1. Introduction
Ganoderma (Ganodermaceae) is a genus of edible and medicinal mushroom that is known as “reishi” in Japanese and “lingzhi” in Chinese [
1]. The medicinal values of this mushroom were documented thousands of years ago [
1]. Among the species of the genus,
G. lucidum (Leyss. ex Fr.) Karst was originally described as coming from Europe and is now known in East Asia, Europe and North America. Recently, a new species
G. lingzhi Sheng H. Wu, Y. Cao & Y.C. Dai, which is distributed in East Asia, was proposed. The most striking characteristics that differentiate
G. lingzhi from
G. lucidum are the presence of 1–2 black melanoid bands in the context, a yellow pore surface and thick dissepiments (80–120 μm) at maturity (in contrast,
G. lucidum has a white pore surface and thin dissepiments (40–80 μm)) [
2]. Modern chemistry has demonstrated that
Ganoderma species contain a variety of phytochemicals. Two biologically active compounds that have been demonstrated to possess diverse and potentially significant pharmacological activities are polysaccharides and triterpenes (especially triterpene acids, TA) [
1,
3,
4,
5,
6,
7,
8,
9,
10,
11,
12,
13,
14,
15,
16]. Recent studies have revealed that
Ganoderma TA has a few important pharmacological activities that include antitumor effects, inhibition of cholesterol synthesis, anti-HIV-1 and anti-HIV-protease activities, and inhibition of U46619-induced platelet aggregation [
