
Comparison of Anti-Oxidant and Anti-Inflammatory Effects between Fresh and Aged Black Garlic Extracts

 ,
, 
Abstract
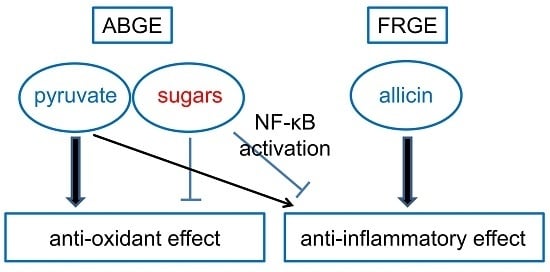
:
1. Introduction
2. Results
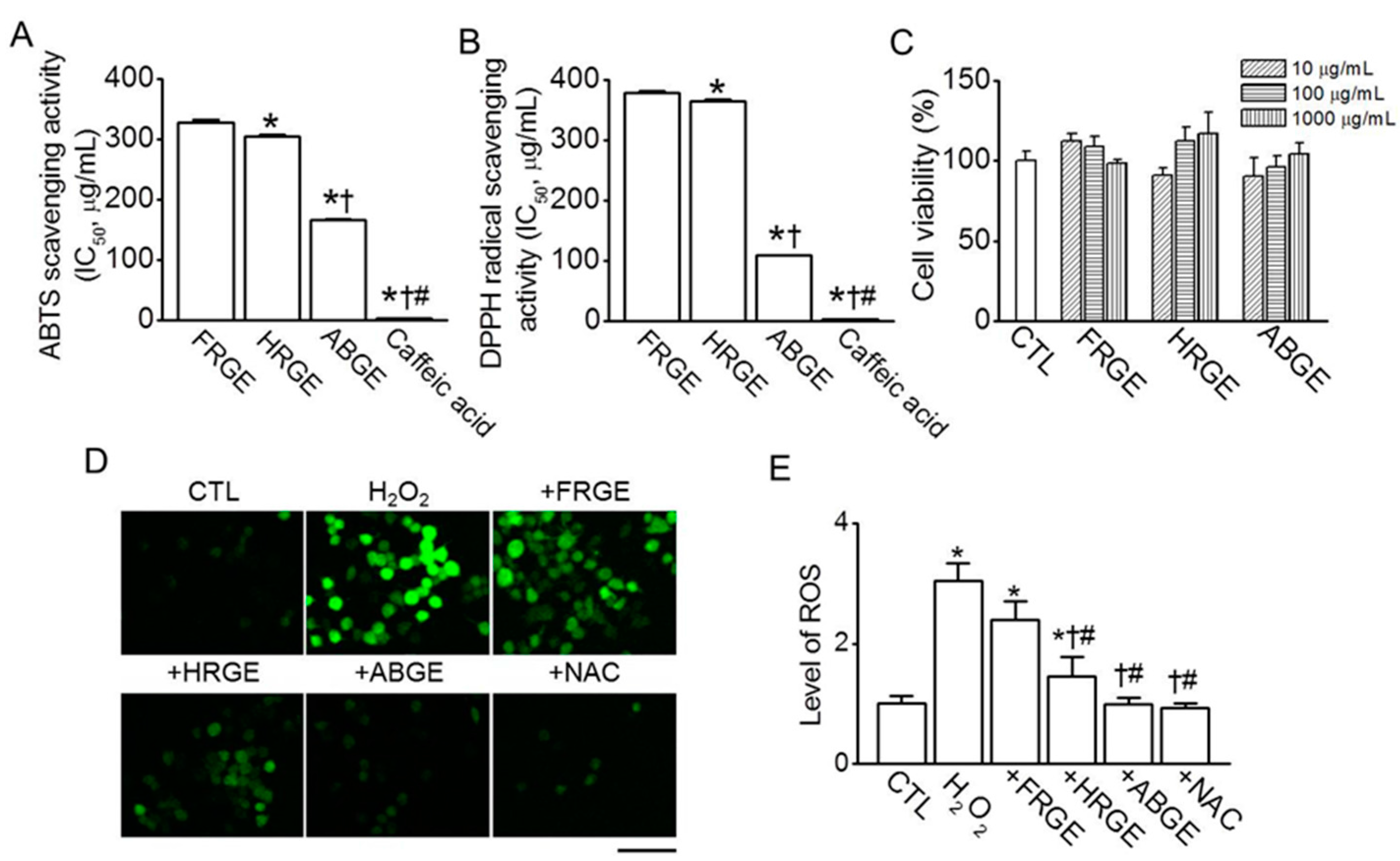
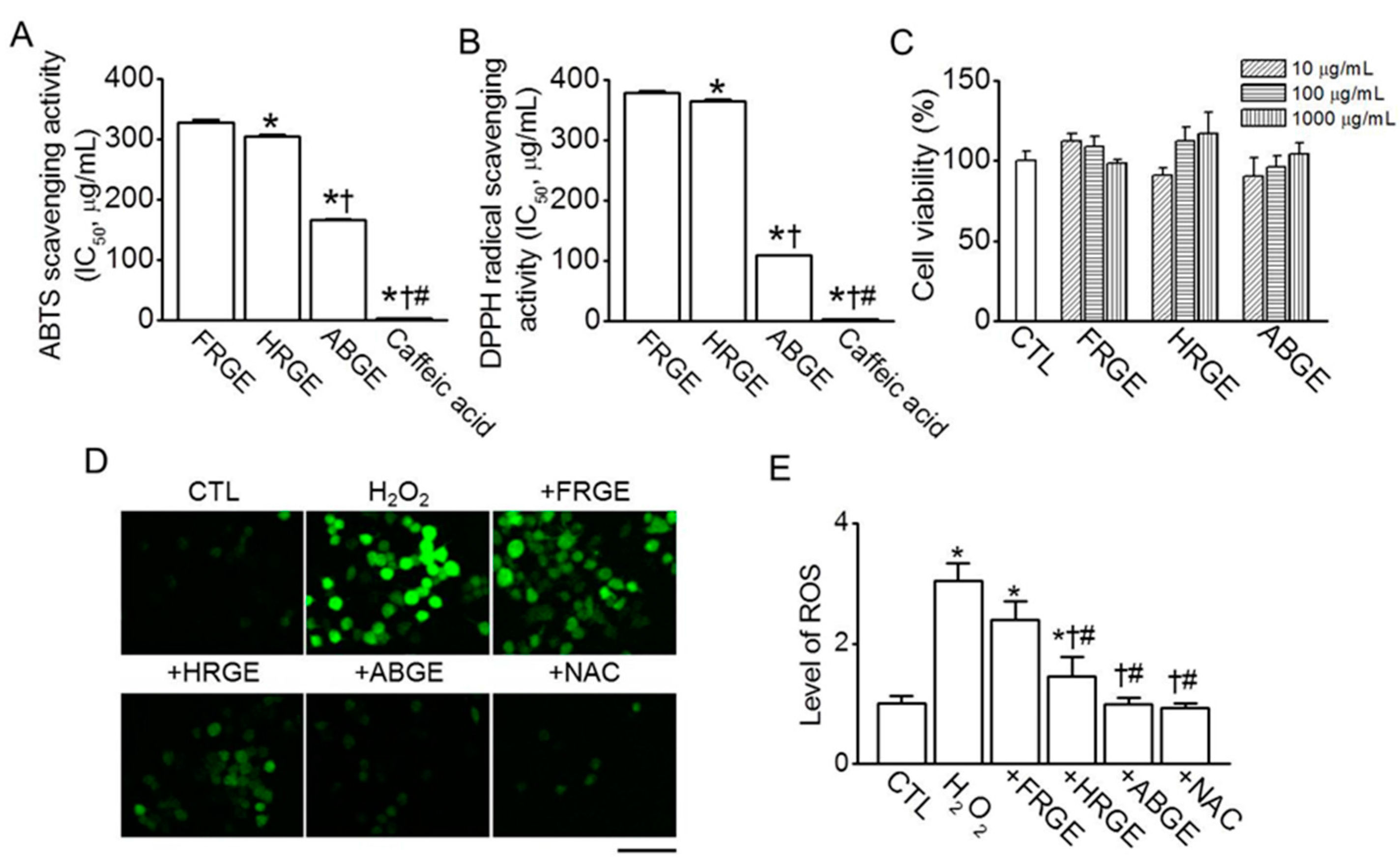
2.1. High Anti-Oxidant Activity in ABGE
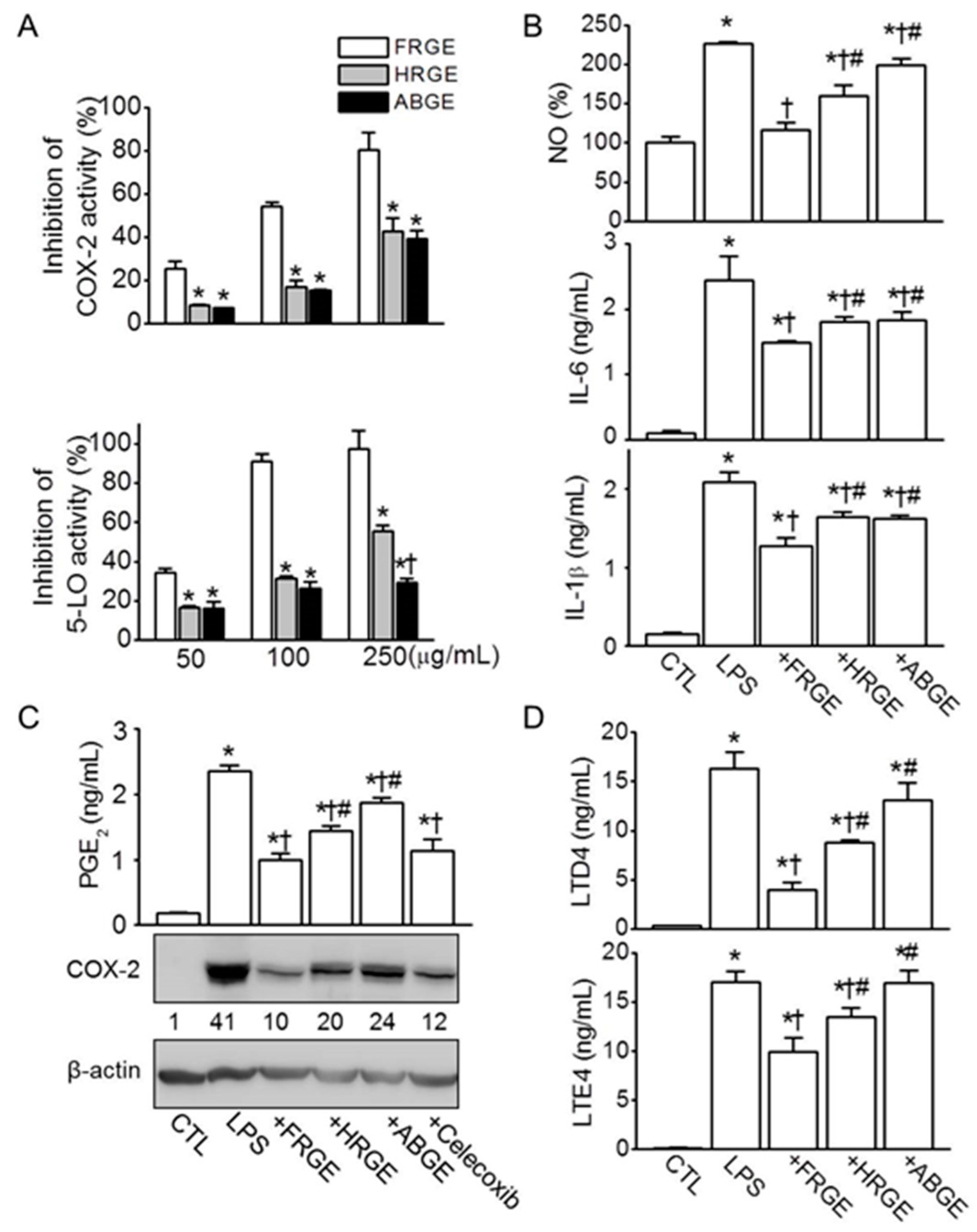
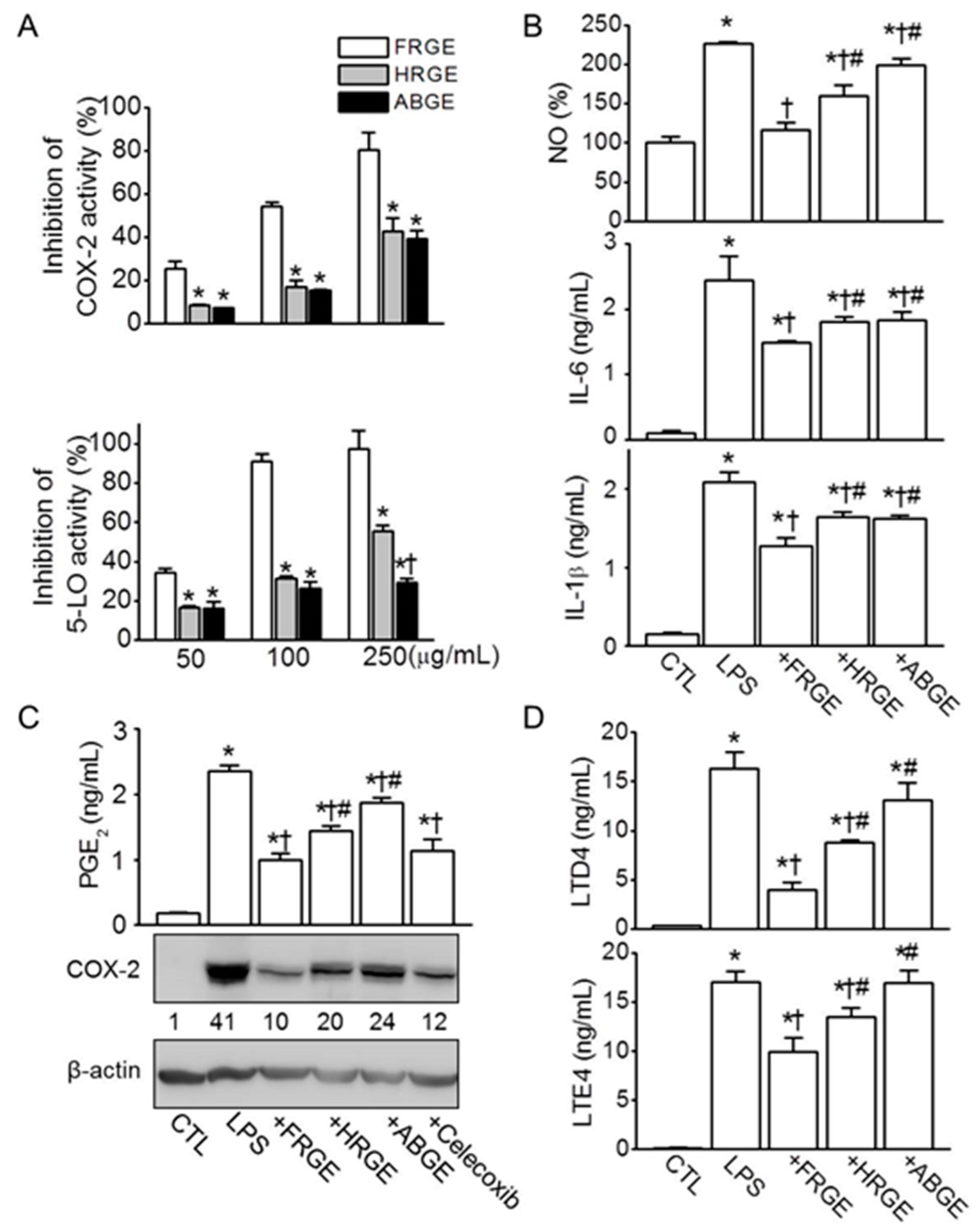
2.2. Low Anti-Inflammatory Activity in ABGE
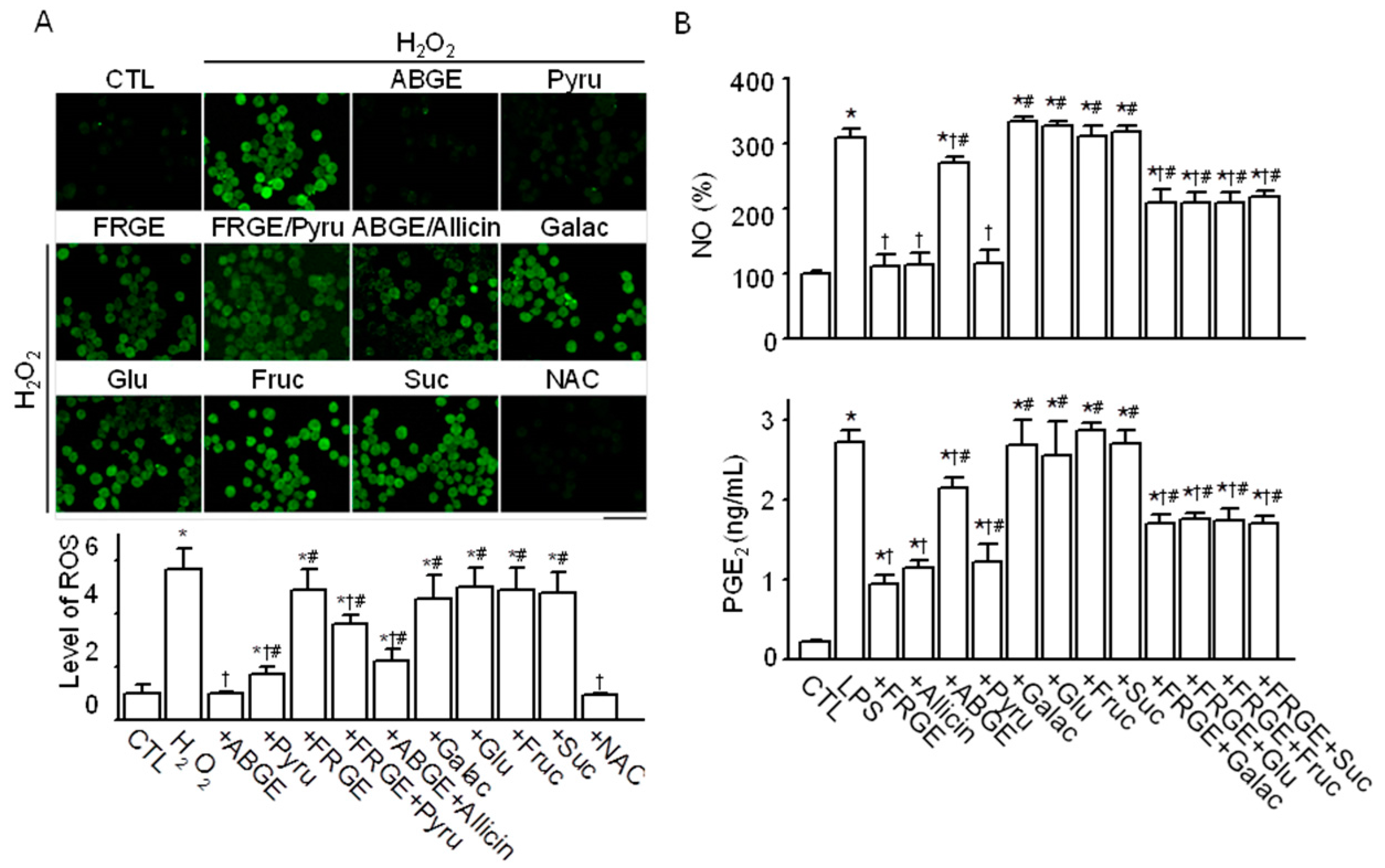
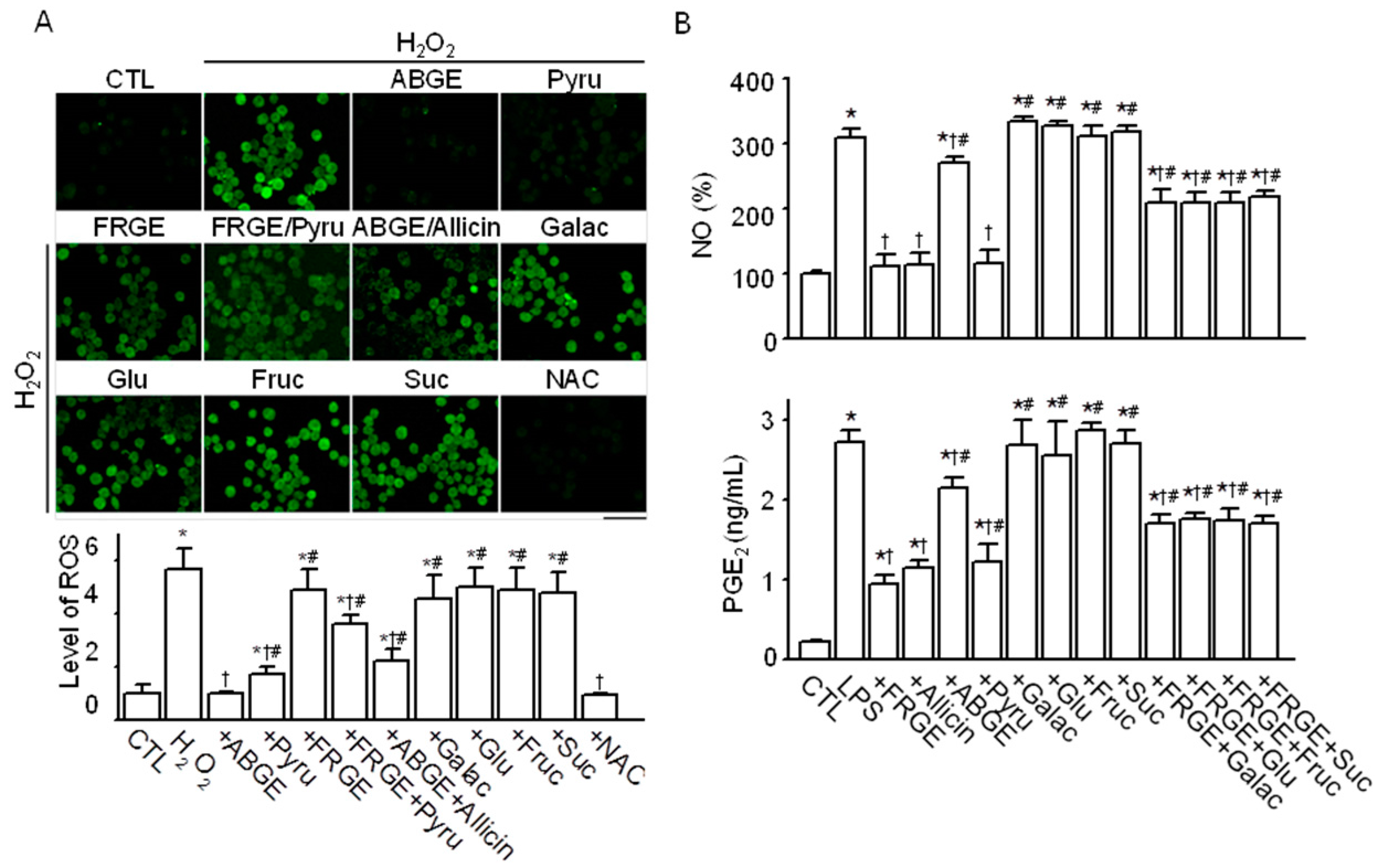
2.3. Pyruvate is Responsible for the High Anti-Oxidant Effects of ABGE
2.4. Sugars are Responsible for the Low Anti-Inflammatory Effect of ABGE
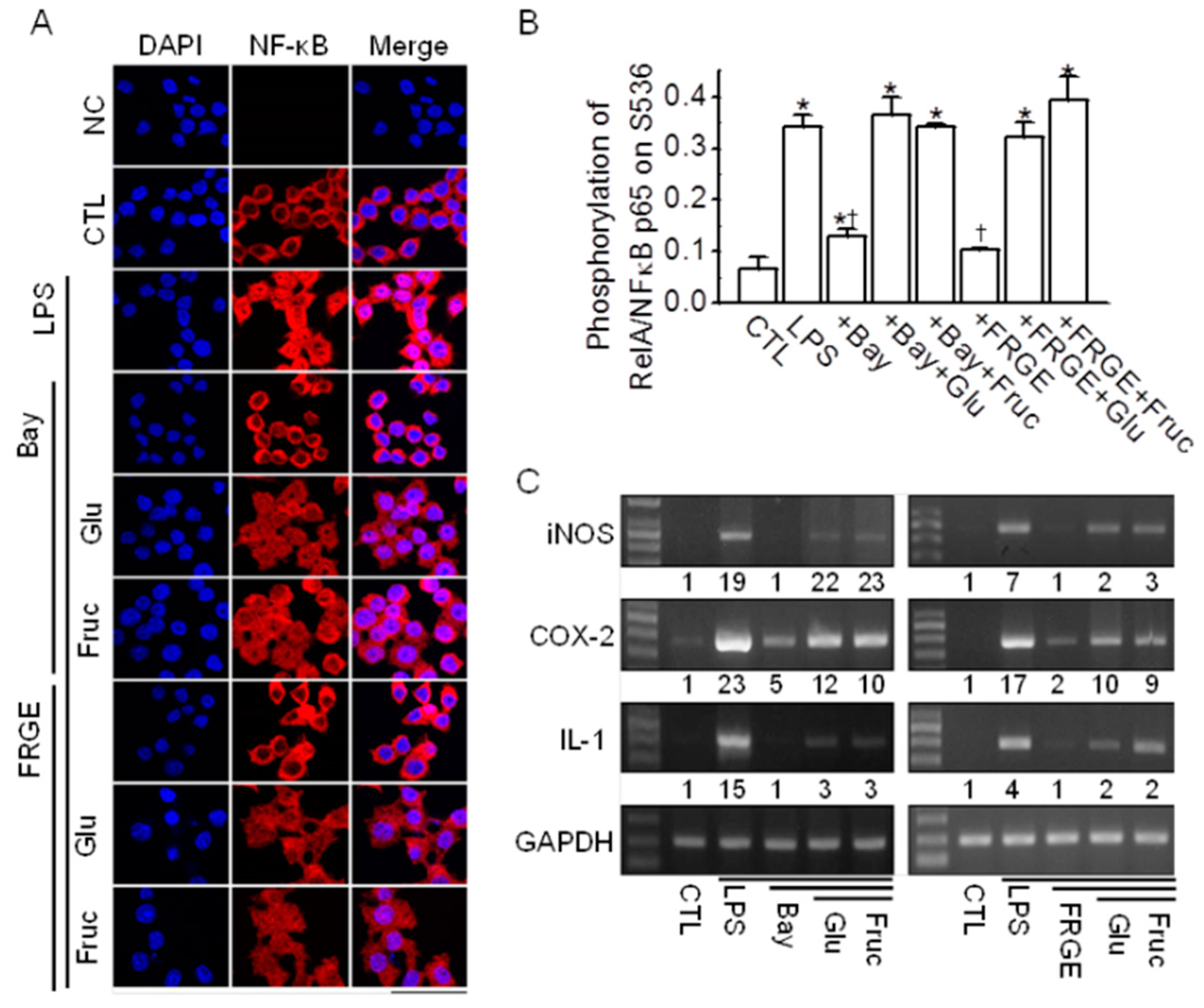
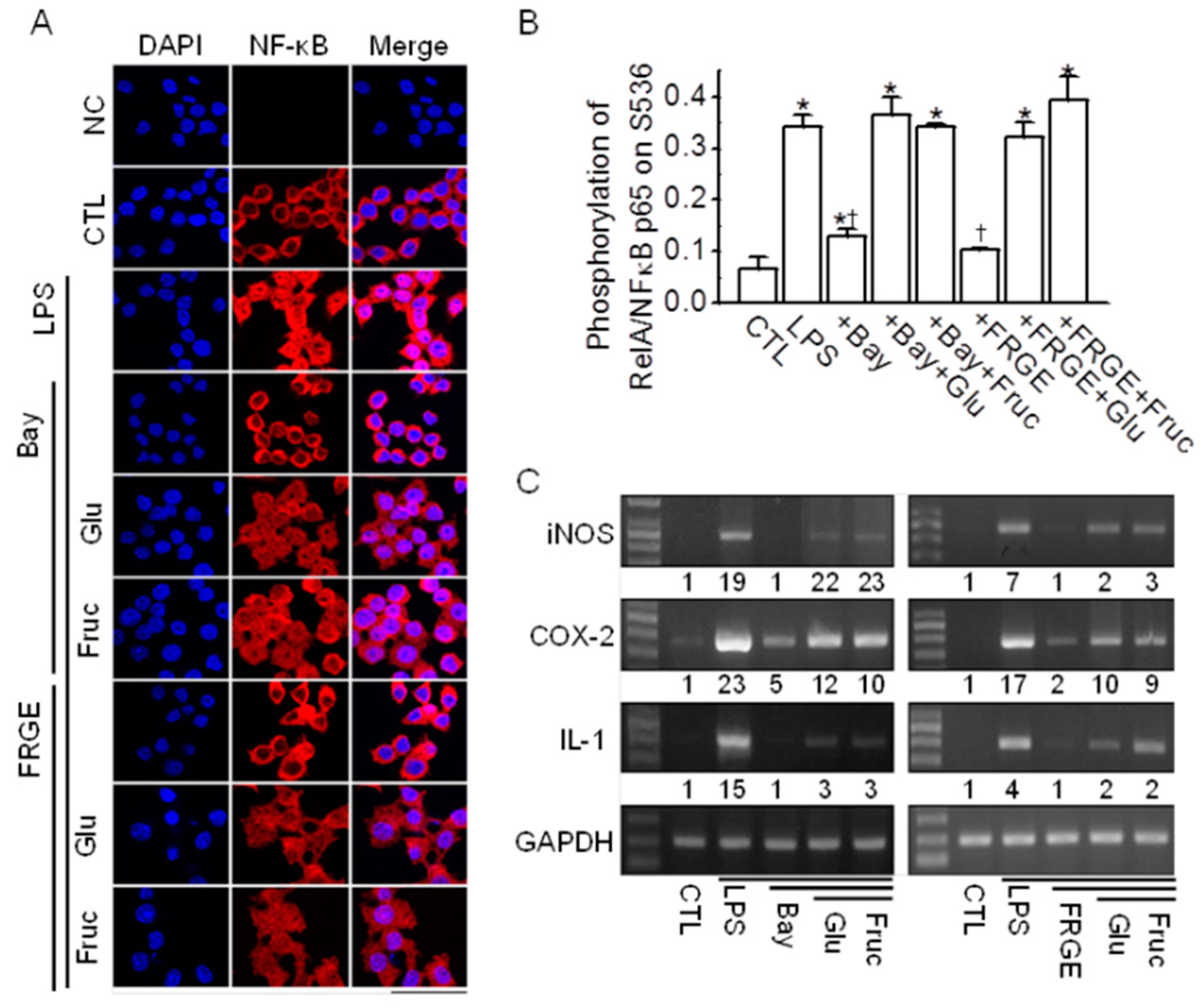
2.5. Sugars Induce NF-κB Activation
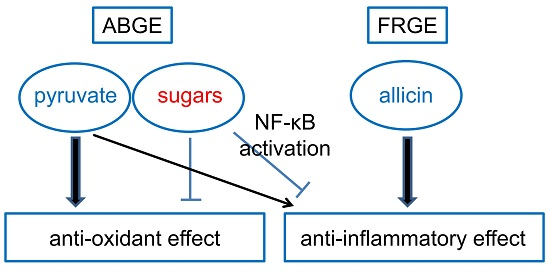
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Reagents
4.2. Preparation of Water-Soluble Garlic Extracts
4.3. Measurement of ABTS Radical Scavenging Activity
4.4. Measurement of DPPH Radical Scavenging Activity
4.5. Measurement of Cyclooxygenase-2 (COX-2)
4.6. Measurement of 5-Lipoxygenase (5-LO)
4.7. Measurement of Allicin, Free Sugar, Pyruvate, Thiosulfate, and SAC
4.8. Cell Viability Assay
4.9. Measurement of ROS Generation
4.10. Measurement of Nitrite Levels
4.11. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.12. Western Blot Analysis
4.13. Immunocytochemistry
4.14. Enzyme-Linked Immunosorbent Assays (ELISA) for Pro-Inflammatory Cytokines and NF-κB
4.15. Data Analysis and Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ABGE | Aged black garlic extract |
| EGCG | Epigallocatechin gallate |
| FRGE | Fresh raw garlic extracts |
| HRGE | Heated raw garlic extract |
| NAC | N-acetyl-L-cysteine |
References
- Freeman, B.A.; Crapo, J.D. Biology of disease: Free radicals and tissue injury. Lab. Investig. 1982, 47, 412–426. [Google Scholar] [PubMed]
