and TNF-α levels in the gingival tissue of patients with periodontitis. Exp. Ther. Med. 2013, 6, 847–851. [Google Scholar] [PubMed]
- Andrukhov, O.; Ulm, C.; Reischl, H.; Nguyen, P.Q.; Matejka, M.; Rausch-Fan, X. Serum cytokine levels in periodontitis patients in relation to the bacterial load. J. Periodontaol. 2011, 82, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Saengsai, J.; Kongtunjanphuk, S.; Yoswatthana, N.; Kummalue, T.; Jiratchariyakul, W. Antibacterial and antiproliferative activities of plumericin, an iridoid Isolated from Momordica charantia vine. Evid. Based Complement. Altern. Med. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Kubola, J.; Siriamornpun, S. Phenolic contents and antioxidant activities of bitter gourd (Momordica charantia L.) leaf, stem and fruit fraction extracts in vitro. Food Chem. 2008, 110, 881–890. [Google Scholar] [PubMed]
- Nagarani, G.; Abirami, A.; Siddhuraju, P. Food prospects and nutraceutical attributes of Momordica species: A potential tropical bioresources—A review. Food Sci. Hum. Wellness 2014, 3, 36–46. [Google Scholar] [CrossRef]
- Tsai, T.H.; Huang, C.J.; Wu, W.H.; Huang, W.C.; Chyuan, J.H.; Tsai, P.J. Antioxidant, cell-protective, and anti-melanogenic activities of leaf extracts from wild bitter melon (Momordica charantia Linn. var. abbreviata Ser.) cultivars. Bot. Stud. 2014, 55, 78–88. [Google Scholar] [CrossRef]
- Huang, W.C.; Tsai, T.H.; Huang, C.J.; Li, Y.Y.; Chyuan, J.H.; Chuang, L.T.; Tsai, P.J. Inhibitory effects of wild bitter melon leaf extract on Propionibacterium acnes-induced skin inflammation in mouse and cytokine production in vitro. Food Funct. 2015, 6, 2550–2560. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C. Bioassay-Guided Isolation and Identification of Anti-Inflammatory Compounds from Wild Bitter Melon Leaf against Propionibacterium acnes. Ph.D. Thesis, National Taiwan Normal University, Taipei, Taiwan, 2016. [Google Scholar]
- Petrovic, M.S.; Kesic, L.G.; Kitic, D.V.; Milašin, J.M.; Obradovic, R.R.; Bojovic, M.D.; Simonovic, A.A. Periodontal disease and phytotherapy. Org. Hyg. Health 2015, 3, 172–175. [Google Scholar]
- Jagan Rao, N.; Subash, K.R.; Sandeep Kumar, K. Role of phytotherapy in gingivitis: A review. Int. J. Pharmacol. 2012, 8, 1–5. [Google Scholar] [CrossRef]
- Lii, C.K.; Chen, H.W.; Yun, W.T.; Liu, K. Suppressive effects of wild bitter gourd (Momordica charantia Linn. var. abbreviate ser.) fruit extracts on inflammatory responses in RAW 264.7 macrophages. J. Ethnopharmacol. 2009, 122, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.; Tsai, T.H.; Li, Y.Y.; Wu, W.H.; Huang, C.J.; Tsai, P.J. Wild bitter melon (Momordica charantia Linn. var. abbreviata Ser.) extract and its bioactive components suppress Propionibacterium acnes-induced inflammation. Food Chem. 2012, 135, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Ciou, S.Y.; Hsu, C.C.; Kuo, Y.H.; Chao, C.Y. Effect of wild bitter gourd treatment on inflammatory responses in BALB/c mice with sepsis. Biomedicine 2014, 4, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Ohara-Nemoto, Y.; Shimoyama, Y.; Ishikawa, T.; Sasaki, M. Pathogenic factors of P. gingivalis and the host defense mechanisms. In Pathogenesis and Treatment of Periodontitis; Buduneli, N., Ed.; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Lim, T.K. Momordica charantia. In Edible Medicinal and Non-Medicinal Plants; Lim, T.K., Ed.; Springer: New York, NY, USA, 2012; Volume 2, pp. 331–368. [Google Scholar]
