
Metabolic Profiling and Antioxidant Assay of Metabolites from Three Radish Cultivars (Raphanus sativus)


 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Anthocyanin Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No.a | RT (min) | Trivial Name | Seo Ho | Man Tang Hong | Hong Peng No. 1 |
|---|---|---|---|---|---|
| 1 | 8.073 | Pelargonidin 3-diglucoside-5-glucoside | ND b | 0.08 ± 0.00 | 0.01 ± 0.00 |
| 2 | 8.966 | Pelargonidin 3-diglucoside-5-(malonyl)glucoside | ND | 0.06 ± 0.00 | 0.01 ± 0.00 |
| 3 | 10.846 | Pelargonidin 3-(caffeoyl)diglucoside-5-glucoside | ND | 0.02 ± 0.01 | ND |
| 4 | 12.172 | Cyanidin 3-(glucosyl)rhamnoside | ND | 0.09 ± 0.01 | 0.01 ± 0.00 |
| 5 | 12.994 | Pelargonidin 3-(p-coumaroyl)diglucoside-5-(malonyl)glucoside | ND | 0.16 ± 0.01 | 0.04 ± 0.00 |
| 6 | 13.309 | Pelargonidin 3-(caffeoyl)diglucoside-5-(malonyl)glucoside | ND | 0.02 ± 0.00 | ND |
| 7 | 14.668 | Pelargonidin 3-(p-coumaroyl)diglucoside-5-(malonyl)glucoside | ND | 0.22 ± 0.01 | 0.02 ± 0.00 |
| 8 | 15.424 | Pelargonidin 3-(p-coumaroyl)(caffecoyl)diglucoside-5-(malonyl)glucoside | ND | 0.19 ± 0.01 | 0.01 ± 0.00 |
| 9 | 15.954 | Pelargonidin 3-(feruloyl)diglucoside-5-(malonyl)glucoside | ND | 0.02 ± 0.00 | ND |
| 10 | 16.674 | Pelargonidin 3-(p-coumaroyl)diglucoside-5-(malonyl)glucoside | ND | 0.45 ± 0.02 | 0.10 ± 0.00 |
| 11 | 17.782 | Pelargonidin 3-(feruloyl)diglucoside-5-(malonyl)glucoside | ND | 0.47 ± 0.01 | 0.03 ± 0.00 |