- Berlett, B.S.; Stadtman, E.R. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [PubMed]
- Salzano, S.; Checconi, P.; Hanschmann, E.M.; Lillig, C.H.; Bowler, L.D.; Chan, P.; Vaudry, D.; Mengozzi, M.; Coppo, L.; Sacre, S.; et al. Linkage of inflammation and oxidative stress via release of glutathionylated peroxiredoxin-2, which acts as a danger signal. Proc. Natl. Acad. Sci. USA 2014, 111, 12157–12162. [Google Scholar] [CrossRef] [PubMed]
- Naik, E.; Dixit, V.M. Mitochondrial reactive oxygen species drive proinflammatory cytokine production. J. Exp. Med. 2011, 208, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Yoo, Y.C.; Kim, H.J.; Shin, S.K.; Sohn, E.J.; Min, A.Y.; Sung, N.Y.; Kim, M.R. Aged black garlic exerts anti-inflammatory effects by decreasing no and proinflammatory cytokine production with less cytoxicity in lps-stimulated RAW264.7 macrophages and lps-induced septicemia mice. J. Med. Food 2014, 17, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Qi, J.; Feng, F.; Wang, M.D.; Bao, G.; Wang, T.; Xiang, M.; Xie, W.F. Neuroprotective effect of allicin against traumatic brain injury via akt/endothelial nitric oxide synthase pathway-mediated anti-inflammatory and anti-oxidative activities. Neurochem. Int. 2014, 68, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Ryu, J.H.; Kang, M.J.; Hwang, C.R.; Han, J.; Kang, D. Short-term heating reduces the anti-inflammatory effects of fresh raw garlic extracts on the lps-induced production of no and pro-inflammatory cytokines by downregulating allicin activity in RAW264.7 macrophages. Food Chem. Toxicol. 2013, 58, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Shen, X.; Cheng, S.; Li, P.; Du, J.; Chang, Y.; Meng, H. Evaluation of garlic cultivars for polyphenolic content and antioxidant properties. PLoS ONE 2013, 8, e79730. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Gweon, O.C.; Seo, Y.J.; Im, J.; Kang, M.J.; Kim, M.J.; Kim, J.I. Antioxidant effect of garlic and aged black garlic in animal model of type 2 diabetes mellitus. Nutr. Res. Pract. 2009, 3, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kim, M.J.; Lee, J.H.; Han, J.I.; Kim, J.H.; Sok, D.E.; Kim, M.R. Hepatoprotective effect of aged black garlic on chronic alcohol-induced liver injury in rats. J. Med. Food 2011, 14, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Lee, C.W.; Oh, S.J.; Yun, J.; Kang, M.R.; Han, S.B.; Park, H.; Jung, J.C.; Chung, Y.H.; Kang, J.S. Hepatoprotective effect of aged black garlic extract in rodents. Toxicol. Res. 2014, 30, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.; Lee, S.J.; Shin, J.H.; Sung, N.J. Antioxidant and inhibition of nitrosodimethylamine formation in marketing black garlic. J. Agric. Life Sci. 2012, 46, 151–162. [Google Scholar]
- Imai, J.; Ide, N.; Nagae, S.; Moriguchi, T.; Matsuura, H.; Itakura, Y. Antioxidant and radical scavenging effects of aged garlic extract and its constituents. Planta Med. 1994, 60, 417–420. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Shin, J.H.; Kang, M.J.; Jung, W.J.; Ryu, J.H.; Kim, R.J.; Sung, N.J. Antioxidants activity of aged red garlic. J. Life Sci. 2010, 20, 775–781. [Google Scholar]
- Shin, J.H.; Choi, D.J.; Lee, S.J.; Kwon, O.C.; Cha, J.Y.; Sung, N.J. Antioxidant activity of black garlic (Allium sativum). J. Korean Soc. Food Sci. Nutr. 2008, 37, 965–971. [Google Scholar] [CrossRef]
- Amagase, H.; Petesch, B.L.; Matsuura, H.; Kasuga, S.; Itakura, Y. Intake of garlic and its bioactive components. J. Nutr. 2001, 131, 955S–962S. [Google Scholar] [PubMed]
- Borek, C. Antioxidant health effects of aged garlic extract. J. Nutr. 2001, 131, 1010S–1015S. [Google Scholar] [PubMed]