- Hsu, C.; Hsieh, C.L.; Kuo, Y.H.; Huang, C.J. Isolation and identification of cucurbitane-type triterpenoids with partial agonist/antagonist potential for estrogen receptors from Momordica charantia. J. Agric. Food Chem. 2011, 59, 4553–4561. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.R.; Liao, Y.W.; Wang, L.; Kuo, Y.H.; Liu, H.J.; Shih, W.L.; Cheng, H.L.; Chang, C.I. Cucurbitane triterpenoids from Momordica charantia and their cytoprotective activity in tert-butyl hydroperoxide-induced hepatotoxicity of HepG2 cells. Chem. Pharm. Bull. 2010, 58, 1639–1642. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.R.; Bai, L.Y.; Chiu, C.F.; Hu, J.L.; Chiu, S.J.; Wu, C.Y. Cucurbitane triterpenoid from Momordica charantia induces apoptosis and autophagy in breast cancer cells, in part, through peroxisome proliferator-activated receptor 𝛾 activation. Evid. Based Complement. Altern. Med. 2013, 2013, 935675. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.L.; Huang, H.K.; Chang, C.I.; Tsai, C.P.; Chou, C.H. A cell-based screening identifies compounds from the stem of Momordica charantia that overcome insulin resistance and activate AMP-activated protein kinase. J. Agric. Food Chem. 2008, 56, 6835–6843. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, Y.; Kikuchi, T.; Tokuda, H.; Suzuki, N.; Inafuku, K.; Miura, M.; Motohashi, S.; Suzuki, T.; Akihisa, T. Cucurbitane triterpenoids from the leaves of Momordica charantia, and their cancer chemopreventive effects and cytotoxicities. Chem. Biodivers. 2012, 9, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Harinantenaina, L.; Tanaka, M.; Takaoka, S.; Oda, M.; Mogami, O.; Uchida, M.; Asakawa, Y. Momordica charantia constituents and antidiabetic screening of the isolated major compounds. Chem. Pharm. Bull. 2006, 54, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.T.; Liu, J.Q.; Deng, Y.Y.; Li, H.Z.; Chen, J.C.; Zhang, Z.R.; Zhou, L.; Qiu, M.H. Cucurbitane-type triterpenoids from the stems and leaves of Momordica charantia. Fitoterapia 2014, 95, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Z.B.; Tang, C.H.; Chen, G.; Shi, Y.S. Study on colorimetric determination of oleanolic acid in Chinese quince. Nat. Prod. Res. Dev. 2001, 13, 23–26. [Google Scholar]
- Zhou, Q.; Amar, S. Identification of signaling pathways in macrophage exposed to Porphyromonas gingivalis or to its purified cell wall components. J. Immunol. 2007, 179, 7777–7790. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Y.; Ouyang, X. Beyond toll-like receptors: Porphyromonas gingivalis induces IL-6, IL-8, and VCAM-1 expression through NOD-mediated NF-κB and ERK signaling pathways in periodontal fibroblasts. Inflammation 2014, 37, 522–533. [Google Scholar] [CrossRef] [PubMed]
- Newton, K.; Dixit, V.M. Signaling in Innate Immunity and Inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef] [PubMed]
- Hienz, S.A.; Paliwal, S.; Ivanoki, S. Mechanisms of bone resorption in periodontitis. J. Immunol. Res. 2015, 2015, 615486. [Google Scholar] [CrossRef] [PubMed]
