
Molecular Mechanism of the Flavonoid Natural Product Dryocrassin ABBA against Staphylococcus aureus Sortase A


Abstract
:1. Introduction
2. Results and Discussion
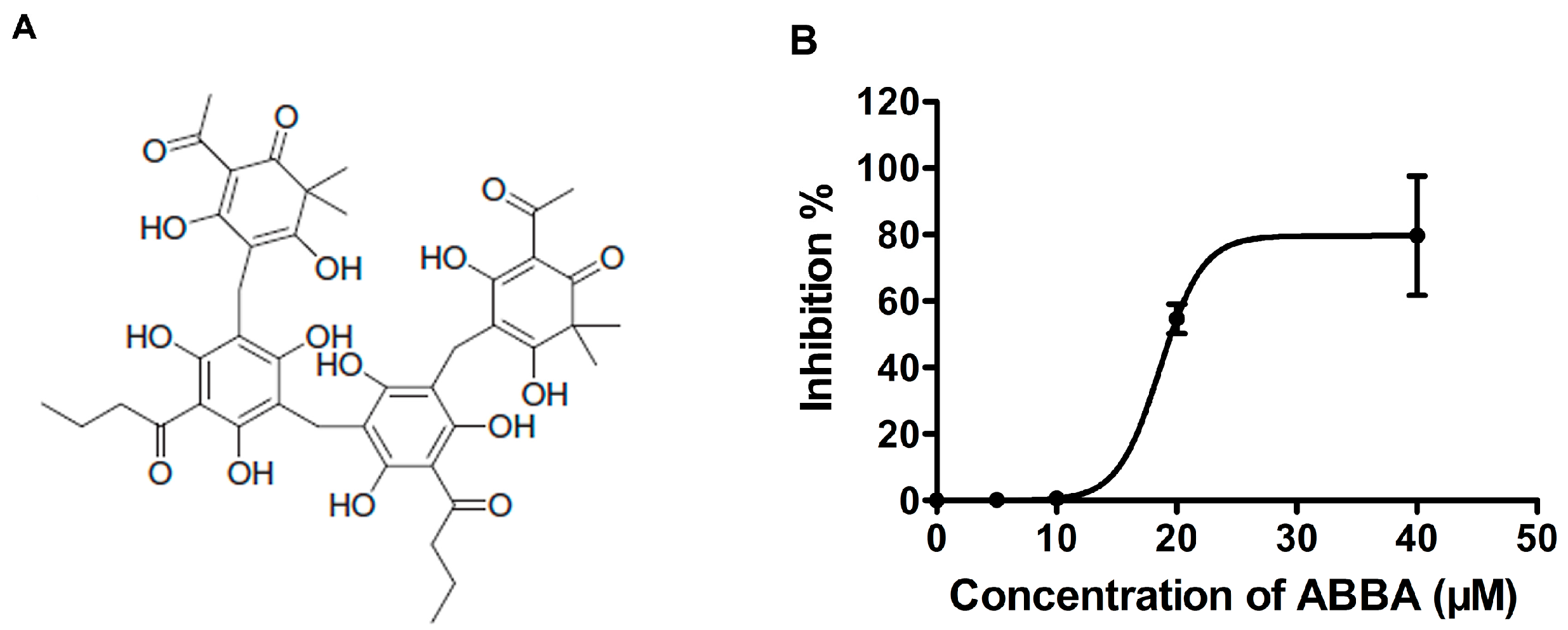
2.1. ABBA Inhibits the Activity of S. aureus SrtA
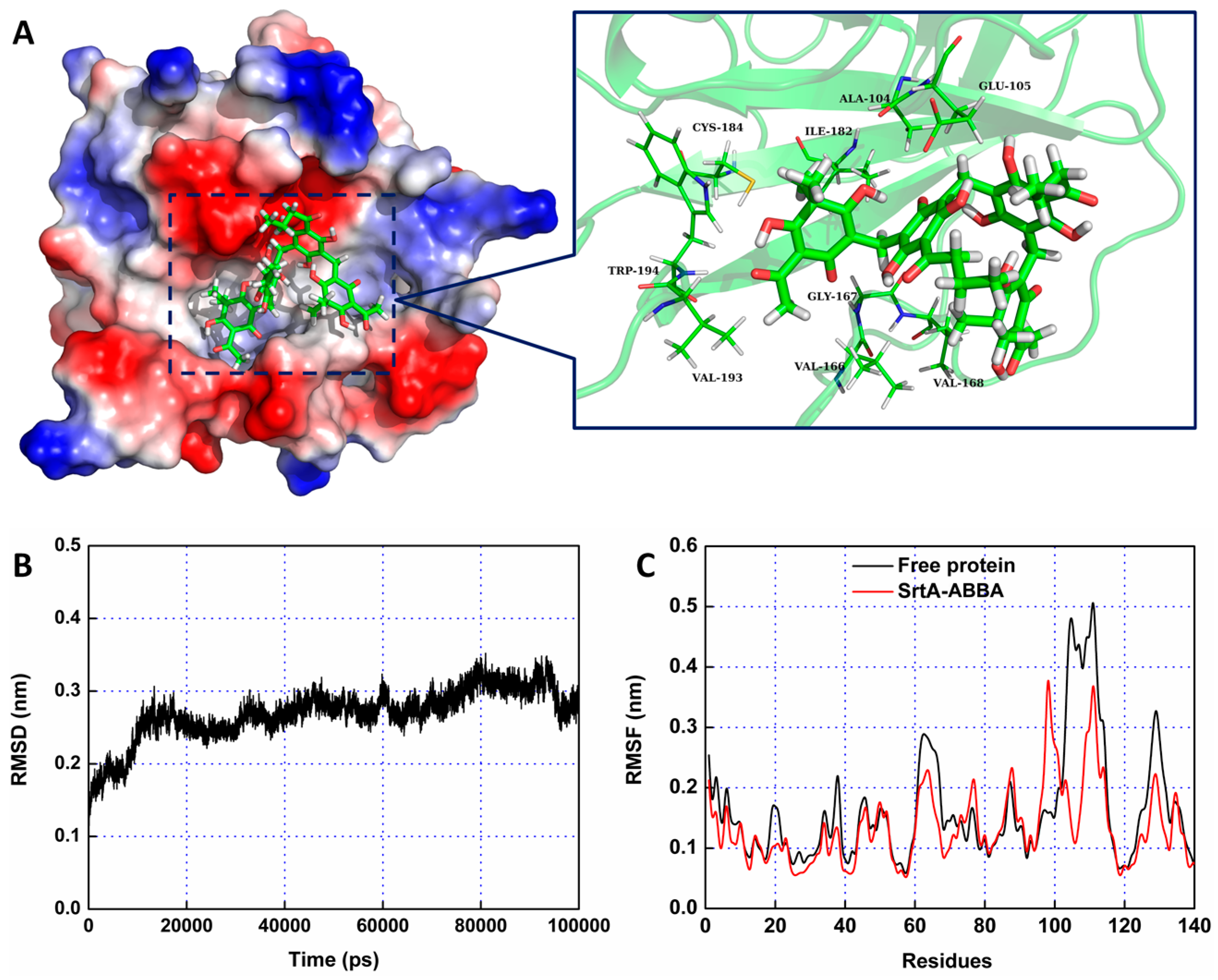
2.2. Molecular Dynamics Simulation for SrtA-ABBA
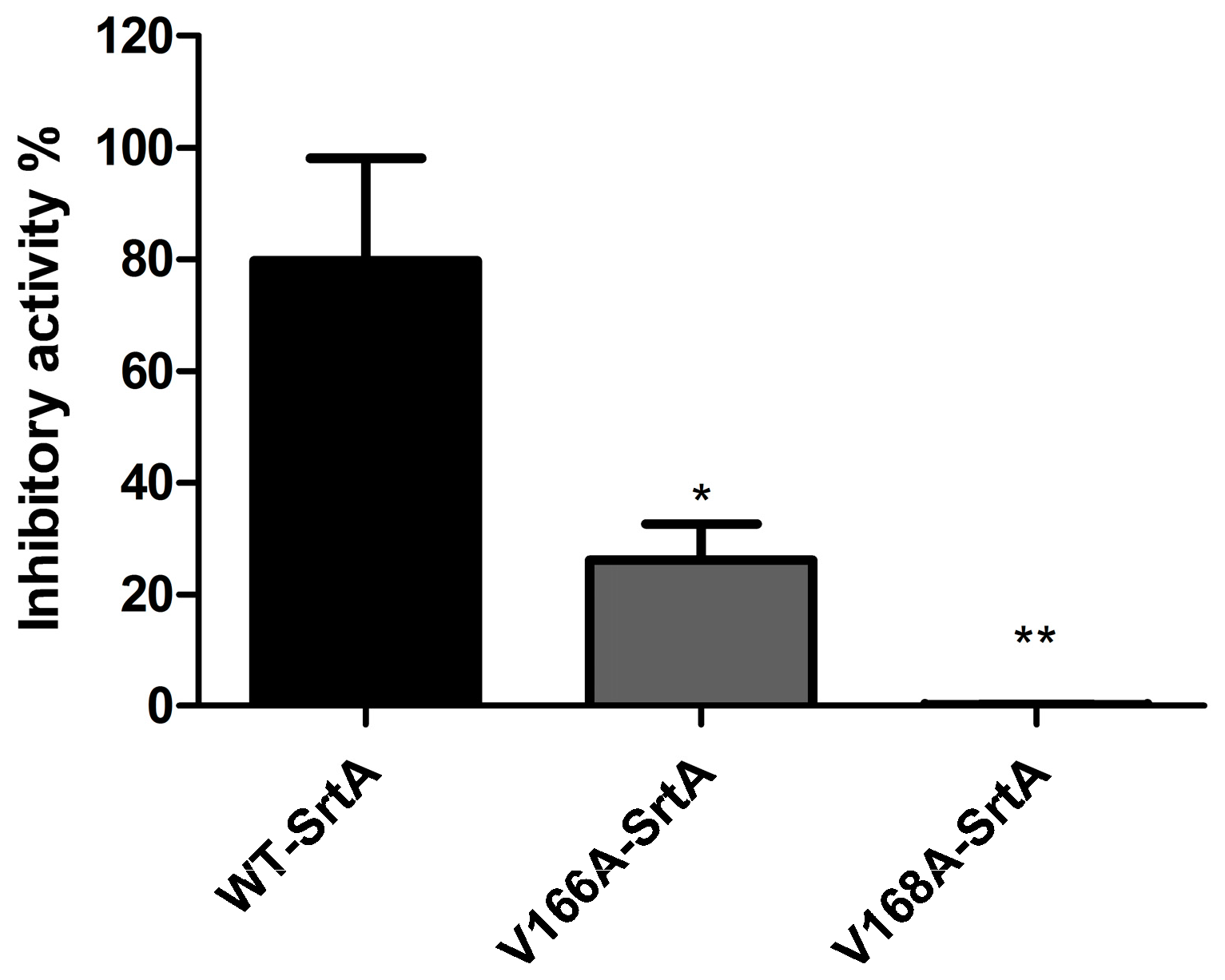
2.3. Validation of the Molecular Basis of ABBA against SrtA
3. Experimental Methods
3.1. Bacterial Strains, Growth and Reagents
3.2. Construction of Plasmids Encoding Wild-Type (WT)-SrtA, V166A-SrtA, V168A-SrtA
3.3. Expression and Purification of WT-SrtA, V166A-SrtA and V168A-SrtA
3.4. Determination of Mutant and WT SrtA Activity
3.5. Molecular Modeling
3.6. Binding Affinity Determination of ABBA with SrtA
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Deurenberg, R.H.; Stobberingh, E.E. The evolution of Staphylococcus aureus. Infect. Genet. Evol. 2008, 8, 747–763. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.Y.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G., Jr. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.L. Epidemiology of drug resistance: Implications for a post-antimicrobial era. Science 1992, 257, 1050–1055. [Google Scholar] [CrossRef] [PubMed]
- Tomasz, A. Multiple-antibiotic-resistant pathogenic bacteria. A report on the Rockefeller University Workshop. N. Engl. J. Med. 1994, 330, 1247–1251. [Google Scholar] [CrossRef] [PubMed]
- Swartz, M.N. Use of antimicrobial agents and drug resistance. N. Engl. J. Med. 1997, 337, 491–492. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, X.; Li, W.; Deng, X.; Deng, Y.; Niu, X. Curcumin protects mice from Staphylococcus aureus pneumonia by interfering with the self-assembly process of alpha-hemolysin. Sci. Rep. 2016, 6, 28254. