
Antioxidant and DNA Damage Protecting Activity of Exopolysaccharides from the Endophytic Bacterium Bacillus cereus SZ1


Abstract
:1. Introduction
2. Results
2.1. Strain Isolation, Biochemical and Molecular Characterization


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Result | Characteristics | Result |
|---|---|---|---|
| Gram staining | + a | d-glucose | + |
| Swelling spore | − b | l-arabinose | − |
| Aerobic growth | + | d-xylose | + |
| Anaerobic growth | − | d-mannitol | + |
| Nitrate reduction | + | Lactose | + |
| Utilization of citrate | + | Raffinose | + |
| Voges-proskauer test | + | Cellobiose | + |
| Milk solidification | + | Indole production | − |
| Milk peptonization | + | Starch hydrolysis | + |
| Catalase | + | Indole production | − |
| Oxidase | + | Gas from glucose | + |
| urease | − | Growth temperature range | 10–40 °C |
| Contactwise | + | Tolerance to NaCl | <10% |
2.2. Growth Curve and EPS Production
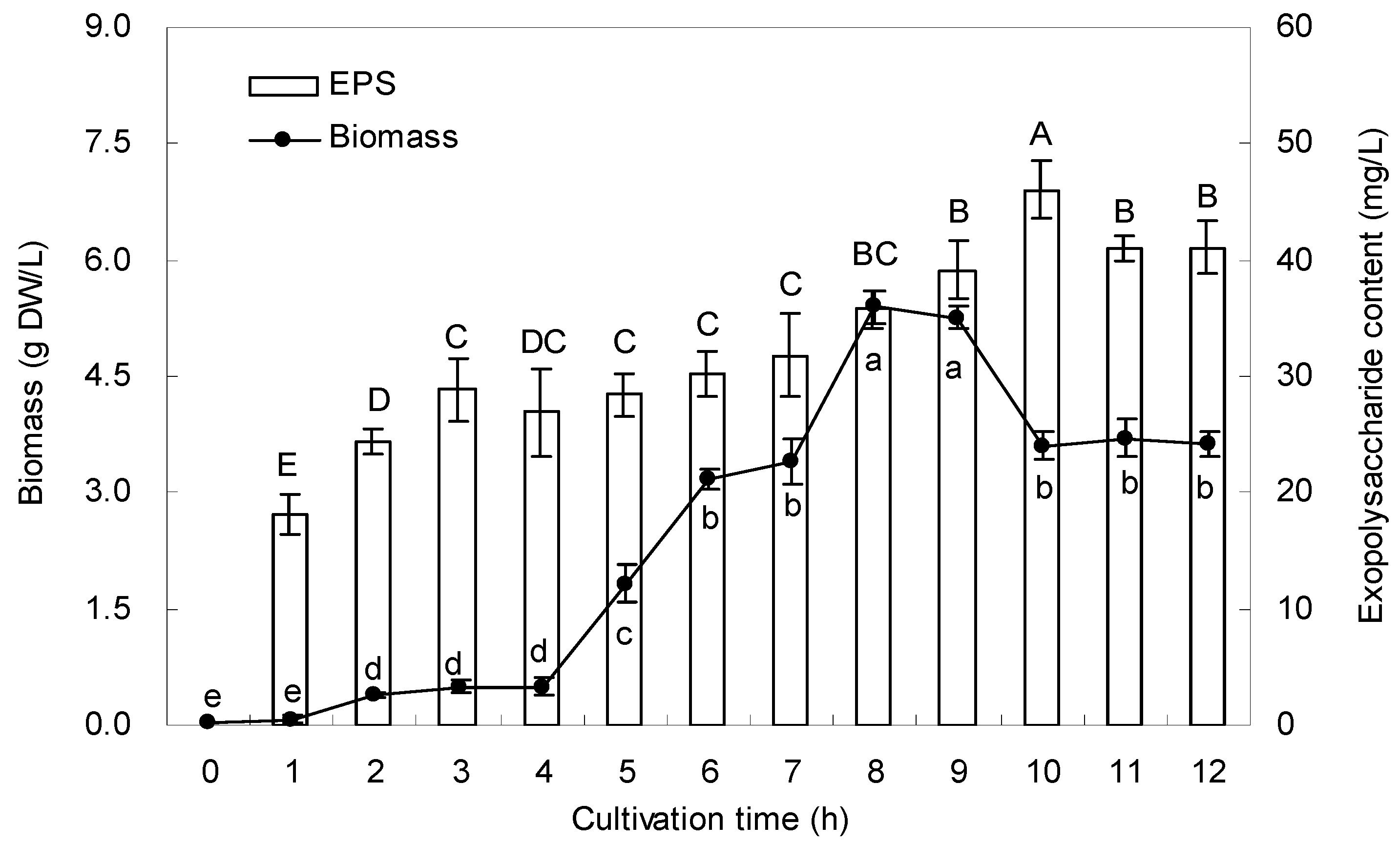
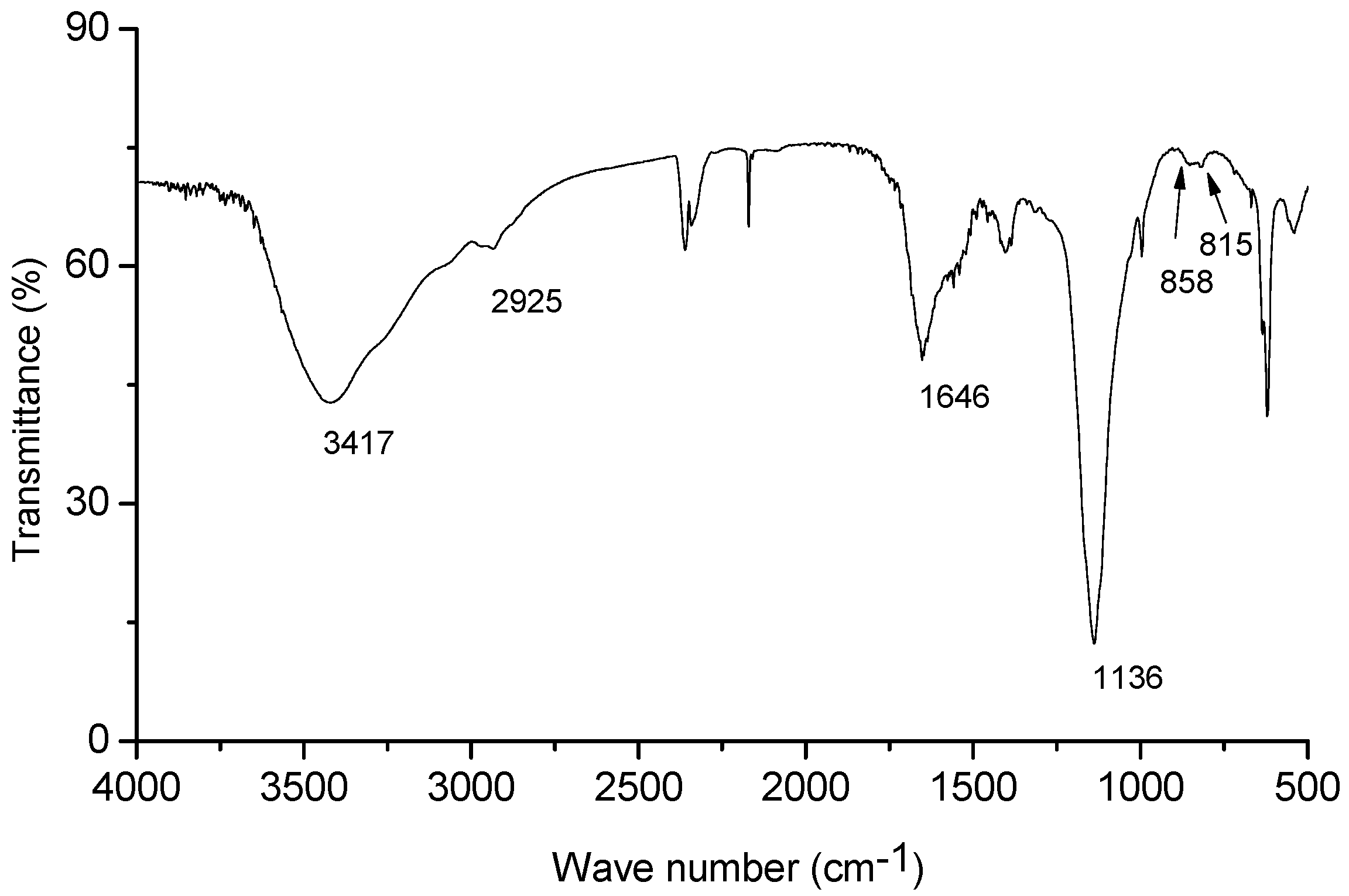
2.3. Chemical Analysis of EPS

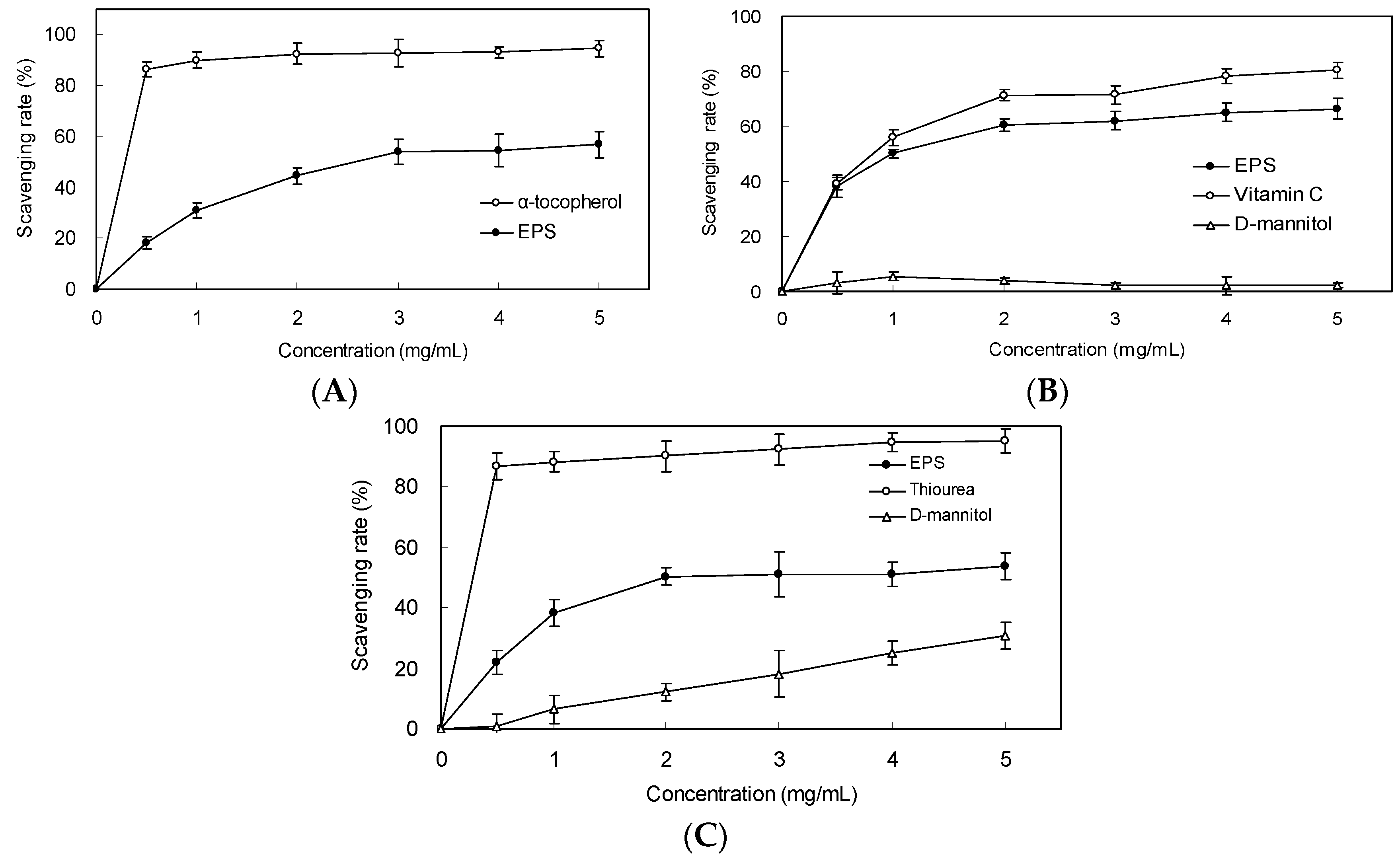
2.4. Scavenging Effect on DPPH Radicals
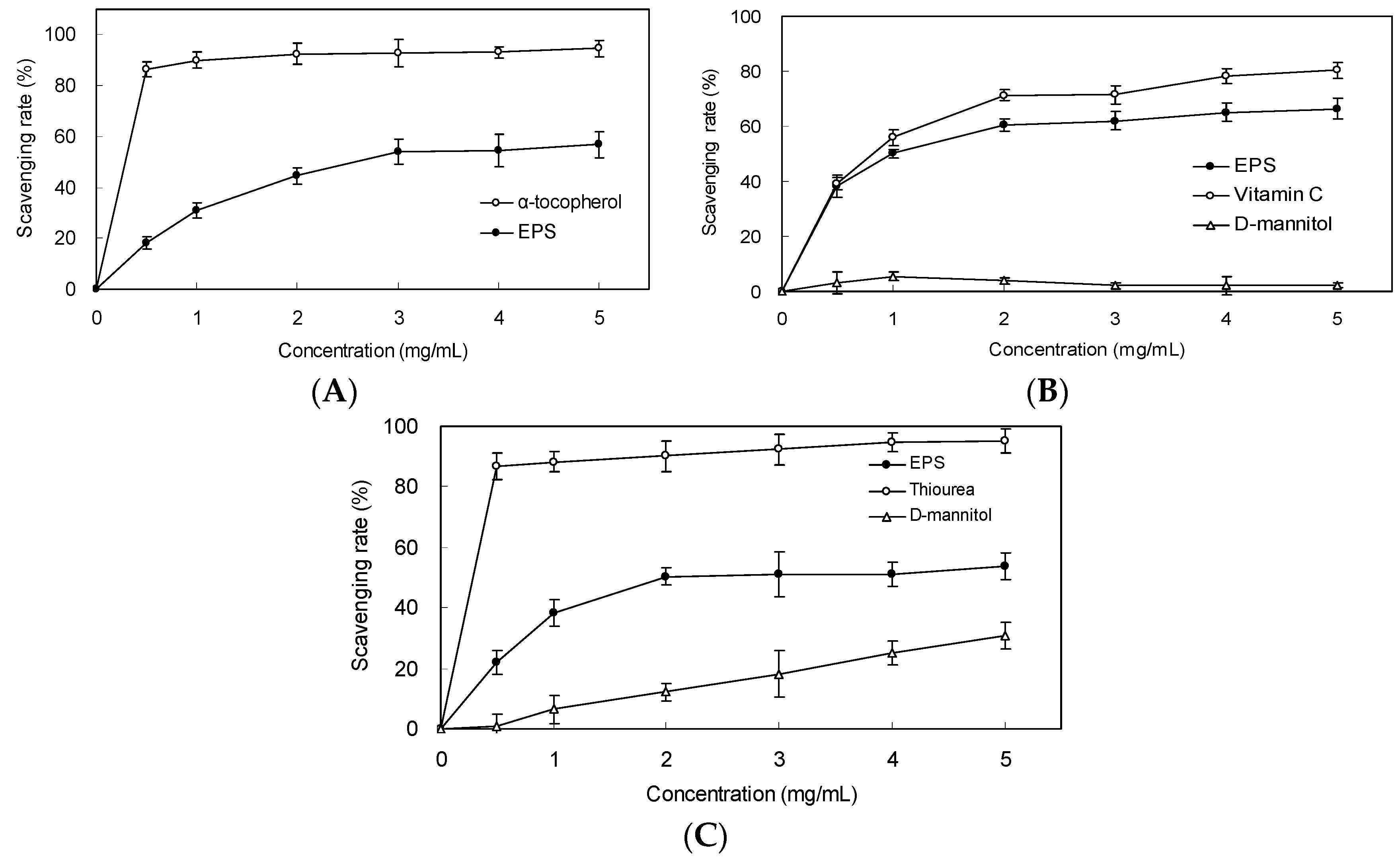
2.5. Scavenging Effect on Superoxide Anion and Hydroxyl Radicals
2.6. Reducing Power
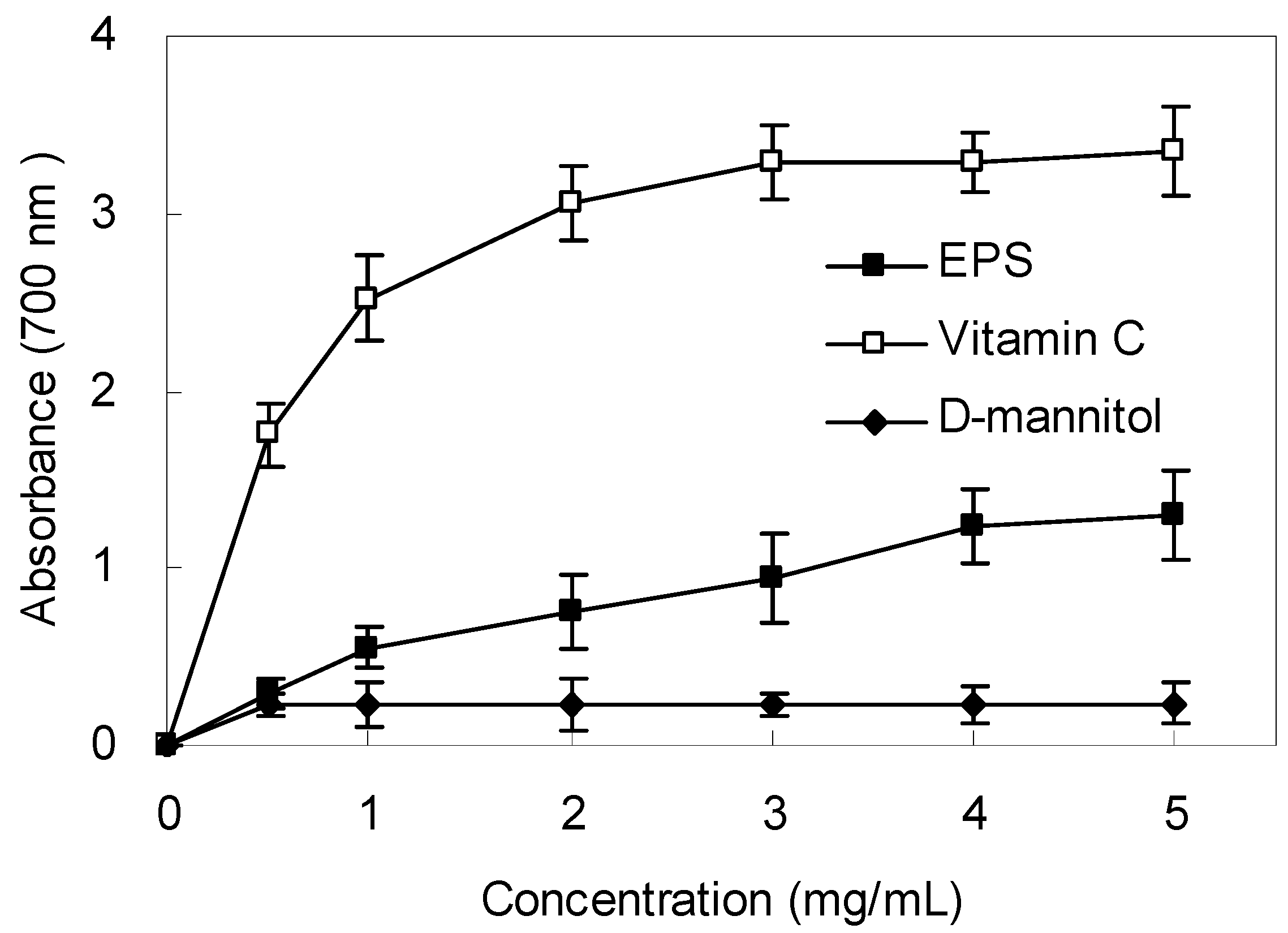
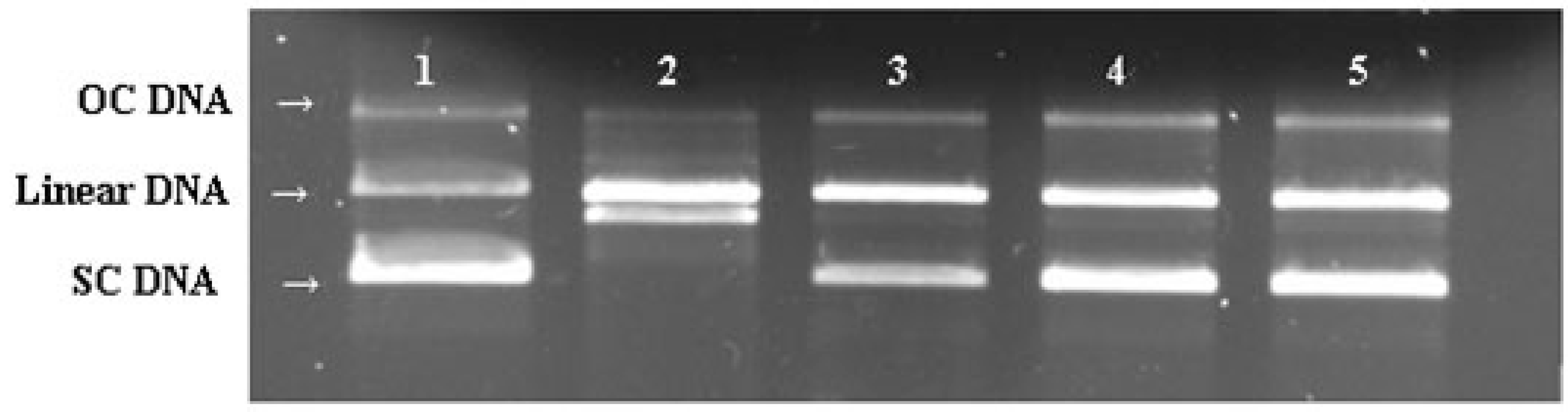
2.7. DNA Protective Effect

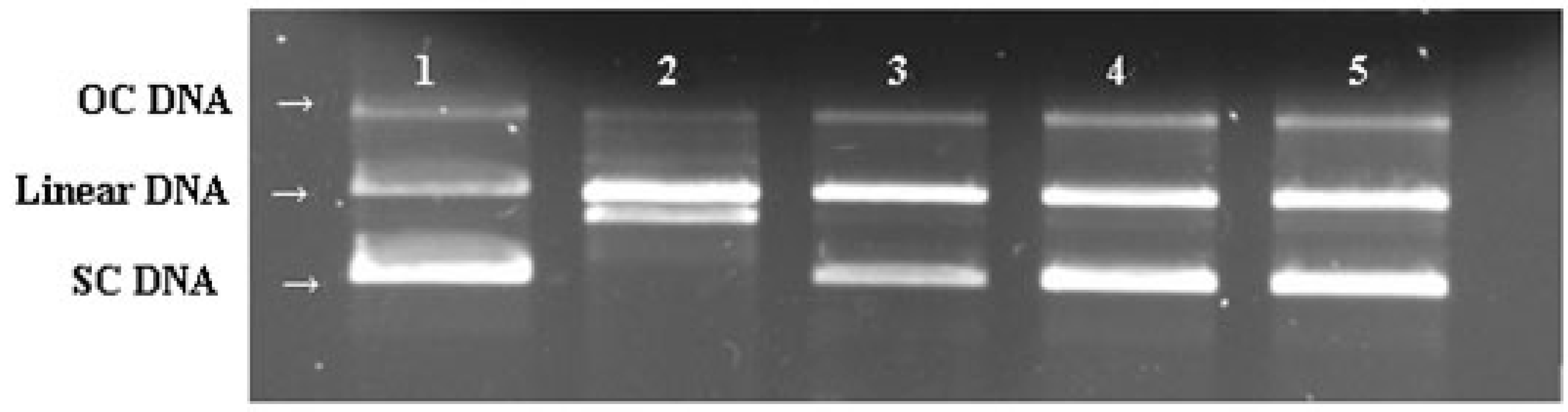
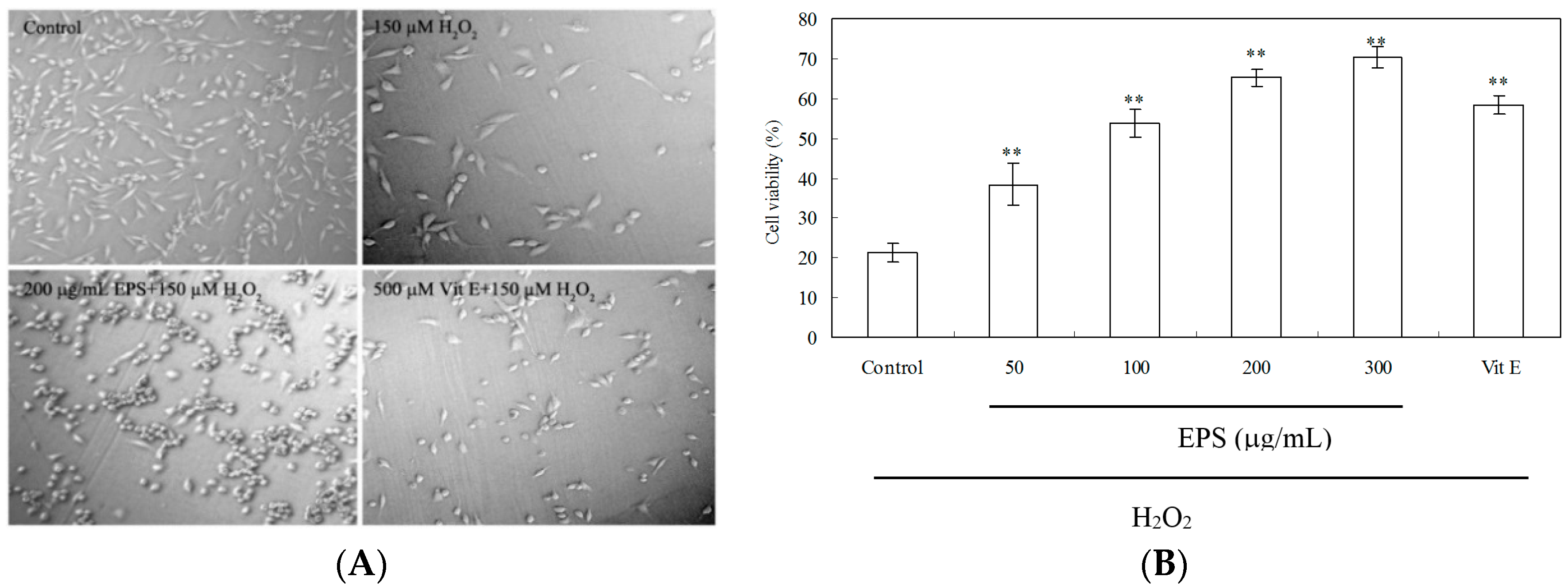
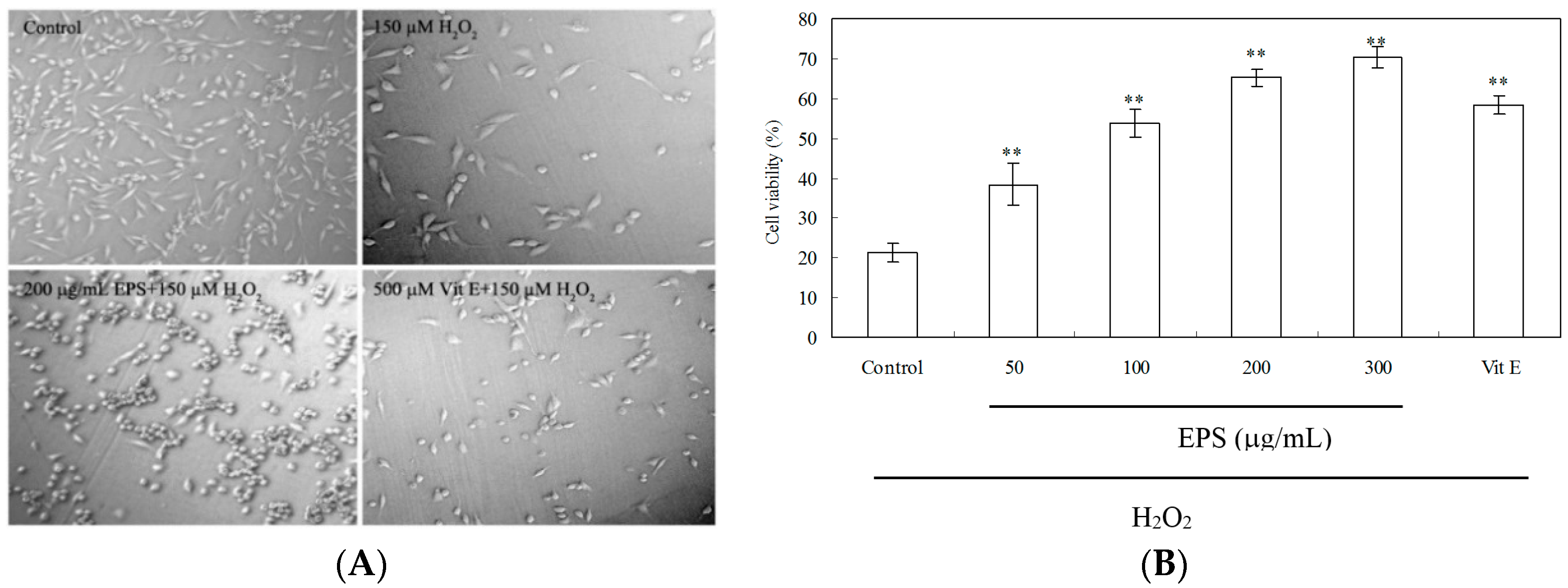
2.8. Protection of PC12 Cells from H2O2-Induced Injury

3. Discussion
4. Experimental Section
4.1. Reagents and Materials
4.2. Isolation and Identification of Selected Endophytic Bacteria
4.3. Cultivation and Extraction of EPS
4.4. Chemical Analysis of EPS