5,
6,
7,
8,
9,
10].
Currently, the submerged fermentation of
Ganoderma is viewed as a promising approach for the efficient production of these polysaccharides and triterpenes because it is fast, cost-effective, and it is easy to control the product quality [
11,
12,
13,
14,
15,
16,
17,
18,
19]. Many researchers have focused on the fermentation conditions and fermentation modes to stimulate the production of
Ganoderma polysaccharides [
11,
12,
13,
14,
15]. These polysaccharides have been used in traditional medicine as antitumor and immunomodulating agents [
3,
20]. In contrast, the triterpenes or TA products of
Ganoderma are scarce on the international market, especially the pure individual triterpene compounds because the triterpene fermentation yield is very low and the required fermentation process is complex and not suitable for large-scale production [
10,
21]. Currently, few groups have studied triterpenes and specifically examined the production of ganoderic acids (a class of lanostane-type triterpenes) by
G. lucidum [
16,
17,
18,
19], but data on the submerged fermentation of TA with other
Ganoderma species are relatively scarce. Therefore, to meet the demand for TA for large-scale production, pharmacological testing and clinical applications, further study of TA production by
Ganoderma is expected to improve the operability of the industrial application by a simple and feasible fermentation technology. Additionally, there are more than 100 kinds of triterpene compounds isolated from
Ganoderma fruiting bodies, spores and mycelium (especially fruiting bodies). However, the constituents of the triterpenes from the three different sources are different, and most of the triterpenes or TAs with biological activities are isolated from the fruiting bodies of
Ganoderma [
4,
22,
23,
24,
25,
26,
27,
28,
29], and whether the triterpenes or TAs prepared from submerged culture
Ganoderma (especially
G. lingzhi) mycelium possesses antitumor activity needs to be further proved.
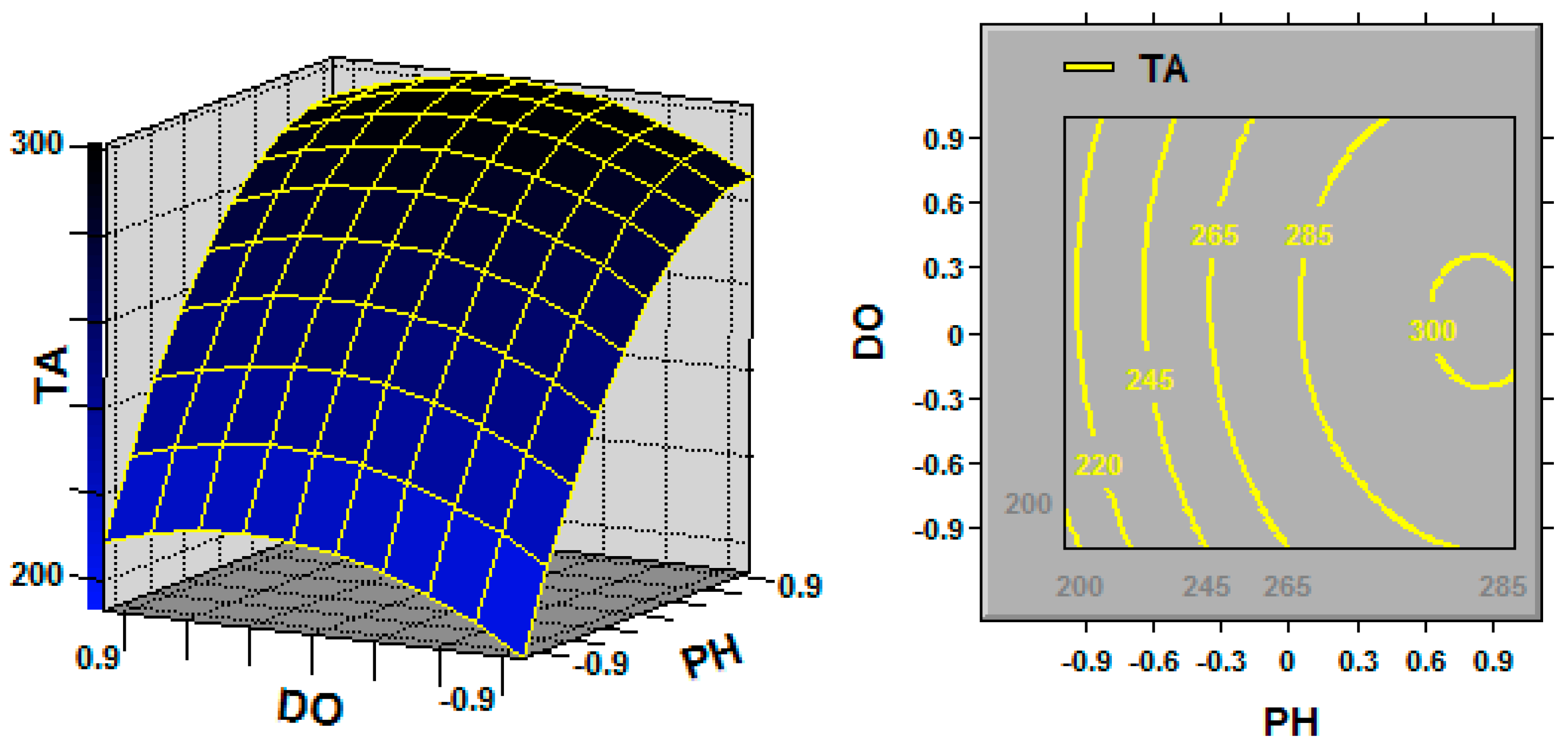
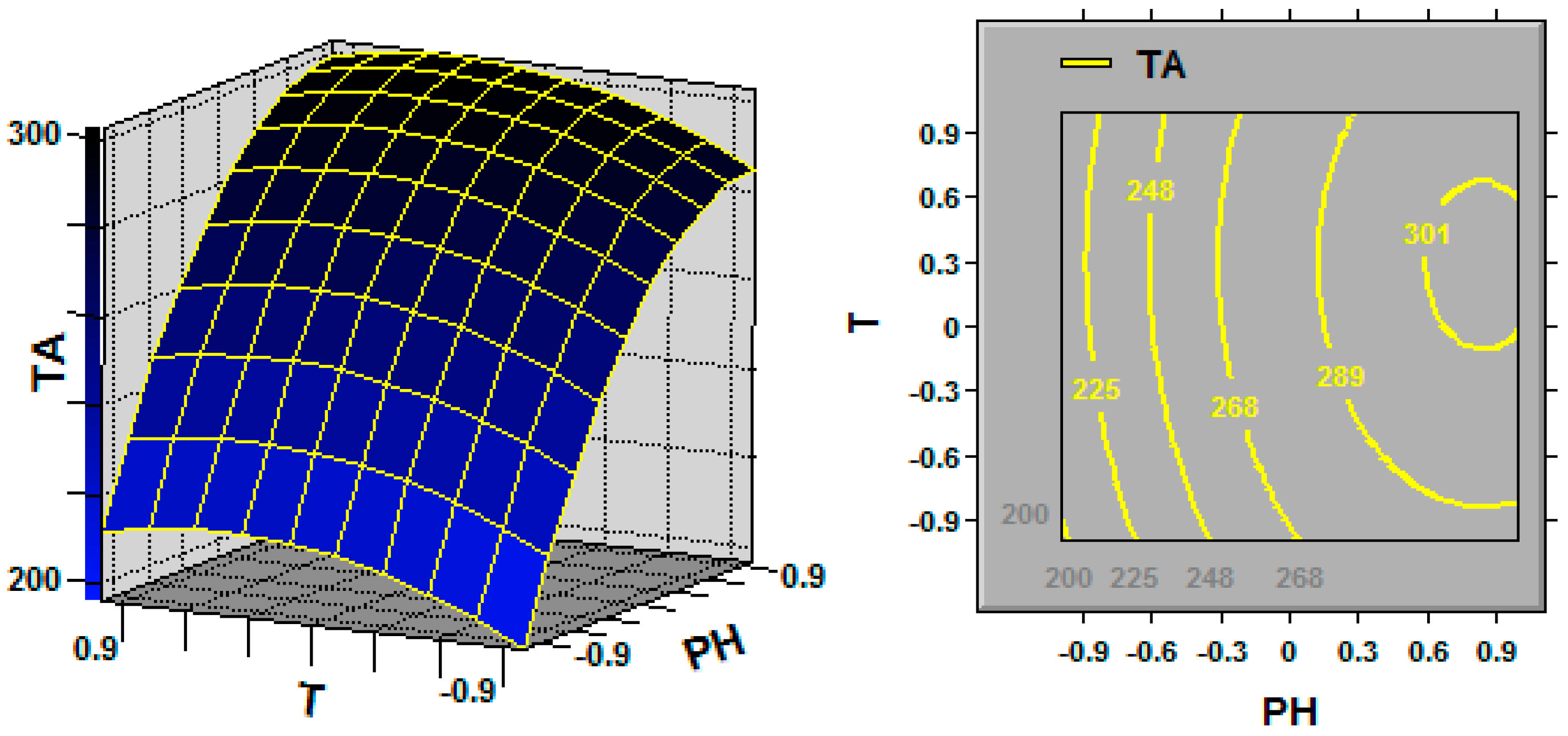
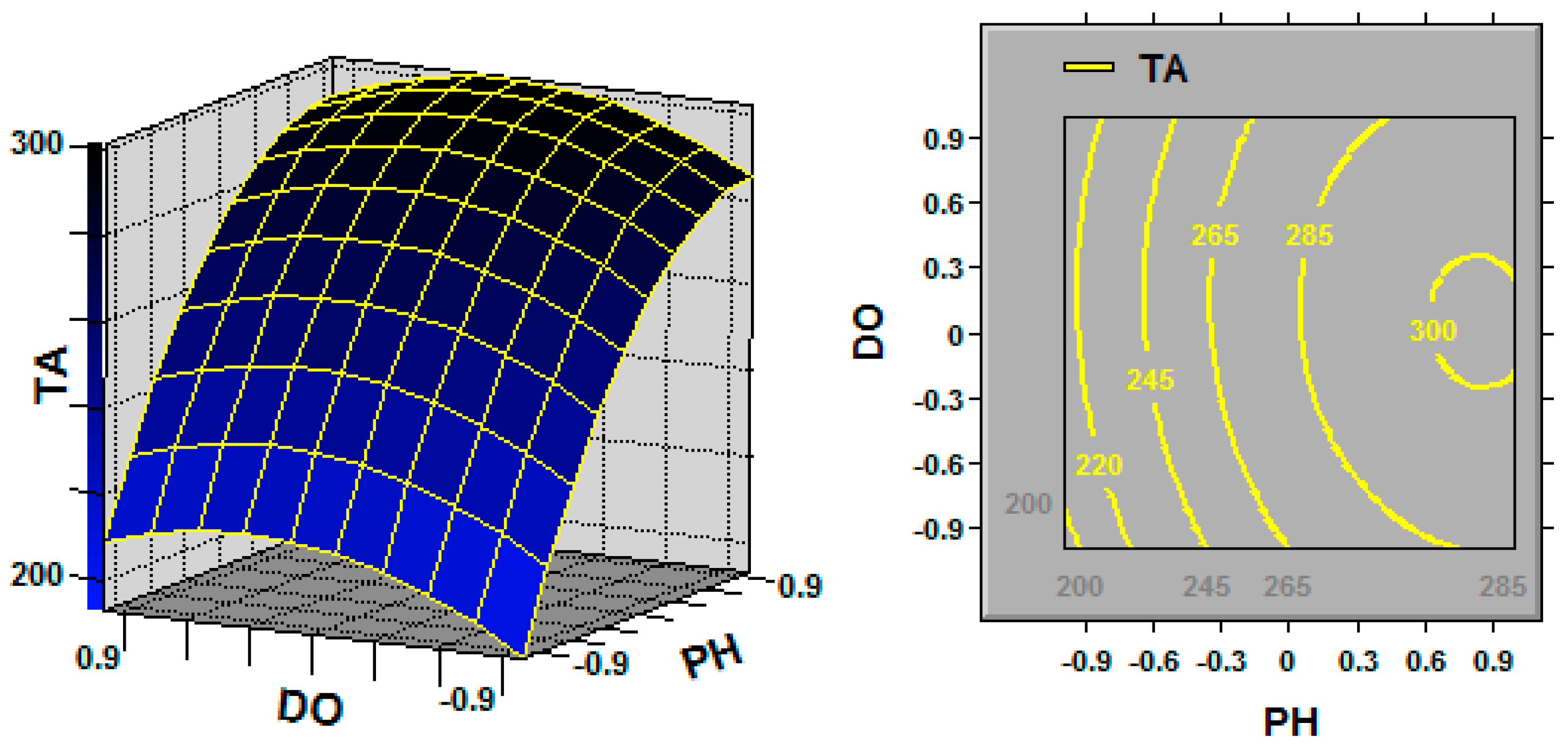
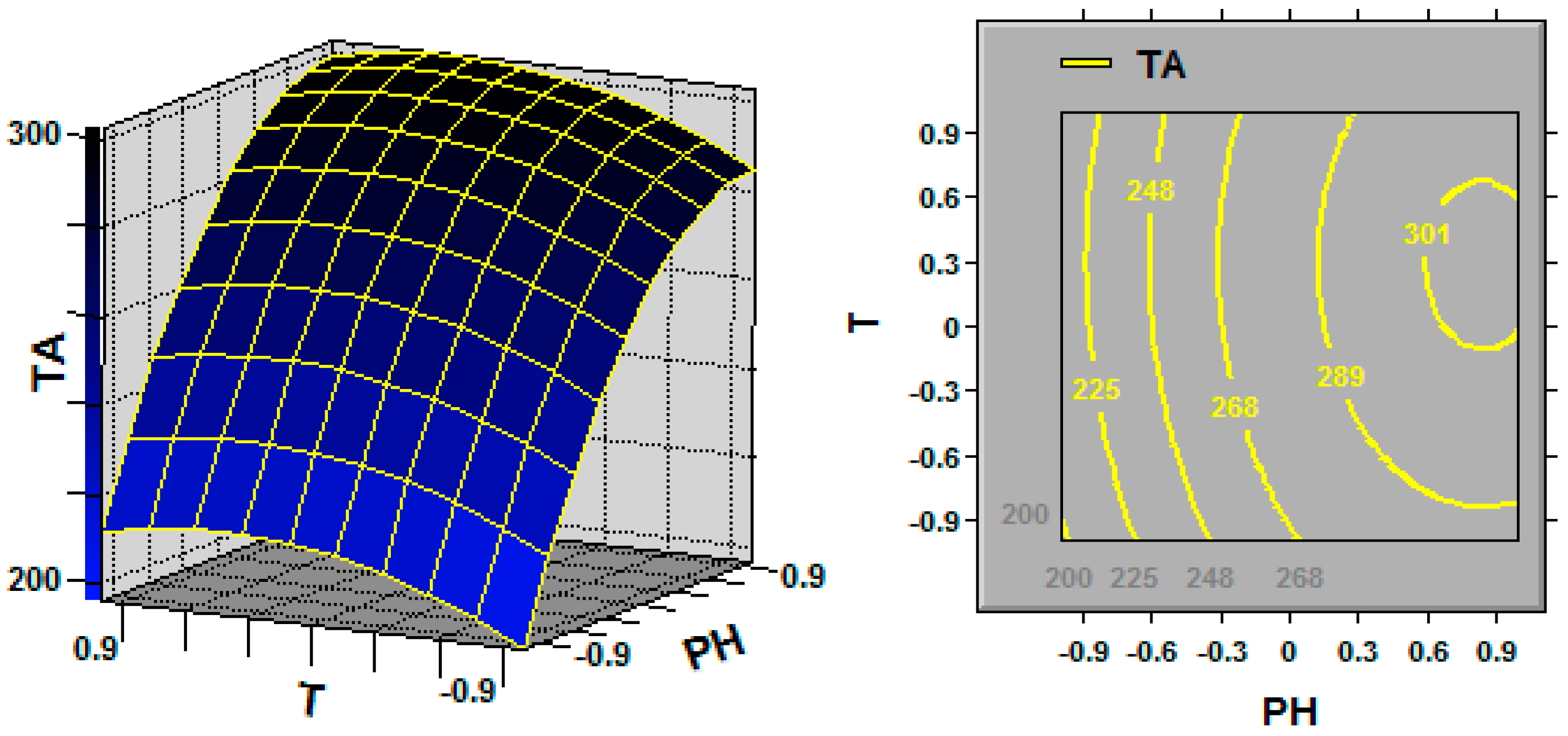
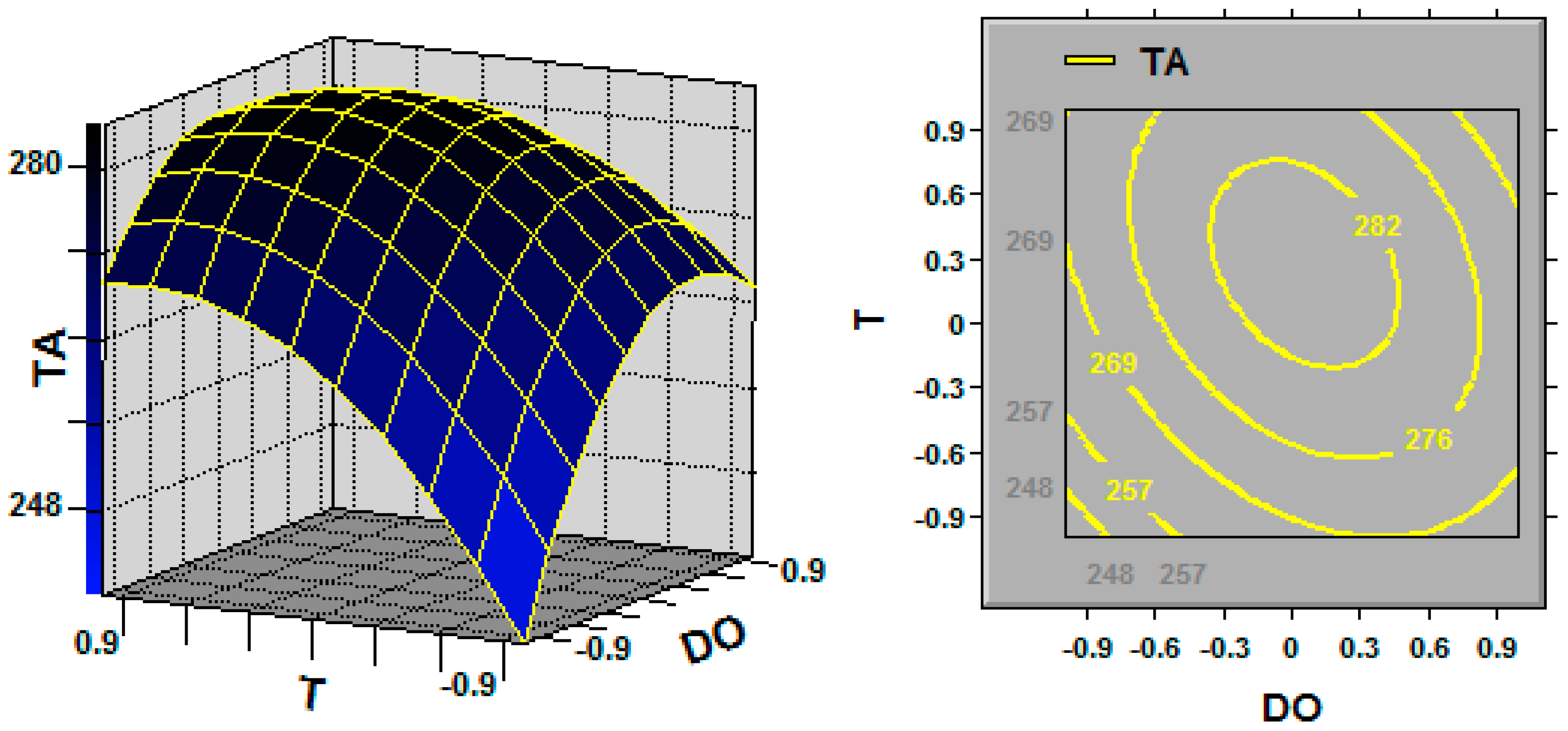
In statistics, response surface methodology (RSM) explores the relationships between several explanatory variables and one or more response variables, and the interactions between explanatory variables can be identified and quantified with this technique [
30,
31]. For a liquid rotation fermentation system for
Ganoderma, the key fermentation conditions (explanatory variables or factors) for TA production (response variable) are the initial pH, fermentation temperature and dissolved oxygen (DO) [