| 12 | 20.916 | Pelargonidin 3-(feruloyl)(caffecoyl)diglucoside-5-(malonyl)glucoside | ND | 0.04 ± 0.01 | ND |
| 13 | 25.130 | Pelargonidin 3-(p-coumaroyl)(feruloy)diglucoside-5-(malonyl)glucoside | ND | 0.07 ± 0.01 | ND |
| Total | ND | 1.89 ± 0.07 | 0.23 ± 0.00 | ||
2.2. Total Phenolics and Flavonoid Contents
| Cultivars | Total Phenolics (mg/g) | Total Flavonoid (mg/g) |
|---|---|---|
| Seo Ho | 0.0147 ± 0.0010 b | 0.0060 ± 0.0007 b |
| Man Tang Hong | 0.0664 ± 0.0033 a | 0.0096 ± 0.0004 a |
| Hong Feng No. 1 | 0.0088 ± 0.0006 b | 0 c |
2.3. Metabolic Profiles among Radish Cultivars Using GC-TOFMS Analysis
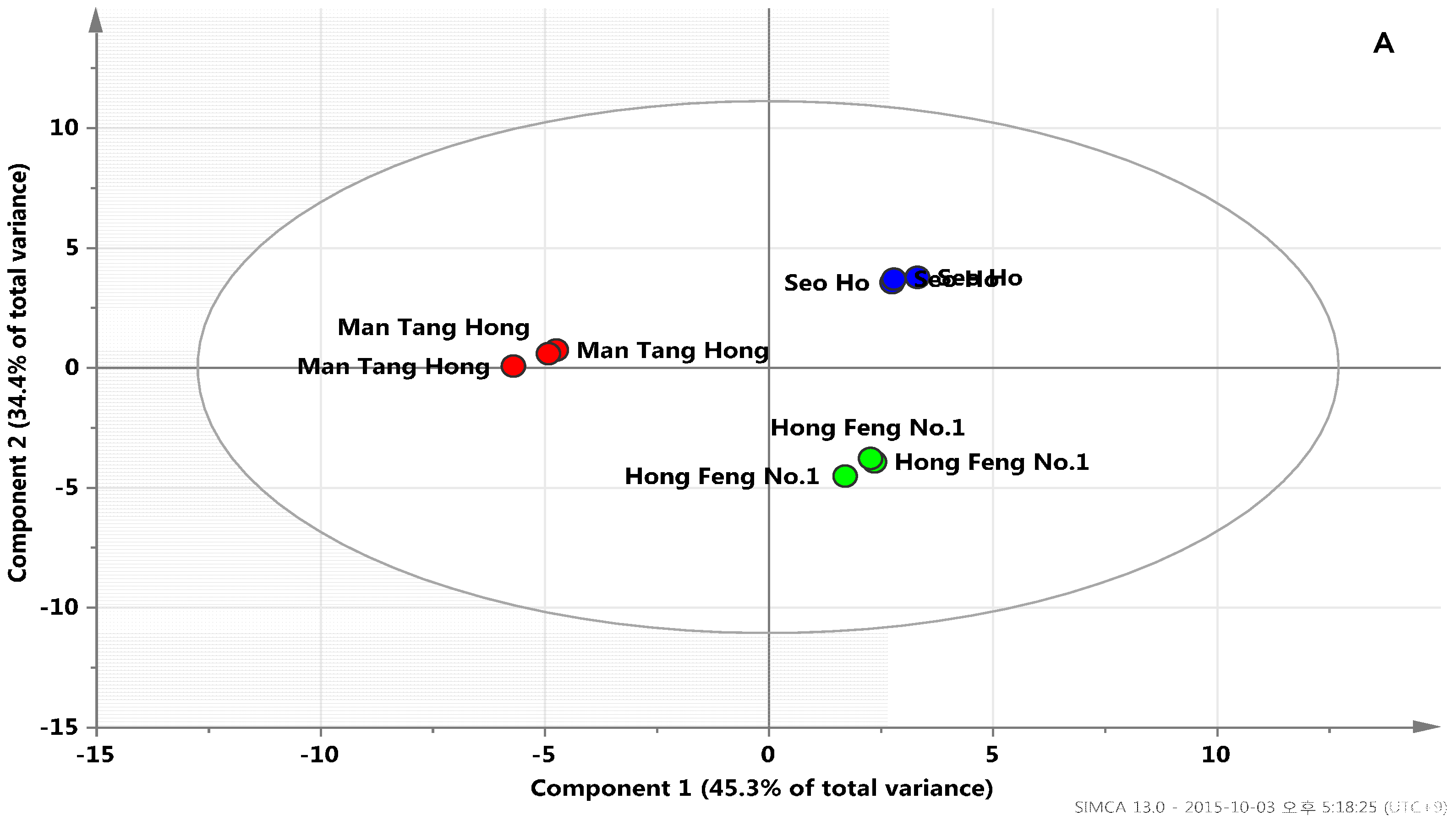
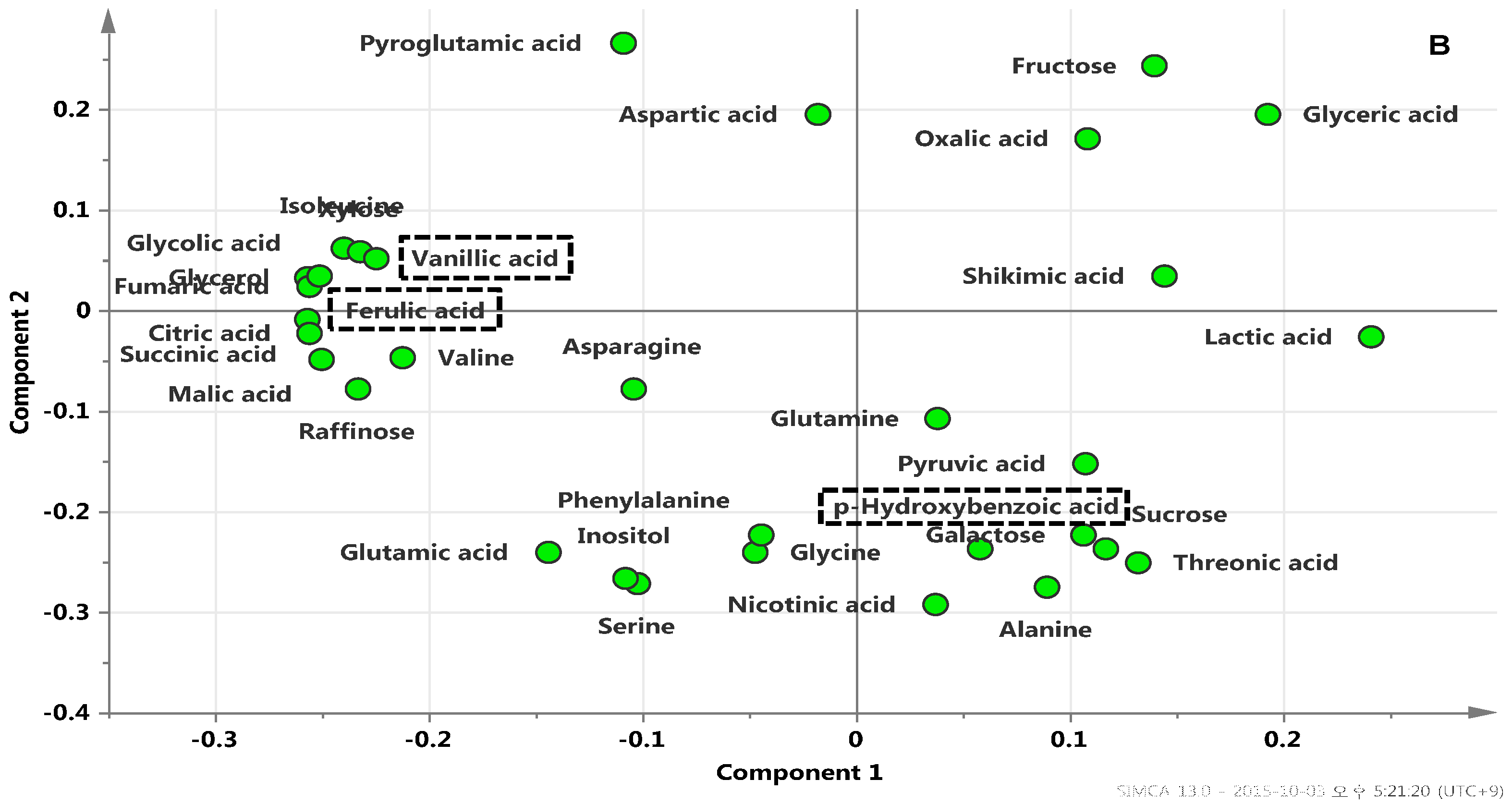
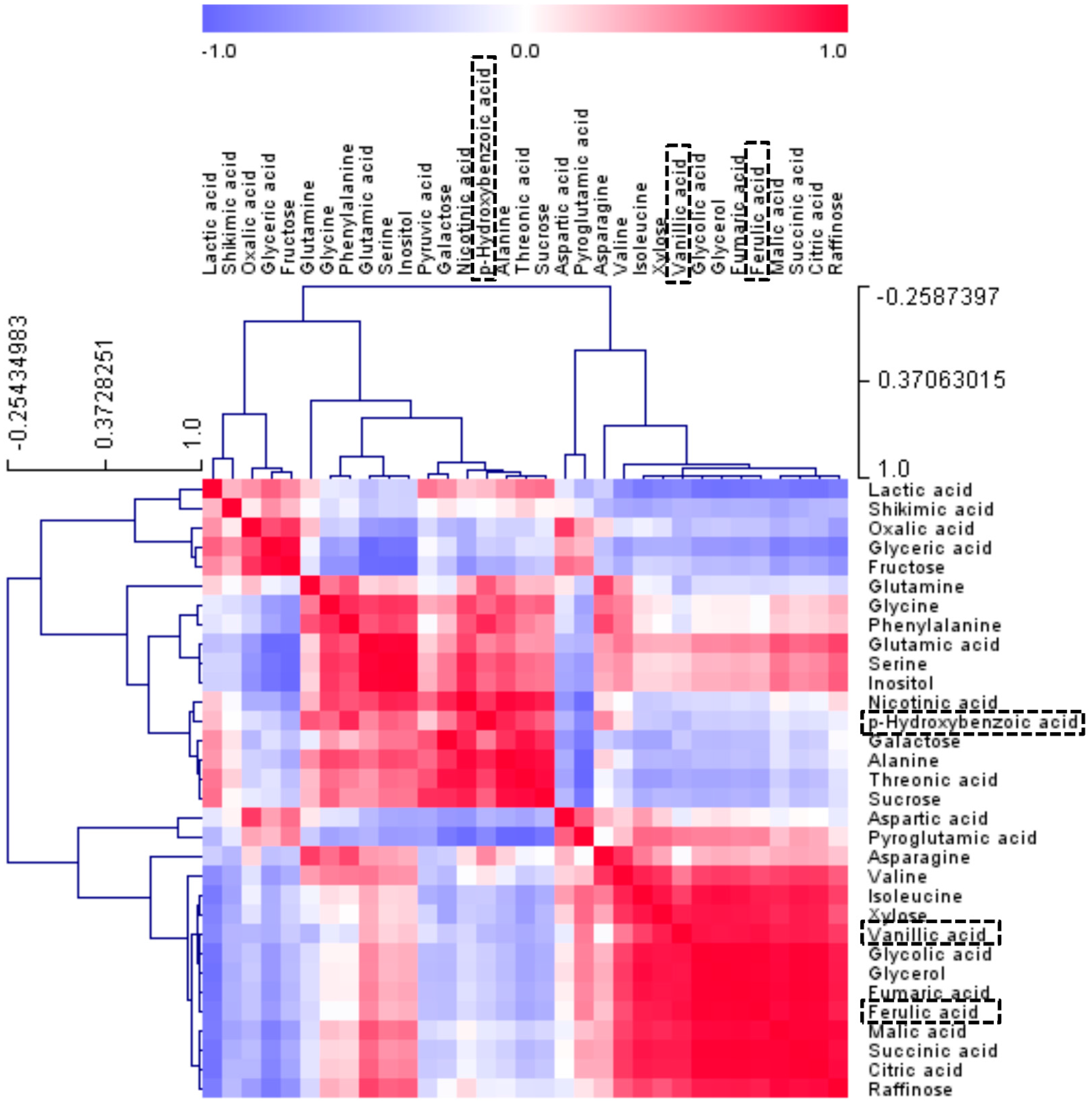
2.4. In-Vitro Antioxidant Assay