- Gu, X.; Wu, H.; Fu, P. Allicin attenuates inflammation and suppresses HLA-B27 protein expression in ankylosing spondylitis mice. Biomed. Res. Int. 2013, 2013, 171573. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.N.; Choi, Y.W.; Kim, H.K.; Park, J.K.; Kim, H.J.; Kim, M.J.; Lee, H.W.; Kim, K.H.; Bae, S.S.; Kim, B.S.; et al. Chloroform extract of aged black garlic attenuates TNF-a-induced ros generation, VCAM-1 expression, NF-kB activation and adhesiveness for monocytes in human umbilical vein endothelial cells. Phytother. Res. 2011, 25, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Gweon, O.C.; Choi, Y.H.; Kim, J.I. Aged black garlic inhibits cyclooxygenase-2 expression and prostaglandin E2 production by phorbol 12-myristate-13-acetate through inactivation of nuclear factor-kappab. Cancer Prev. Res. 2009, 14, 161–170. [Google Scholar]
- Choi, D.J.; Lee, S.J.; Kang, M.J.; Cho, H.S.; Sung, N.J.; Shin, J.H. Physicochemical characteristics of black garlic (Allium sativum). J. Korean Soc. Food Sci. Nutr. 2008, 37, 465–471. [Google Scholar] [CrossRef]
- Choi, I.S.; Cha, H.S.; Lee, Y.S. Physicochemical and antioxidant properties of black garlic. Molecules 2014, 19, 16811–16823. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.; Perez-Jimenez, A.; Coutinho, F.; Diaz-Rosales, P.; Serra, C.A.; Panserat, S.; Corraze, G.; Peres, H.; Oliva-Teles, A. Dietary carbohydrate and lipid sources affect differently the oxidative status of european sea bass (dicentrarchus labrax) juveniles. Br. J. Nutr. 2015, 114, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Oarada, M.; Takahashi-Nakaguchi, A.; Abe, T.; Nikawa, T.; Miki, T.; Gonoi, T. Refeeding with glucose rather than fructose elicits greater hepatic inflammatory gene expression in mice. Nutrition 2015, 31, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Jameel, F.; Phang, M.; Wood, L.G.; Garg, M.L. Acute effects of feeding fructose, glucose and sucrose on blood lipid levels and systemic inflammation. Lipids Health Dis. 2014, 13, 195. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Sucrose, fructose, glucose, and their link to metabolic syndrome and cancer. Nutrition 2015, 31, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Palanisamy, N.; Kannappan, S.; Anuradha, C.V. Genistein modulates NF-kB-associated renal inflammation, fibrosis and podocyte abnormalities in fructose-fed rats. Eur. J. Pharmacol. 2011, 667, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Zhong, P.; Wu, L.; Qian, Y.; Fang, Q.; Liang, D.; Wang, J.; Zeng, C.; Wang, Y.; Liang, G. Blockage of ros and NF-κB-mediated inflammation by a new chalcone l6h9 protects cardiomyocytes from hyperglycemia-induced injuries. Biochim. Biophys. Acta 2015, 1852, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, I. Antioxidant activity of caffeic acid (3,4-dihydroxycinnamic acid). Toxicology 2006, 217, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Genaro-Mattos, T.C.; Mauricio, A.Q.; Rettori, D.; Alonso, A.; Hermes-Lima, M. Antioxidant activity of caffeic acid against iron-induced free radical generation—A chemical approach. PLoS ONE 2015, 10, e0142402. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Pyruvate is an endogenous anti-inflammatory and anti-oxidant molecule. Med. Sci. Monit. 2006, 12, RA79–RA84. [Google Scholar] [PubMed]
- Gupta, D.; Du, Y.; Piluek, J.; Jakub, A.M.; Buela, K.A.; Abbott, A.; Schuman, J.S.; SundarRaj, N. Ethyl pyruvate ameliorates endotoxin-induced corneal inflammation. Investig. Ophthalmol. Vis. Sci. 2012, 53, 6589–6599. [Google Scholar] [CrossRef] [PubMed]
- Iciek, M.; Kwiecien, I.; Wlodek, L. Biological properties of garlic and garlic-derived organosulfur compounds. Environ. Mol. Mutagen. 2009, 50, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Choi, D.J.; Lee, S.J.; Cha, J.Y.; Kim, J.G.; Sung, N.J. Changes of physicochemical components and antioxidant activity of garlic during its processing. J. Life Sci. 2008, 18, 1123–1131. [Google Scholar] [CrossRef]