16,
17,
18,
32]. Previous studies have mainly focused on an individual fermentation variable of pH or dissolved oxygen [
16,
17,
18], and data about the relationships and interactions between several fermentation variables and TA production are scarce. Additionally, most studies have focused on
G. lucidum [
10,
16,
17,
18,
19,
20], and data about the submerged fermentation of TAs with
G. lingzhi are relatively limited. The aim of this work was to statistically optimize TA biosynthesis by the
Ganoderma species
G. lingzhi, which is distributed in East Asia, using RSM to develop a simple and feasible batch fermentation technology for large-scale TA production and to evaluate the in vitro and in vivo antitumor properties of the TA from submerged culture
G. lingzhi mycelium.
3. Discussion
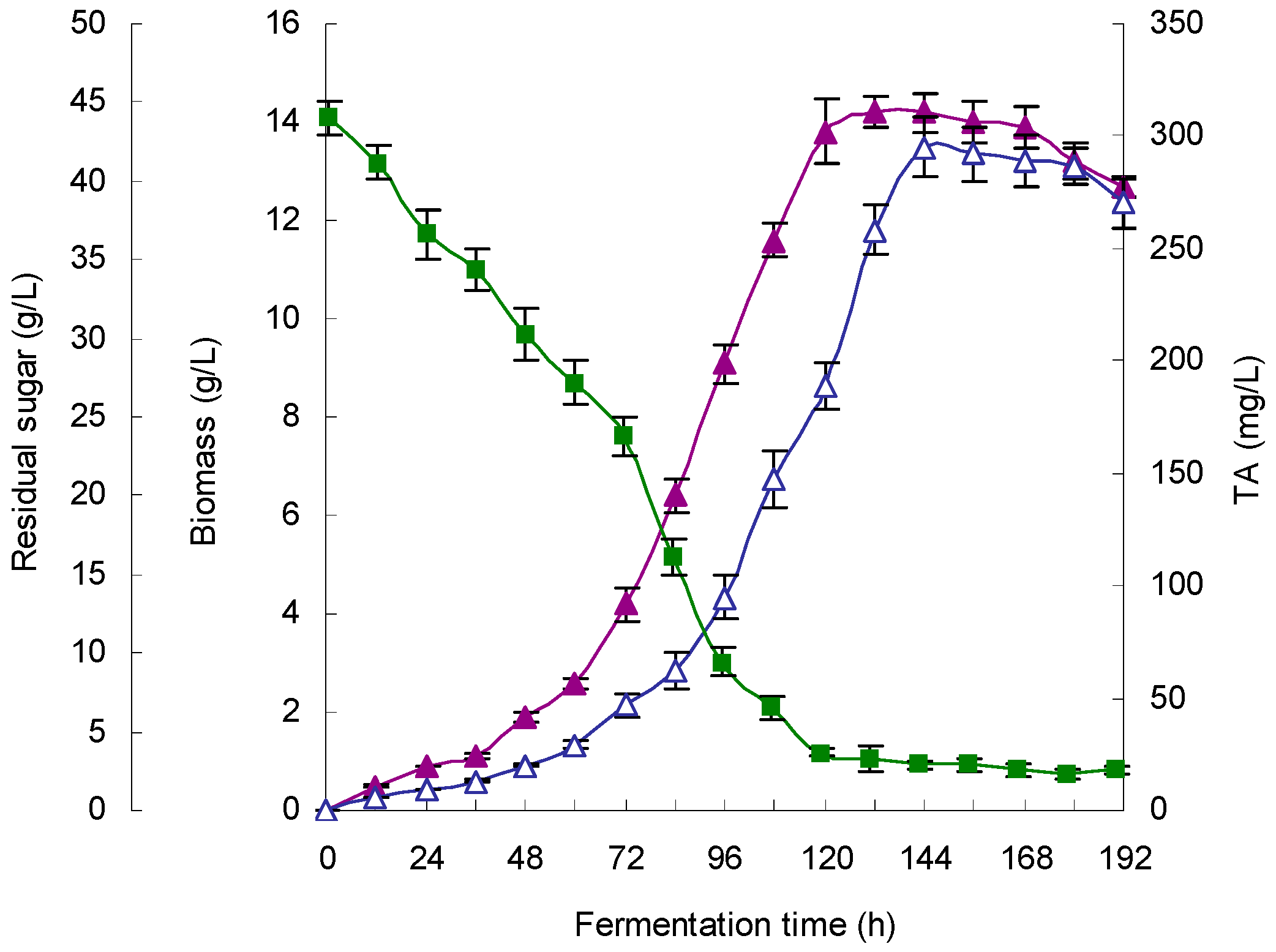
Dissolved oxygen (DO) has an important influence on the aerobic microbial fermentation process of. Oxygen supply affects cell growth, nutrient uptake and metabolite biosynthesis. Typically, sufficient oxygen supply results in an increase in the specific growth rate; however, the relationship between oxygen supply and metabolite biosynthesis is not clear [
33,
34,
35,
36]. In the cultivation of the filamentous fungus
Schizophyllum commune, a sufficient oxygen supply results in a decrease in the production of extracellular glucan [