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Qiu, J.; Tan, W.; Zhang, Y.; Wang, H.; Zhou, X.; Liu, S.; Feng, H.; Li, W.; Niu, X.; et al. Fisetin inhibits listeria monocytogenes virulence by interfering with the oligomerization of listeriolysin o. J. Infect. Dis. 2015, 211, 1376–1387. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Chang, I.M.; Oh, K.B. Inhibition of the bacterial surface protein anchoring transpeptidase sortase by medicinal plants. Biosci. Biotechnol. Biochem. 2002, 66, 2751–2754. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, H.; Wang, L.; Song, Z.; Shi, L.; Li, W.; Deng, X.; Wang, J. Isorhamnetin attenuates Staphylococcus aureus-induced lung cell injury by inhibiting α-hemolysin expression. J. Microbiol. Biotechnol. 2016, 26, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.R.; Barnett, T.C. Surface proteins of Gram-positive bacteria and how they get there. Annu. Rev. Microbiol. 2006, 60, 397–423. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Liu, G.; Hung, T.T.; Schneewind, O. Staphylococcus aureus sortase, an enzyme that anchors surface proteins to the cell wall. Science 1999, 285, 760–763. [Google Scholar] [CrossRef] [PubMed]
- Schneewind, O.; Model, P.; Fischetti, V.A. Sorting of protein a to the staphylococcal cell wall. Cell 1992, 70, 267–281. [Google Scholar] [CrossRef]
- Schneewind, O.; Mihaylova-Petkov, D.; Model, P. Cell wall sorting signals in surface proteins of Gram-positive bacteria. EMBO 1993, 12, 4803–4811. [Google Scholar]
- Navarre, W.W.; Schneewind, O. Proteolytic cleavage and cell wall anchoring at the LPXTG motif of surface proteins in Gram-positive bacteria. Mol. Microbiol. 1994, 14, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Schneewind, O.; Fowler, A.; Faull, K.F. Structure of the cell wall anchor of surface proteins in Staphylococcus aureus. Science 1995, 268, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Ton-That, H.; Faull, K.F.; Schneewind, O. Anchor structure of staphylococcal surface proteins. A branched peptide that links the carboxyl terminus of proteins to the cell wall. J. Biol. Chem. 1997, 272, 22285–22292. [Google Scholar] [CrossRef] [PubMed]