2.4.1. Superoxide Radical Scavenging Activity

2.4.2. DPPH Assay
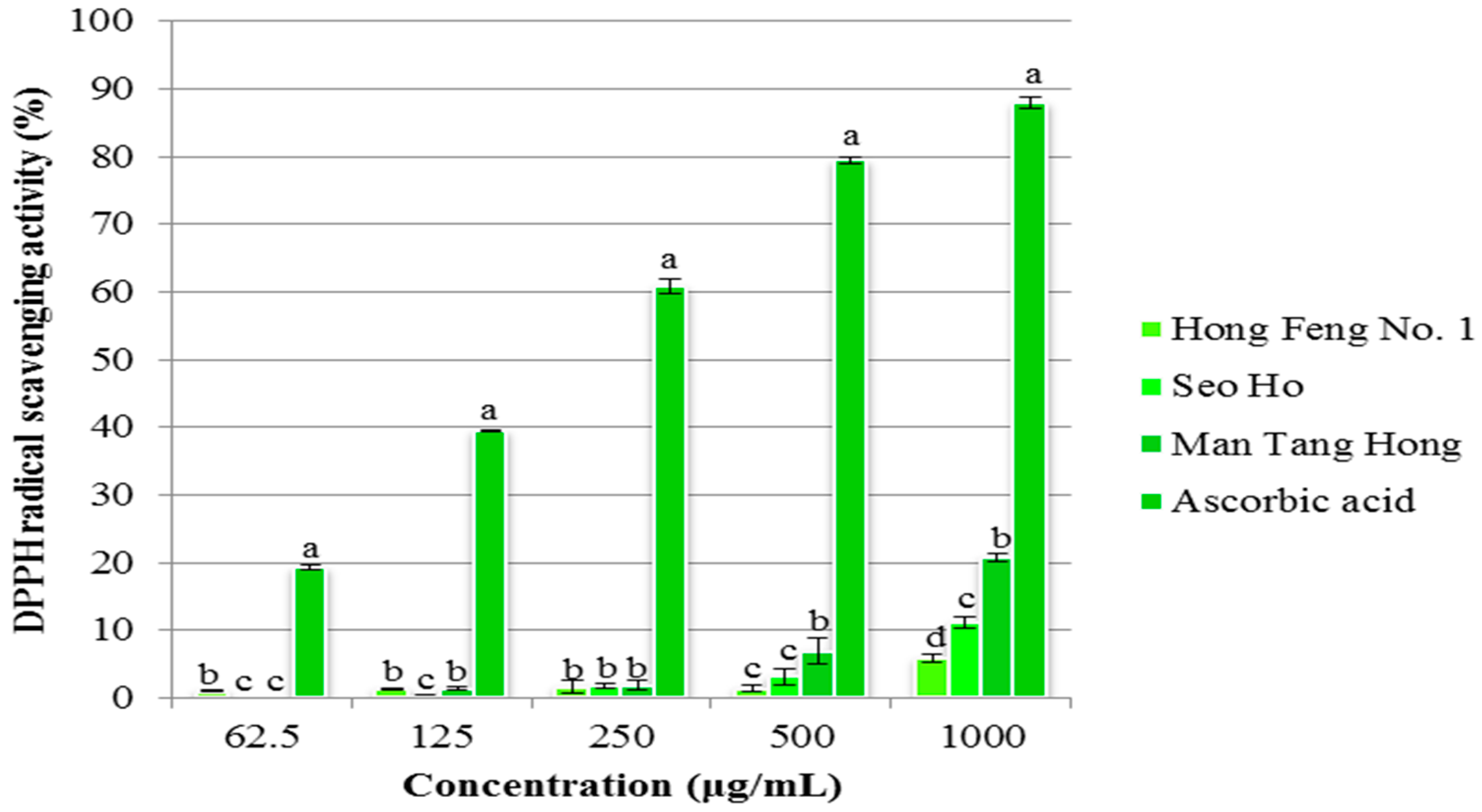
3. Experimental Section
3.1. Plant Materials
3.2. Anthocyanin Extraction and Analysis
3.3. GC-TOFMS Analysis of Polar Metabolites
3.4. In-Vitro Antioxidant Assay
3.4.1. Superoxide Radical Scavenging Activity
3.4.2. DPPH Assay
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ogra, Y.; Kitaguchi, T.; Ishiwata, K.; Suzuki, N.; Iwashita, Y.; Suzuki, K.T. Identification of selenohomolanthionine in selenium-enriched Japanese pungent radish. J. Anal. At. Spectrom. 2007, 22, 1390–1396. [Google Scholar] [CrossRef]
- UNT Digital Library. Avaliable online: http://digital.library.unt.edu/ark:/67531/metadc1562/ (accessed on 26 January 2016).