4.5. Radical Scavenging Property and Antioxidant Activity in Vitro
4.5.1. DPPH Radical Scavenging Assay
4.5.2. Superoxide Radical Scavenging Assay
4.5.3. Hydroxyl Radical Scavenging Assay
4.5.4. Measurement of Reducing Power
4.5.5. Chemiluminescence Assay for H2O2-Induced DNA Damage
4.5.6. DNA Nicking Assay for Hydroxyl Radical Scavenging Activity
4.6. Culture of PC12 Cells and Viability Assay
4.7. MDA, LDH, GSH, CAT and SOD Assays
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pryor, W.A. Oxy-radicals and related species: Their formation, lifetimes, and reactions. Annu. Rev. Physiol. 1986, 48, 657–667. [Google Scholar] [PubMed]
- Aruoma, O.I. Free radicals, oxidative stress, and antioxidants in human health and disease. J. Am. Oil Chem. Soc. 1998, 75, 199–212. [Google Scholar] [CrossRef]
- Velioglu, Y.S.; Mazza, G.; Gao, L.; Oomah, B.D. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Denis, M.C.; Furtos, A.; Dudonné, S.; Montoudis, A.; Garofalo, C.; Desjardins, Y.; Delvin, E.; Levy, E. Apple peel polyphenols and their beneficial actions on oxidative stress and inflammation. PLoS ONE 2013, 8, e53725. [Google Scholar]
- Giampieri, F.; Alvarez-Suarez, J.M.; Battino, M. Strawberry and human health: Effects beyond antioxidant activity. J. Agric. Food Chem. 2014, 62, 3867–3876. [Google Scholar] [CrossRef] [PubMed]
- Asker, M.M.S.; Ahmed, Y.M.; Ramadan, M.F. Chemical characteristics and antioxidant activity of exopolysaccharide fractions from Microbacterium terregens. Carbohydr. Polym. 2009, 77, 563–567. [Google Scholar] [CrossRef]
- Liu, J.; Luo, J.G.; Ye, H.; Sun, Y.; Lu, Z.X.; Zeng, X.X. In vitro and in vivo antioxidant activity of exopolysaccharides from endophytic bacterium Paenibacillus polymyxa EJS-3. Carbohydr. Polym. 2010, 82, 1278–1283. [Google Scholar] [CrossRef]