- Navarre, W.W.; Ton-That, H.; Faull, K.F.; Schneewind, O. Anchor structure of staphylococcal surface proteins. II. COOH-terminal structure of muramidase and amidase-solubilized surface protein. J. Biol. Chem. 1998, 273, 29135–29142. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.X.; Eriksson, L.A. Catalytic mechanism and roles of Arg197 and Thr183 in the Staphylococcus aureus sortase a enzyme. J. Phys. Chem. B 2011, 115, 13003–13011. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Liu, G.; Jensen, E.R.; Lenoy, E.; Schneewind, O. Staphylococcus aureus sortase mutants defective in the display of surface proteins and in the pathogenesis of animal infections. Proc. Natl. Acad. Sci. USA 2000, 97, 5510–5515. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.G.; Kim, H.K.; Burts, M.L.; Krausz, T.; Schneewind, O.; Missiakas, D.M. Genetic requirements for Staphylococcus aureus abscess formation and persistence in host tissues. FASEB J. 2009, 23, 3393–3404. [Google Scholar] [CrossRef] [PubMed]
- McAdow, M.; Kim, H.K.; Dedent, A.C.; Hendrickx, A.P.; Schneewind, O.; Missiakas, D.M. Preventing Staphylococcus aureus sepsis through the inhibition of its agglutination in blood. PLoS Pathog. 2011, 7, e1002307. [Google Scholar] [CrossRef] [PubMed]
- Moellering, R.C., Jr. Discovering new antimicrobial agents. Int. J. Antimicrob. Agents 2011, 37, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Ou, C.; Zhang, Q.; Wu, G.; Shi, N.; He, C. Dryocrassin ABBA, a novel active substance for use against amantadine-resistant H5N1 avian influenza virus. Front. Microbiol. 2015, 6, 592. [Google Scholar] [CrossRef] [PubMed]