- Muminović, J.; Merz, A.; Melchinger, A.E.; Lübberstedt, T. Genetic structure and diversity among radish varieties as inferred from AFLP and ISSR analyses. J. Am. Soc. Hortic. Sci. 2005, 130, 79–87. [Google Scholar]
- Hanlon, P.R.; Barnes, D.M. Phytochemical composition and biological activity of 8 varieties of radish (Raphanus sativus L.) sprouts and mature taproots. J. Food Sci. 2011, 76, C185–C192. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathi, P.; Kirshnappa, K.; Chandre Gowda, M.; Shiva Reddy, N.; Anjanappa, M. Growth and yield of certain radish varieties to varying levels of fertility. Karnataka J. Agric. Sci. 1999, 12, 148–153. [Google Scholar]
- George, R.; Evans, D. A classification of winter radish cultivars. Euphytica 1981, 30, 483–492. [Google Scholar] [CrossRef]
- Giusti, M.M.; Rodríguez-Saona, L.E.; Baggett, J.R.; Reed, G.L.; Durst, R.W.; Wrolstad, R.E. Anthocyanin pigment composition of red radish cultivars as potential food colorants. J. Food Sci. 1998, 63. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Radish anthocyanin extract as a natural red colorant for maraschino cherries. J. Food Sci. 1996, 61, 688–694. [Google Scholar] [CrossRef]
- Ishikura, N.; Hayashi, K. Chromatographic separation and characterization of the component anthocyanins in radish root. Studies on anthocyanins, XXXVIII. Bot. Mag. Tokyo 1963, 76, 6–13. [Google Scholar] [CrossRef]
- Mazza, G. Anthocyanins and heart health. Ann. Ist. Super. Sanita 2007, 43, 369–374. [Google Scholar] [PubMed]
- Tanaka, Y.; Ohmiya, A. Seeing is believing: Engineering anthocyanin and carotenoid biosynthetic pathways. Curr. Opin. Biotechnol. 2008, 19, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Prior, R.L. Identification and characterization of anthocyanins by high-performance liquid chromatography-electrospray ionization-tandem mass spectrometry in common foods in the United States: Vegetables, nuts, and grains. J. Agric. Food Chem. 2005, 53, 3101–3113. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.M.; Ghanadan, H.; Wrolstad, R.E. Elucidation of the structure and conformation of red radish (Raphanus sativus) anthocyanins using one-and two-dimensional nuclear magnetic resonance techniques. J. Agric. Food Chem. 1998, 46, 4858–4863. [Google Scholar] [CrossRef]
- Harborne, J. Flavonoid sophorosides. Experientia. 1963, 19, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, R.M.P.; Perez, R.L. Raphanus sativus (Radish): Their chemistry and biology. Sci. World J. 2004, 4, 811–837. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-W.; Kim, E.-K.; Lee, S.-J.; Kim, Y.-S.; Choi, D.-K.; Park, T.-K.; Moon, S.-H.; Jeon, B.-T.; Park, P.-J. Anthocyanin effectively scavenges free radicals and protects retinal cells from H2O2-triggered G2/M arrest. Eur. Food Res. Technol. 2012, 234, 431–439. [Google Scholar] [CrossRef]
- Wang, L.-S.; Stoner, G.D. Anthocyanins and their role in cancer prevention. Cancer Lett. 2008, 269, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Wallace, T.C. Anthocyanins in cardiovascular disease. Adv. Nutr. Inter. Rev. J. 2011, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.S.; Jeon, D.S.; Park, S.G.; Arasu, M.V.; Al-Dhabi, N.A.; Kim, S.C.; Kim, S.J. Effect of cold storage on the contents of glucosinolates in Chinese cabbage (Brassica rapa L. ssp. pekinensis). South Ind. J. Biol. Sci. 2015, 1, 38–42. [Google Scholar]
- Ghosh, D.; Konishi, T. Anthocyanins and anthocyanin-rich extracts: Role in diabetes and eye function. Asia Pacific J. Clin. Nutr. 2007, 16, 200–208. [Google Scholar]
- Clarke, C.J.; Haselden, J.N. Metabolic profiling as a tool for understanding mechanisms of toxicity. Toxicol. Pathol. 2008, 36, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Schad, M.; Mungur, R.; Fiehn, O.; Kehr, J. Metabolic profiling of laser microdissected vascular bundles of Arabidopsis thaliana. Plant Methods 2005, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry–based metabolite profiling in plants. Nat. Protoc. Electron. Ed. 2006, 1, 387. [Google Scholar] [CrossRef] [PubMed]
- Roessner, U.; Luedemann, A.; Brust, D.; Fiehn, O.; Linke, T.; Willmitzer, L.; Fernie, A.R. Metabolic profiling allows comprehensive phenotyping of genetically or environmentally modified plant systems. Plant Cell 2001, 13, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Kopka, J.; Dörmann, P.; Altmann, T.; Trethewey, R.N.; Willmitzer, L. Metabolite profiling for plant functional genomics. Nat. Biotechnol. 2000, 18, 1157–1161. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.; Sefkow, M.; Kopka, J. Construction and application of a mass spectral and retention time index database generated from plant GC/EI-TOF-MS metabolite profiles. Phytochemistry 2003, 62, 887–900. [Google Scholar] [CrossRef]