33], whereas in the submerged fermentation of
Monascus ruber, improving the oxygen supply increases both the biomass and secondary metabolite (red pigment and citrinin) yields [
34]. Regarding
Ganoderma fermentation, one study reported that the production of ganoderic acids (a class of lanostane-type triterpenes) and polysaccharides by
G. lucidum are significantly enhanced in 2-L stirred-tank bioreactors when the DO is maintained between 20% and 35% air saturation [
35]. Another study revealed that the production of the extracellular polysaccharides of
G. lucidum under constant DO control at 10% is higher than that under 25% DO [
36]. In the present study, we first examined the effects of DO on TA production by
G. lingzhi; the TA yield was increased by maintaining the DO at 20% over the entire fermentation process, and the cell growth was also promoted by maintaining the DO at 20%.
Environmental factors, such as culture pH and temperature, can greatly affect metabolite production. Both of these factors can affect the uptake of various nutrients and the activity of important enzymes associated with metabolite biosynthesis [
37,
38,
39,
40], and the culture pH can also affect the cell membrane function [
37,
38,
39]. Regarding
Ganoderma fermentation, Fang and Zhong [
37] found that at an initial pH of 6.5, a maximum biomass of 17.3 g/L as well as a maximal production of ganoderic acid (a class of lanostane-type triterpenes) of 207.9 mg/L were obtained in a shake-flask culture of
G. lucidum at 30 °C, and the suitable initial pH and temperature for
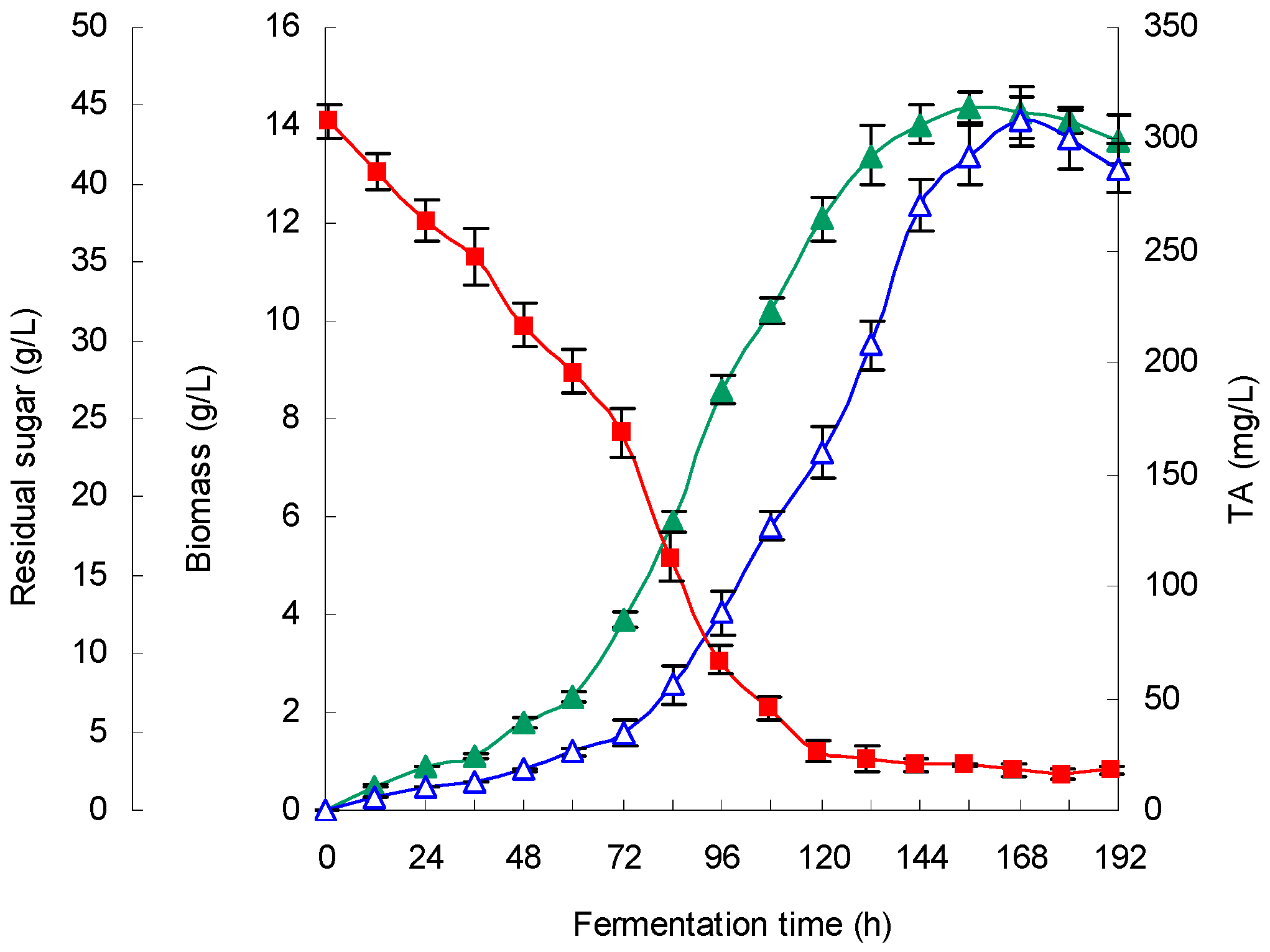
G. lingzhi fermentation were found to be 5.9 and 28.6 °C, respectively, in this work, and a TA production of 308.1 mg/L was obtained in a 5-L stirred bioreactor.
The scaling up of key fermentation factors is essential for potential commercial applications. There are some reports about large-scale submerged mushroom fermentation [
41,
42]. The scaling up of the filamentous fungus
Glarea lozoyensis from a pilot scale (0.07 m
3, 0.8 m
3, and 19 m
3) to a production scale (57 m
3) to produce the pharmaceutically important compound pneumocandin has been successfully completed by maintaining a constant DO [
41]. In the submerged fermentation of
Streptomyces gilvosporeus LK-196, the dissolved oxygen level and culture pH are also considered to be two key factors in the scaling up from 30- and 1000- to 18,000-L, and a high level of natamycin production was reproduced with the control of the dissolved oxygen level over 30% and a constant pH of 7.0 [
42]. Tang et al. developed a good fermentation strategy involving a pH shift and a DOT shift integrated into a fed-batch fermentation process for total ganoderic acid production by
G. lucidum [
17], and this process was successfully step-wise scaled-up from a 7.5- to a 200-L stirred-tank bioreactor [
43]. Nevertheless, previous studies on
Ganoderma have mainly focused on an individual fermentation factor, i.e., pH or dissolved oxygen [
16,
17,
18,
35,
36,
37], and the interactions between the fermentation variables were not been investigated. In the present work, the relationships between several fermentation variables (i.e., pH, dissolved oxygen and temperature) and TA production and the interactions between the fermentation variables were analyzed by RSM. The TA was examined following a simple batch fermentation process, and it was found to efficiently accumulate in the
G. lingzhi mycelium following statistical optimization of the key fermentation parameters. Moreover, the optimized process conditions were successfully reproduced in 5-L stirred-tank bioreactors and were also scaled up to a 200-L fermenter while the DO was controlled at 20.0%, the pH at 5.9 and the temperature at 28.6 °C. Furthermore, the
G. lingzhi fermentation time (6 days) in the 200-L scaled bioreactor was much shorter than that (18 days) of the fed-batch fermentation process for
G. lucidum that has been reported in the literature [
17]. Thus, the batch fermentation process was simpler, which may make the process easier to apply in large-scale production.