- Maresso, A.W.; Schneewind, O. Sortase as a target of anti-infective therapy. Pharmacol. Rev. 2008, 60, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.; Bayley, H. Key residues for membrane binding, oligomerization, and pore forming activity of staphylococcal alpha-hemolysin identified by cysteine scanning mutagenesis and targeted chemical modification. J. Biol. Chem. 1995, 270, 23065–23071. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Hobaugh, M.R.; Shustak, C.; Cheley, S.; Bayley, H.; Gouaux, J.E. Structure of staphylococcal alpha-hemolysin, a heptameric transmembrane pore. Science 1996, 274, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Ton-That, H.; Liu, G.; Mazmanian, S.K.; Faull, K.F.; Schneewind, O. Purification and characterization of sortase, the transpeptidase that cleaves surface proteins of Staphylococcus aureus at the LPXTG motif. Proc. Natl. Acad. Sci. USA 1999, 96, 12424–12429. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Qiu, J.; Zhang, Y.; Lu, C.; Dai, X.; Wang, J.; Li, H.; Wang, X.; Tan, W.; Luo, M.; et al. Oroxylin a inhibits hemolysis via hindering the self-assembly of α-hemolysin heptameric transmembrane pore. PLoS Comput. Biol. 2013, 9, e1002869. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Qiu, J.; Wang, X.; Gao, X.; Dong, J.; Wang, J.; Li, H.; Zhang, Y.; Dai, X.; Lu, C.; et al. Molecular insight into the inhibition mechanism of cyrtominetin to alpha-hemolysin by molecular dynamics simulation. Eur. J. Med. Chem. 2013, 62, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Wang, H.S.; Niu, X.D. Molecular dynamics simulations reveal insight into key structural elements of aaptamines as sortase inhibitors with free energy calculations. Chem. Phys. Lett. 2013, 585, 171–177. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, X.; Liu, S.; Li, G.; Shi, L.; Dong, J.; Li, W.; Deng, X.; Niu, X. Morin hydrate attenuates Staphylococcus aureus virulence by inhibiting the self-assembly of alpha-hemolysin. Eur. J. Appl. Microbiol. 2015, 118, 753–763. [Google Scholar] [CrossRef] [PubMed]