- Zhang, H.W.; Song, Y.C.; Tan, R.X. Biology and chemistry of endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.Y.; Cai, Y.Z.; Xing, J.; Corke, H.; Sun, M. A potential antioxidant resource: Endophytic fungi from medicinal plants. Econ. Bot. 2007, 61, 14–30. [Google Scholar] [CrossRef]
- Liu, X.; Dong, M.; Chen, X.; Jiang, M.; Lv, X.; Yan, G. Antioxidant activity and phenolics of an endophytic Xylaria sp. from Ginkgo biloba. Food Chem. 2007, 105, 548–554. [Google Scholar] [CrossRef]
- Liu, J.; Luo, J.G.; Ye, H.; Sun, Y.; Lu, Z.X.; Zeng, X.X. Production, characterization and antioxidant activities in vitro of exopolysaccharides from endophytic bacterium Paenibacilluspolymyxa EJS-3. Carbohydr. Polym. 2009, 78, 275–281. [Google Scholar] [CrossRef]
- Vijayabaskar, P.; Babinastarlin, S.; Shankar, T.; Sivakumar, T.; Anandapandian, K.T.K. Quantification and characterization of exopolysaccharides from Bacillus subtilis (MTCC 121). Adv. Biol. Res. 2011, 5, 71–76. [Google Scholar]
- Arena, A.; Maugeri, T.L.; Pavone, B.; Iannello, D.; Gugliandolo, C.; Bisignano, G. Antiviral and immunoregulatory effect of a novel exopolysaccharide from a marine thermotolerant Bacillus licheniformis. Int. Immunopharm. 2006, 6, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Kodali, V.P.; Perali, R.S.; Sen, R. Purification and partial elucidation of the structure of an antioxidant carbohydrate biopolymer from the probiotic bacterium Bacillus coagulans RK-02. J. Nat. Prod. 2011, 74, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Wu, J.H.; Huang, W.Y.; Tan, R.X. Laccase production by Monotospora sp., an endophytic fungus in Cynodon dactylon. Bioresour. Technol. 2006, 97, 786–789. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Zheng, L.P.; Zhang, B.; Zou, T. Stimulation of artemisinin synthesis by combined cerebroside and nitric oxide elicitation in Artemisia annua hairy roots. Appl. Microbiol. Biotechnol. 2009, 85, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.W.; Wang, J.W. Antioxiant potential and DNA damage protecting activity of aqueous extract from Armillaria mellea. J. Food Biochem. 2012, 36, 139–148. [Google Scholar] [CrossRef]
- Santhiya, D.; Subramanian, S.; Natarajan, K.A. Surface chemical studies on sphalerite and galena using extracellular polysaccharides isolated from Bacillus polymyxa. J. Colloid. Interface. Sci. 2002, 256, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Ananthi, S.; Raghavendran, H.R.B.; Sunil, A.G.; Gayathri, V.; Ramakrishnan, G.; Vasanthi, H.R. In vitro antioxidant and in vivo anti-inflammatory potential of crude polysaccharide from Turbinaria ornate (Marine brown alga). Food Chem. Toxicol. 2010, 48, 187–192. [Google Scholar]
- Yan, J.K.; Li, L.; Wang, Z.M.; Wu, J.W. Structural elucidation of an exopolysaccharide from mycelial fermentation of a Tolypocladium sp. fungus isolated from wild Cordyceps sinensis. Carbohydr. Polym. 2010, 70, 125–130. [Google Scholar] [CrossRef]
- Lu, H.; Zou, W.X.; Meng, J.C.; Hu, J.; Tan, R.X. New bioactive metabolites produced by Colletotrichum sp., an endophytic fungus in Artemisia annua. Plant Sci. 2000, 151, 67–73. [Google Scholar] [CrossRef]
- Li, J.; Zhao, G.-Z.; Huang, H.-Y.; Qin, S.; Zhu, W.-Y.; Zhao, L.-X.; Xu, L.-H.; Zhang, S.; Li, W.-J.; Strobel, G. Isolation and characterization of culturable endophytic actinobacteria associated with Artemisia annua L. Anton. Leeuwenhoek 2012, 101, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Palaniappan, P.; Chauhan, P.S.; Saravanan, V.S.; Anandham, R.; Sa, T. Isolation and characterization of plant growth promoting endophytic bacterial isolates from root nodule of Lespedeza sp. Biol. Fertil. Soils 2010, 46, 807–816. [Google Scholar] [CrossRef]