- Cochran, D.L. Inflammation and bone loss in periodontal disease. J. Periodontol. 2008, 79, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Mesa, F.; Aguilar, M.; Galindo-Moreno, P.; Bravo, M.; O’Valle, F. Cyclooxygenase-2 expression in gingival biopsies from periodontal patients is correlated with connective tissue loss. J. Periodontol. 2012, 83, 1538–1545. [Google Scholar] [CrossRef] [PubMed]
- Younis, M.N.; Machtei, E.E.; Oetlinger-Barak, O.; Peled, M. The significance of gingival crevicular fluid and IL-1β, TNF-α and PGE2 in periodontal disease: A literature review. Oral Biosci. Med. 2004, 1, 249–257. [Google Scholar]
- Fatope, M.O.; Takeda, Y.; Yamashita, H.; Okabe, H.; Yamauchi, T. New cucurbitane triterpenoids from Momordica charantia. J. Nat. Prod. 1990, 53, 1491–1497. [Google Scholar] [CrossRef]
- Mulholland, D.A.; Sewram, V.; Osborne, R.; Pegel, K.H.; Connolly, J.D. Cucurbitane triterpenoids from the leaves of Momordica foetida. Phytochemistry 1997, 45, 391–395. [Google Scholar] [CrossRef]
- Baqui, A.A.M.A.; Meiller, T.F.; Falkler, W.A. Enhanced interleukin-8 production in THP-1 human monocytic cells by lipopolysaccharide from oral microorganisms and granulocyte-macrophage colony-stimulating factor. Oral Microbiol. Immunol. 1999, 14, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Chen, L.; Li, S.; Gu, Z.; Yan, J. Lipopolysaccharide (LPS) of Porphyromonas gingivalis induces IL-1β, TNF-α and IL-6 production by THP-1 cells in a way different from that of Escherichia coli LPS. Innate Immun. 2008, 14, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Gutie´rrez-Venegas, G.; Contreras-Sa´nchez, A. Luteolin and fisetin inhibit the effects of lipopolysaccharide obtained from Porphyromonas gingivalis in human gingival fibroblasts. Mol. Biol. Rep. 2013, 40, 477–485. [Google Scholar] [CrossRef] [PubMed]
- De Molon, R.S.; de Avila, E.D.; Boas Nogueira, A.V.; Chaves de Souza, J.A.; Avila-Campos, M.J.; de Andrade, C.R.; Cirelli, J.A. Evaluation of the host response in various models of induced periodontal disease in mice. J. Periodontol. 2014, 85, 465–477. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds, 5β,19-epoxycucurbita-6,23-diene-3β,19,25-triol and 3β,7β,25-trihydroxycucurbita-5,23-dien-19-al are not available from the authors.

© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, T.-H.; Huang, W.-C.; Ying, H.-T.; Kuo, Y.-H.; Shen, C.-C.; Lin, Y.-K.; Tsai, P.-J. Wild Bitter Melon Leaf Extract Inhibits Porphyromonas gingivalis-Induced Inflammation: Identification of Active Compounds through Bioassay-Guided Isolation. Molecules 2016, 21, 454. https://doi.org/10.3390/molecules21040454
Tsai T-H, Huang W-C, Ying H-T, Kuo Y-H, Shen C-C, Lin Y-K, Tsai P-J. Wild Bitter Melon Leaf Extract Inhibits Porphyromonas gingivalis-Induced Inflammation: Identification of Active Compounds through Bioassay-Guided Isolation. Molecules. 2016; 21(4):454. https://doi.org/10.3390/molecules21040454
Chicago/Turabian StyleTsai, Tzung-Hsun, Wen-Cheng Huang, How-Ting Ying, Yueh-Hsiung Kuo, Chien-Chang Shen, Yin-Ku Lin, and Po-Jung Tsai. 2016. "Wild Bitter Melon Leaf Extract Inhibits Porphyromonas gingivalis-Induced Inflammation: Identification of Active Compounds through Bioassay-Guided Isolation" Molecules 21, no. 4: 454. https://doi.org/10.3390/molecules21040454