- Jurasekova, Z.; Marconi, G.; Sanchez-Cortes, S.; Torreggiani, A. Spectroscopic and molecular modeling studies on the binding of the flavonoid luteolin and human serum albumin. Biopolymers 2009, 91, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, S.; Valder, C.R.; Huynh, H.G.; Ren, H.; Allison, W.S. The βG156c substitution in the A1-ATPase from the thermophilic bacillus PS3 affects catalytic site cooperativity by destabilizing the closed conformation of the catalytic site. Biochemistry 2002, 41, 14421–14429. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds screened in this study are available from the authors.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | WT-SrtA | V166A | V168A |
|---|---|---|---|
| The binding energy | −25.8 ± 2.8 | −19.2 ± 3.1 | −15.3 ± 1.9 |
| KA (1 × 104) L·mol−1 | 45.78 ± 6.5 | 43.5 ± 7.3 | 35.8 ± 5.5 |
| Primer Name | Oligonucleotide (5–3) a |
|---|---|
| WT-SrtA-F | GCGGGATCCCAAGCTAAACCTCAAATTCC |
| WT-SrtA-R | CCGCTCGAGTTATTTGACTTCTGTAGCTACAA |
| V166A-SrtA-F | GTTAAGCCTACAGATGCGGGAGTTCTAGATGAAC |
| V166A-SrtA-R | GTTCATCTAGAACTCCCGCATCTGTAGGCTTAAC |
| V168A-SrtA-F | CCTACAGATGTAGGAGCGCTAGATGAACAAAAAGG |
| V168A-SrtA-R | CCTTTTTGTTCATCTAGCGCTCCTACATCTGTAGG |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, B.; Wang, X.; Wang, L.; Chen, S.; Shi, D.; Wang, H. Molecular Mechanism of the Flavonoid Natural Product Dryocrassin ABBA against Staphylococcus aureus Sortase A. Molecules 2016, 21, 1428. https://doi.org/10.3390/molecules21111428
Zhang B, Wang X, Wang L, Chen S, Shi D, Wang H. Molecular Mechanism of the Flavonoid Natural Product Dryocrassin ABBA against Staphylococcus aureus Sortase A. Molecules. 2016; 21(11):1428. https://doi.org/10.3390/molecules21111428
Chicago/Turabian StyleZhang, Bing, Xiyan Wang, Lin Wang, Shuiye Chen, Dongxue Shi, and Hongsu Wang. 2016. "Molecular Mechanism of the Flavonoid Natural Product Dryocrassin ABBA against Staphylococcus aureus Sortase A" Molecules 21, no. 11: 1428. https://doi.org/10.3390/molecules21111428