- Kopka, J.; Fernie, A.; Weckwerth, W.; Gibon, Y.; Stitt, M. Metabolite profiling in plant biology: Platforms and destinations. Genome Biol. 2004, 5, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumner, L.W.; Mendes, P.; Dixon, R.A. Plant metabolomics: Large-scale phytochemistry in the functional genomics era. Phytochemistry 2003, 62, 817–836. [Google Scholar] [CrossRef]
- Ksiksi, T.; Hamza, A.A. Antioxidant, lipoxygenase and histone deacetylase inhibitory activities of Acridocarbus orientalis from al ain and oman. Molecules 2012, 17, 12521–12532. [Google Scholar] [CrossRef] [PubMed]
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A review of the antioxidant potential of medicinal plant species. Food Bioprod. Process. 2011, 89, 217–233. [Google Scholar] [CrossRef]
- García-Andrade, M.; González-Laredo, R.; Rocha-Guzmán, N.; Gallegos-Infante, J.; Rosales-Castro, M.; Medina-Torres, L. Mesquite leaves (Prosopis laevigata), a natural resource with antioxidant capacity and cardioprotection potential. Ind. Crops Prod. 2013, 44, 336–342. [Google Scholar] [CrossRef]
- Tsouvaltzis, P.; Brecht, J.K. Changes in Quality and Antioxidant Enzyme Activities of Bunched and Topped Radish (Raphanus sativus L.) Plants during Storage at 5 or 10C. J. Food Qual. 2014, 37, 157–167. [Google Scholar] [CrossRef]
- Takaya, Y.; Kondo, Y.; Furukawa, T.; Niwa, M. Antioxidant constituents of radish sprout (kaiware-daikon), Raphanus sativus L. J. Agric. Food Chem. 2003, 51, 8061–8066. [Google Scholar] [CrossRef] [PubMed]
- Beevi, S.S.; Narasu, M.L.; Gowda, B.B. Polyphenolics profile, antioxidant and radical scavenging activity of leaves and stem of Raphanus sativus L. Plant Food Human Nutr. 2010, 65, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Beevi, S.S.; Mangamoori, L.N.; Gowda, B.B. Polyphenolics profile and antioxidant properties of Raphanus sativus L. Nat. Prod. Res. 2012, 26, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Goyeneche, R.; Roura, S.; Ponce, A.; Vega-Gálvez, A.; Quispe-Fuentes, I.; Uribe, E.; Di Scala, K. Chemical characterization and antioxidant capacity of red radish (Raphanus sativus L.) leaves and roots. J. Funct. Foods 2015, 16, 256–264. [Google Scholar] [CrossRef]
- Jing, P.; Zhao, S.; Ruan, S.; Sui, Z.; Chen, L.; Jiang, L.; Qian, B. Quantitative studies on structure–ORAC relationships of anthocyanins from eggplant and radish using 3D-QSAR. Food Chem. 2014, 145, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Murakami, N.; Wang, L.; Zhang, S. Preparative high-performance liquid chromatography for the purification of natural acylated anthocyanins from red radish (Raphanus sativus L.). J. Chromatogr. Sci. 2008, 46, 743–746. [Google Scholar] [CrossRef] [PubMed]
- Matsufuji, H.; Otsuki, T.; Takeda, T.; Chino, M.; Takeda, M. Identification of reaction products of acylated anthocyanins from red radish with peroxyl radicals. J. Agric. Food Chem. 2003, 51, 3157–3161. [Google Scholar] [CrossRef] [PubMed]
- Park, N.I.; Xu, H.; Li, X.; Jang, I.H.; Park, S.; Ahn, G.H.; Lim, Y.P.; Kim, S.J.; Park, S.U. Anthocyanin accumulation and expression of anthocyanin biosynthetic genes in radish (Raphanus sativus). J. Agric. Food Chem. 2011, 59, 6034–6039. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Park, S.-Y.; Lim, S.-H.; Yeo, Y.; Cho, H.S.; Ha, S.-H. Comparative metabolic profiling of pigmented rice (Oryza sativa L.) cultivars reveals primary metabolites are correlated with secondary metabolites. J. Cereal Sci. 2013, 57, 14–20. [Google Scholar] [CrossRef]
- Joseph, M.A.; Moysich, K.B.; Freudenheim, J.L.; Shields, P.G.; Bowman, E.D.; Zhang, Y.; Marshall, J.R.; Ambrosone, C.B. Cruciferous vegetables, genetic polymorphisms in glutathione S-transferases M1 and T1, and prostate cancer risk. Nutr. Cancer 2004, 50, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.O.; Lee, I.S. Induction of quinone reductase, the phase 2 anticarcinogenic marker enzyme, in Hepa1c1c7 cells by radish sprouts, Raphanus sativus L. J. Food Sci. 2006, 71, S144–S148. [Google Scholar] [CrossRef]