- Islam, S.M.A.; Math, R.K.; Kim, J.M.; Yun, M.G.; Cho, J.J.; Kim, E.J.; Lee, Y.H.; Yun, H.D. Effect of plant age on endophytic bacterial diversity of balloon flower (Platycodon grandiflorum) root and their antimicrobial activities. Curr. Microbiol. 2010, 61, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.-M.; Farag, M.A.; Hu, C.-H.; Reddy, M.S.; Wei, H.-X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [PubMed]
- Pavlo, A.; Leonid, O.; Iryna, Z.; Natalia, K.; Maria, P.A. Endophytic bacteria enhancing growth and disease resistance of potato (Solanum tuberosum L.). Biol. Control. 2011, 56, 43–49. [Google Scholar] [CrossRef]
- Duke, S.O. Artemisinin, constituent of annual wormwood (Artemisia annua), is a selective phytotoxin. Weed Sci. 1987, 35, 499–505. [Google Scholar]
- Patil, S.V.; Bathe, G.A.; Patil, A.V.; Patil, R.H.; Salunkea, B.K. Production of bioflocculant exopolysaccharide by Bacillus subtilis. Adv. Biotech. 2009, 9, 14–18. [Google Scholar]
- Fusconi, R.; Godinho, M.J.L. Screening for exopolysaccharide-producing bacteria from sub-tropical polluted groundwater. Braz. J. Biol. 2002, 62, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Kodali, V.P.; Sen, R. Antioxidant and free radical scavenging activities of an exopolysaccharide from a probiotic bacterium. Biotechnol. J. 2008, 3, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.R.; Song, N.-E.; Kim, J.-H.; Nho, Y.-C.; Baik, S.-H. Exopolysaccharide produced by Bacillus licheniformis strains isolated from Kimchi. J. Gen. Appl. Microbiol. 