- Lei, M.; Xu, M.; Zhang, Z.; Zhang, M.; Gao, Y. The analysis of saccharide in black garlic and its antioxidant activity. Adv. J. Food Sci. Technol. 2014, 6, 755–760. [Google Scholar]
- Ma, T.; Liaset, B.; Hao, Q.; Petersen, R.K.; Fjaere, E.; Ngo, H.T.; Lillefosse, H.H.; Ringholm, S.; Sonne, S.B.; Treebak, J.T.; et al. Sucrose counteracts the anti-inflammatory effect of fish oil in adipose tissue and increases obesity development in mice. PLoS ONE 2011, 6, e21647. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.J.; Cha, J.Y.; Yim, J.H.; Kim, J.H. Effect of aging temperature and time on the conversion of garlic (allium sativum) components. J. Korean Soc. Food Sci. Nutr. 2011, 40, 84–88. [Google Scholar] [CrossRef]
- Yerneni, K.K.; Bai, W.; Khan, B.V.; Medford, R.M.; Natarajan, R. Hyperglycemia-induced activation of nuclear transcription factor kappab in vascular smooth muscle cells. Diabetes 1999, 48, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, A.C.; Adeli, K. Fructose and the metabolic syndrome: Pathophysiology and molecular mechanisms. Nutr. Rev. 2007, 65, S13–S23. [Google Scholar] [CrossRef] [PubMed]
- Nagae, S.; Ushijima, M.; Hatono, S.; Imai, J.; Kasuga, S.; Matsuura, H.; Itakura, Y.; Higashi, Y. Pharmacokinetics of the garlic compound s-allylcysteine. Planta Med. 1994, 60, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Steiner, M.; Li, W. Aged garlic extract, a modulator of cardiovascular risk factors: A dose-finding study on the effects of age on platelet functions. J. Nutr. 2001, 131, 980S–984S. [Google Scholar] [PubMed]
- Hoshino, T.; Kashimoto, N.; Kasuga, S. Effects of garlic preparations on the gastrointestinal mucosa. J. Nutr. 2001, 131, 1109S–1113S. [Google Scholar] [PubMed]