- Higdon, J.V.; Delage, B.; Williams, D.E.; Dashwood, R.H. Cruciferous vegetables and human cancer risk: Epidemiologic evidence and mechanistic basis. Pharmacol. Res. 2007, 55, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Lima, G.P.P.; da Rocha, S.A.; Takaki, M.; Ramos, P.R.R.; Ono, E.O. Comparison of polyamine, phenol and flavonoid contents in plants grown under conventional and organic methods. Int. J. Food Sci. Technol. 2008, 43, 1838–1843. [Google Scholar] [CrossRef]
- Truong, T.; Baron-Dubourdieu, D.; Rougier, Y.; Guénel, P. Role of dietary iodine and cruciferous vegetables in thyroid cancer: A countrywide case–control study in New Caledonia. Cancer Causes Control 2010, 21, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Manchali, S.; Murthy, K.N.C.; Patil, B.S. Crucial facts about health benefits of popular cruciferous vegetables. J. Funct. Foods 2012, 4, 94–106. [Google Scholar] [CrossRef]
- Jing, P.; Song, L.-H.; Shen, S.-Q.; Zhao, S.-J.; Pang, J.; Qian, B.-J. Characterization of phytochemicals and antioxidant activities of red radish brines during lactic acid fermentation. Molecules 2014, 19, 9675–9688. [Google Scholar] [CrossRef] [PubMed]
- Pushkala, R.; Raghuram, P.K.; Srividya, N. Chitosan based powder coating technique to enhance phytochemicals and shelf life quality of radish shreds. Postharvest Biol. Technol. 2013, 86, 402–408. [Google Scholar] [CrossRef]
- Bruni, R.; Sacchetti, G. Factors affecting polyphenol biosynthesis in wild and field grown St. John’s Wort (Hypericum perforatum L. Hypericaceae/Guttiferae). Molecules 2009, 14, 682–725. [Google Scholar] [CrossRef] [PubMed]
- Giusti, M.M.; Wrolstad, R.E. Acylated anthocyanins from edible sources and their applications in food systems. Biochem. Eng. J. 2003, 14, 217–225. [Google Scholar] [CrossRef]
- Park, W.T.; Kim, Y.B.; Seo, J.M.; Kim, S.-J.; Chung, E.; Lee, J.-H.; Park, S.U. Accumulation of anthocyanin and associated gene expression in radish sprouts exposed to light and methyl jasmonate. J. Agric. Food Chem. 2013, 61, 4127–4132. [Google Scholar] [CrossRef] [PubMed]
- De Nicola, G.R.; Bagatta, M.; Pagnotta, E.; Angelino, D.; Gennari, L.; Ninfali, P.; Rollin, P.; Iori, R. Comparison of bioactive phytochemical content and release of isothiocyanates in selected brassica sprouts. Food Chem. 2013, 141, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Tatsuzawa, F.; Saito, N.; Toki, K.; Shinoda, K.; Shigihara, A.; Honda, T. Acylated Cyanidin 3-sophoroside-5-glucosides from the Purple Roots of Red Radish (Raphanus sativus L.) “Benikanmi”. J. Jpn. Soc. Hortic. Sci. 2010, 79, 103–107. [Google Scholar] [CrossRef]
- Matera, R.; Gabbanini, S.; de Nicola, G.R.; Iori, R.; Petrillo, G.; Valgimigli, L. Identification and analysis of isothiocyanates and new acylated anthocyanins in the juice of Raphanus sativus cv. Sango sprouts. Food Chem. 2012, 133, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-Z.; Sun, J.; Chen, P.; Harnly, J.A. LC-PDA-ESI/MSn identification of new anthocyanins in purple Bordeaux radish (Raphanus sativus L. variety). J. Agric. Food Chem. 2011, 59, 6616–6627. [Google Scholar] [CrossRef] [PubMed]
- Barillari, J.; Cervellati, R.; Paolini, M.; Tatibouët, A.; Rollin, P.; Iori, R. Isolation of 4-methylthio-3-butenyl glucosinolate from Raphanus sativus sprouts (Kaiware Daikon) and its redox properties. J. Agric. Food Chem. 2005, 53, 9890–9896. [Google Scholar] [CrossRef] [PubMed]
- Barillari, J.; Cervellati, R.; Costa, S.; Guerra, M.C.; Speroni, E.; Utan, A.; Iori, R. Antioxidant and choleretic properties of Raphanus sativus L. sprout (Kaiware Daikon) extract. J. Agric. Food Chem. 2006, 54, 9773–9778. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, D.T.; Goldbohm, R.A.; van Poppel, G.; Verhagen, H.; van den Brandt, P.A. Epidemiological studies on brassica vegetables and cancer risk. Cancer Epidemiol. Biomark. Prev. 1996, 5, 733–748. [Google Scholar]
- Naczk, M.; Shahidi, F. Extraction and analysis of phenolics in food. J. Chromatogr. A 2004, 1054, 95–111. [Google Scholar] [CrossRef]
- Ross, K.; Beta, T.; Arntfield, S. A comparative study on the phenolic acids identified and quantified in dry beans using HPLC as affected by different extraction and hydrolysis methods. Food Chem. 2009, 113, 336–344. [Google Scholar] [CrossRef]
- Pérez-Balibrea, S.; Moreno, D.A.; García-Viguera, C. Genotypic effects on the phytochemical quality of seeds and sprouts from commercial broccoli cultivars. Food Chem. 2011, 125, 348–354. [Google Scholar] [CrossRef]