2011, 57, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Wang, J.W.; Fang, L.; Gao, X.D.; Tan, R.X. Free radical scavenging and antioxidant activities of EPS2, an exopolysaccharide produced by a marine filamentous fungus Keissleriella sp. YS 4108. Life Sci. 2004, 75, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Wiseman, H.; Kaur, H.; Halliwell, B. DNA damage and cancer: measurement and mechanism. Cancer Lett. 1995, 93, 113–120. [Google Scholar] [CrossRef]
- Yen, G.C.; Shih, P.H.; Song, T.Y.; Tsai, M.C. Antioxidant properties of water-soluble polysaccharides from Antrodia cinnamomea in submerged culture. Food Chem. 2007, 104, 1115–1122. [Google Scholar]
- Ma, W.J.; Cao, E.H.; Zhang, J.; Qin, J.F. Phenanthroline-Cu complex-mediated chemiluminescence of DNA and its potential use in antioxidation evaluation. J. Photochem. Photobiol. B Biol. 1998, 44, 63–68. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Byun, H.G.; Kim, S.K. Protective effect of an antioxidative peptide purified from gastrointestinal digests of oyster, Crassostrea gigas against free radical induced DNA damage. Bioresour. Technol. 2008, 99, 3365–3371. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Park, S.Y.; Kim, J.K. Curcuminoids from Curcuma longa L. (Zingiberaceae) that protect PC12 rat pheochromocytoma and normal human umbilical vein endothelial cells from βA(1–42) insult. Neurosci. Lett. 2001, 303, 57–61. [Google Scholar] [CrossRef]
- Sun, C.; Shan, C.Y.; Gao, X.D.; Tan, R.X. Protection of PC12 cells from hydrogen peroxide-induced injury by EPS2, an exopolysaccharide from a marine filamentous fungus Keissleriella sp. YS4108. J. Biotechnol. 2005, 115, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-Y.; Jin, M.