- Kang, M.J.; Shin, J.H.; Yoon, H.S.; Jeong, S.H. Method for Manufacturing Black Garlic and the Extract. Thereof. Patent No. 10-1236251, 18 February 2013. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Lyckander, I.M.; Malterud, K.E. Lipophilic flavonoids from orthosiphon spicatus as inhibitors of 15-lipoxygenase. Acta Pharm. Nord. 1992, 4, 159–166. [Google Scholar]
- Miron, T.; Shin, I.; Feigenblat, G.; Weiner, L.; Mirelman, D.; Wilchek, M.; Rabinkov, A. A spectrophotometric assay for allicin, alliin, and alliinase (alliin lyase) with a chromogenic thiol: Reaction of 4-mercaptopyridine with thiosulfinates. Anal. Biochem. 2002, 307, 76–83. [Google Scholar] [CrossRef]
- Schwimmer, S.; Guadagni, D.G. Relation between olfactory threshold concentration and pyruvic acid content of onion juice. J. Food Sci. 1962, 27, 94–97. [Google Scholar] [CrossRef]
- Freeman, G.G.; Mcbreen, F. A rapid spectrophotometric methods of determination of thiosulfinate in onion (Allium cepa) and its significance in flavor studies. Biochem. Soc. Trans. 1973, 1, 1150–1152. [Google Scholar] [CrossRef]
- Park, H.J.; Jeon, B.T.; Kim, H.C.; Roh, G.S.; Shin, J.H.; Sung, N.J.; Han, J.; Kang, D. Aged red garlic extract reduces lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophages and acute pulmonary inflammation through haeme oxygenase-1 induction. Acta Physiol. 2012, 205, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Garlic extracts (FRGE, HRGE, and ABGE) are available from the authors.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Garlic Extracts | Pyruvate (μM/g) | Thiosulfinate (μM/g) | Free Sugars (mg/g) | Allicin (mg/g) | SAC (mg/g) | |
|---|---|---|---|---|---|---|
| Glucose | Fructose | |||||
| FRGE | 486.71 ± 12.08 | 6.50 ± 0.29 | - | 7.07 ± 0.08 | 3.62 ± 0.01 | 1.24 ± 9.22 |