- Muhlemann, J.K.; Maeda, H.; Chang, C.-Y.; San Miguel, P.; Baxter, I.; Cooper, B.; Perera, M.A.; Nikolau, B.J.; Vitek, O.; Morgan, J.A. Developmental changes in the metabolic network of snapdragon flowers. PLoS ONE 2012, 7, e40381. [Google Scholar] [CrossRef] [PubMed]
- Lancien, M.; Gadal, P.; Hodges, M. Enzyme redundancy and the importance of 2-oxoglutarate in higher plant ammonium assimilation. Plant Physiol. 2000, 123, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.R.; Krapp, A.; Stitt, M. Reciprocal diurnal changes of phosphoenolpyruvate carboxylase expression and cytosolic pyruvate kinase, citrate synthase and NADP-isocitrate dehydrogenase expression regulate organic acid metabolism during nitrate assimilation in tobacco leaves. Plant Cell Environ. 2000, 23, 1155–1167. [Google Scholar] [CrossRef]
- Martens, S.; Forkmann, G.; Matern, U.; Lukačin, R. Cloning of parsley flavone synthase I. Phytochemistry 2001, 58, 43–46. [Google Scholar] [CrossRef]
- Turnbull, J.J.; Nakajima, J.-I.; Welford, R.W.; Yamazaki, M.; Saito, K.; Schofield, C.J. Mechanistic studies on three 2-oxoglutarate-dependent oxygenases of flavonoid biosynthesis anthocyanidin synthase, flavonol synthase, and flavanone 3β-hydroxylase. J. Biol. Chem. 2004, 279, 1206–1216. [Google Scholar] [CrossRef] [PubMed]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. The evolution of phenylpropanoid metabolism in the green lineage. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 123–152. [Google Scholar] [CrossRef] [PubMed]
- Karaca, H.; Velioglu, Y.S. Effects of ozone treatments on microbial quality and some chemical properties of lettuce, spinach, and parsley. Postharvest Biol. Tech. 2014, 88, 46–53. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kang, O.-J.; Gweon, O.-C. Comparison of phenolic acids and flavonoids in black garlic at different thermal processing steps. J. Funct. Foods 2013, 5, 80–86. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.N.; Mahmoud, A.E.; Abdou, H.M. Quantification of Total Phenolic and Total Flavonoid Contents in Extracts of Some Egyptian Green leaves and Estimation of Antioxidant Activity. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 177–179. [Google Scholar]
- Katsuzaki, H.; Miyahara, Y.; Ota, M.; Imai, K.; Komiya, T. Chemistry and antioxidative activity of hot water extract of Japanese radish (daikon). Biofactors 2004, 21, 211–214. [Google Scholar] [CrossRef]
- Giusti, M.M.; Rodríguez-Saona, L.E.; Griffin, D.; Wrolstad, R.E. Electrospray and tandem mass spectroscopy as tools for anthocyanin characterization. J. Agric. Food Chem. 1999, 47, 4657–4664. [Google Scholar] [CrossRef] [PubMed]
- Tatsuzawa, F.; Toki, K.; Saito, N.; Shinoda, K.; Shigihara, A.; Honda, T. Anthocyanin occurrence in the root peels, petioles and flowers of red radish (Raphanus sativus L.). Dyes Pigments 2008, 79, 83–88. [Google Scholar] [CrossRef]
- Sunil, C.; Irudayaraj, S.S.; Duraipandiyan, V.; Al-Dhabi, N.A.; Agastian, P.; Ignacimuthu, S. Antioxidant and free radical scavenging effects of β-amyrin isolated from S. cochinchinensis Moore. leaves. Ind. Crops Prod. 2014, 61, 510–516. [Google Scholar] [CrossRef]
- Hatano, T.; Kagawa, H.; Yasuhara, T.; Okuda, T. Two new flavonoids and other constituents in licorice root. Their relative astringency and radical scavenging effects. Chem. Pharm. Bull. 1988, 36, 2090–2097. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Not available.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.H.; Baskar, T.B.; Park, S.-Y.; Kim, S.-J.; Valan Arasu, M.; Al-Dhabi, N.A.; Kim, J.K.; Park, S.U. Metabolic Profiling and Antioxidant Assay of Metabolites from Three Radish Cultivars (Raphanus sativus). Molecules 2016, 21, 157. https://doi.org/10.3390/molecules21020157
Park CH, Baskar TB, Park S-Y, Kim S-J, Valan Arasu M, Al-Dhabi NA, Kim JK, Park SU. Metabolic Profiling and Antioxidant Assay of Metabolites from Three Radish Cultivars (Raphanus sativus). Molecules. 2016; 21(2):157. https://doi.org/10.3390/molecules21020157
Chicago/Turabian StylePark, Chang Ha, Thanislas Bastin Baskar, Soo-Yun Park, Sun-Ju Kim, Mariadhas Valan Arasu, Naif Abdullah Al-Dhabi, Jae Kwang Kim, and Sang Un Park. 2016. "Metabolic Profiling and Antioxidant Assay of Metabolites from Three Radish Cultivars (Raphanus sativus)" Molecules 21, no. 2: 157. https://doi.org/10.3390/molecules21020157