-M.; Meng, J.; Gao, S.-M.; Lu, R.-R. Exopolysaccharide from Lactobacillus planterum LP6, Antioxidation and the effect on oxidative stress. Carbohydr. Polym. 2013, 98, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, J.S.; Steinauer, K.K.; Hornung, B.; Irish, J.M.; Lecane, P.; Birrell, G.W.; Peehl, D.M.; Knox, S.J. Role of glutathione depletion and reactive oxygen species generation in apoptotic signaling in a human B lymphoma cell line. Cell Death Differ. 2002, 9, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Barzanti, R.; Ozino, F.; Bazzicalupo, M.; Gabbrielli, R.; Galardi, F.; Gonnelli, C.; Mengoni, A. Isolation and characterization of endophytic bacteria from the nickel hyperaccumulator plant Alyssum bertolonii. Microb. Ecol. 2007, 53, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Sneath, P.H.A. Endospore-forming gram-positive rods and cocci. Bergey’s Manual of Systematic Bacteriology 1986, 2, 1104–1207. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic. Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Staub, A.M. Removal of protein-Sevag method. Method Carbohyd. Chem. 1965, 5, 5–6. [Google Scholar]
- Harbone, J.B. Phytochemical Methods; Chapman and Hall: London, UK, 1973. [Google Scholar]
- Hedge, J.E.; Hofreiter, B.T. Determination of total carbohydrate by anthrone reagent. In Carbohydrate Chemistry; Whistler, R.L., Be Miller, J.N., Eds.; Academic Press: New York, NY, USA, 1962. [Google Scholar]
- Bradford, M.M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Chem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hatano, T.; Edamatsu, R.; Mori, A.; Fujita, Y.; Yasuhara, T.; Yoshida, T.; Okuda, T. Effects of the interaction of tannins with co-existing substances. VI. Effects of tannins and related polyphenols on superoxide anion radical, and on 1,1-diphenylperylhydrazyl radical. Chem. Pharm. Bull. 1989, 37, 2016–2021. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C.; Aruoma, O.L. The deoxyribose method: A simple test-tube assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Ahmadi, F.; Kadivar, M.; Shahedi, M. Antioxidant activity of Kelussia odoratissima Mozaff in model and food systems. Food Chem. 2007, 105, 57–64. [Google Scholar] [CrossRef]
- Kitts, D.D.; Wijewickreme, A.N.; Hu, C. Antioxidant properties of a north American ginseng extract. Mol. Cell Biochem. 2000, 203, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the EPS are available from the authors.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, L.P.; Zou, T.; Ma, Y.J.; Wang, J.W.; Zhang, Y.Q. Antioxidant and DNA Damage Protecting Activity of Exopolysaccharides from the Endophytic Bacterium Bacillus cereus SZ1. Molecules 2016, 21, 174. https://doi.org/10.3390/molecules21020174
Zheng LP, Zou T, Ma YJ, Wang JW, Zhang YQ. Antioxidant and DNA Damage Protecting Activity of Exopolysaccharides from the Endophytic Bacterium Bacillus cereus SZ1. Molecules. 2016; 21(2):174. https://doi.org/10.3390/molecules21020174
Chicago/Turabian StyleZheng, Li Ping, Tin Zou, Yan Jun Ma, Jian Wen Wang, and Yu Qing Zhang. 2016. "Antioxidant and DNA Damage Protecting Activity of Exopolysaccharides from the Endophytic Bacterium Bacillus cereus SZ1" Molecules 21, no. 2: 174. https://doi.org/10.3390/molecules21020174
APA StyleZheng, L. P., Zou, T., Ma, Y. J., Wang, J. W., & Zhang, Y. Q. (2016). Antioxidant and DNA Damage Protecting Activity of Exopolysaccharides from the Endophytic Bacterium Bacillus cereus SZ1. Molecules, 21(2), 174. https://doi.org/10.3390/molecules21020174