| HRGE | 1023.90 ± 9.83 | 6.61 ± 0.25 | 0.21 ± 0.02 | 7.30 ± 0.25 | 1.93 ± 0.02 | 1.82 ± 8.24 |
| ABGE | 2456.54 ± 23.93 | 91.22 ± 0.54 | 2.10 ± 0.05 | 40.02 ± 0.71 | - | 2.12 ± 10.17 |
| Gene Name | GenBank Acc. No. | Primer Sequences (5′–3′) | Expected Size (bp) |
|---|---|---|---|
| iNOS | NM_010927 | Sense: TTGACGCTCGGAACTGTAGCA | 492 |
| Antisense: TGCCCATGTACCAACCATTGA | |||
| COX-2 | NM_011198 | Sense: CCCAGAGCTCCTTTTCAACCA | 382 |
| Antisense: TGCAGCCATTTCCTTCTCTCC | |||
| IL-1 | NM_010554 | Sense: GGAGAGCCGGGTGACAGTAT | 364 |
| Antisense: GGGCTGGTCTTCTCCTTGAG | |||
| IL-6 | NM_031168 | Sense: CTTCACAAGTCCGGAGAGGAG | 489 |
| Antisense: TGGTCTTGGTCCTTAGCCACT | |||
| GAPDH | GU214026 | Sense: CTA AAG GGC ATC CTG GGC | 201 |
| Antisense: TTA CTC CTT GGA GGC CAT |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.Y.; Ryu, J.H.; Shin, J.-H.; Kang, M.J.; Kang, J.R.; Han, J.; Kang, D. Comparison of Anti-Oxidant and Anti-Inflammatory Effects between Fresh and Aged Black Garlic Extracts. Molecules 2016, 21, 430. https://doi.org/10.3390/molecules21040430
Jeong YY, Ryu JH, Shin J-H, Kang MJ, Kang JR, Han J, Kang D. Comparison of Anti-Oxidant and Anti-Inflammatory Effects between Fresh and Aged Black Garlic Extracts. Molecules. 2016; 21(4):430. https://doi.org/10.3390/molecules21040430
Chicago/Turabian StyleJeong, Yi Yeong, Ji Hyeon Ryu, Jung-Hye Shin, Min Jung Kang, Jae Ran Kang, Jaehee Han, and Dawon Kang. 2016. "Comparison of Anti-Oxidant and Anti-Inflammatory Effects between Fresh and Aged Black Garlic Extracts" Molecules 21, no. 4: 430. https://doi.org/10.3390/molecules21040430
APA StyleJeong, Y. Y., Ryu, J. H., Shin, J.-H., Kang, M. J., Kang, J. R., Han, J., & Kang, D. (2016). Comparison of Anti-Oxidant and Anti-Inflammatory Effects between Fresh and Aged Black Garlic Extracts. Molecules, 21(4), 430. https://doi.org/10.3390/molecules21040430