Ganoderma triterpenes have been reported to be cytotoxic to several cancer cell lines in vitro and to inhibit tumor invasion in vitro and in vivo [
4,
5,
6,
27,
28,
29,
44,
45,
46,
47]. Currently, most of the triterpenes and TAs with reported antitumor activities are isolated from the fruiting bodies or spores of
Ganoderma (mainly
G. lucidum) [
4,
6,
27,
28,
29,
47]. Regardless of whether the triterpenes or TAs are prepared from submerged culture
Ganoderma (especially
G. lingzhi) mycelium, the potential antitumor activities of these compounds need to be further proven. Recently, a few works regarding triterpenes (or ganoderic acids) prepared from the mycelium of
G. lucidum have been completed. One study demonstrated a triterpene-enriched fraction, WEES-G6, prepared from the mycelium of
G. lucidum inhibits the growth of human hepatoma Huh-7 cells by suppressing protein kinase C, activating mitogen-activated protein kinases and arresting the cell cycle in the G2 phase [
48]. Another work indicated that ganoderic acid T isolated from
G. lucidum mycelium inhibits the proliferation of human colon carcinoma HCT-116 cells [
5]. Furthermore, three new ganoderic acid T derivatives display marked inhibitory effects on the growth of human cervical cancer HeLa cells, cause cell cycle arrest at the G1 phase and induce apoptosis [
49]. Ganoderic acid Me isolated from
G. lucidum mycelium inhibits proliferation, angiogenesis, and invasion and induces apoptosis in MDA-MB-231 cells [
50], and it also induces the apoptosis of competent T cells and increases the proportion of Treg cells [
51]. In this work, TA from the mycelium of
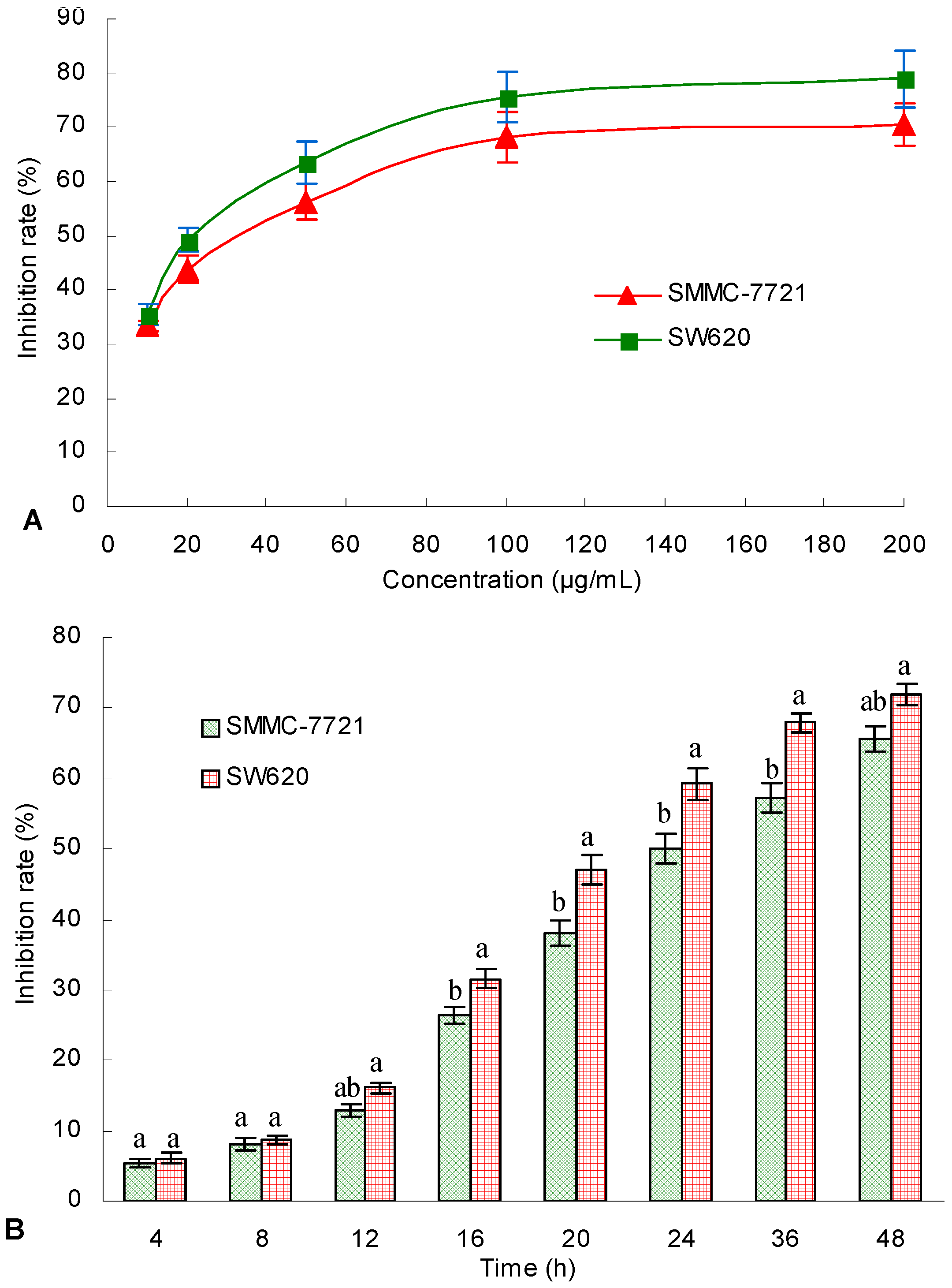
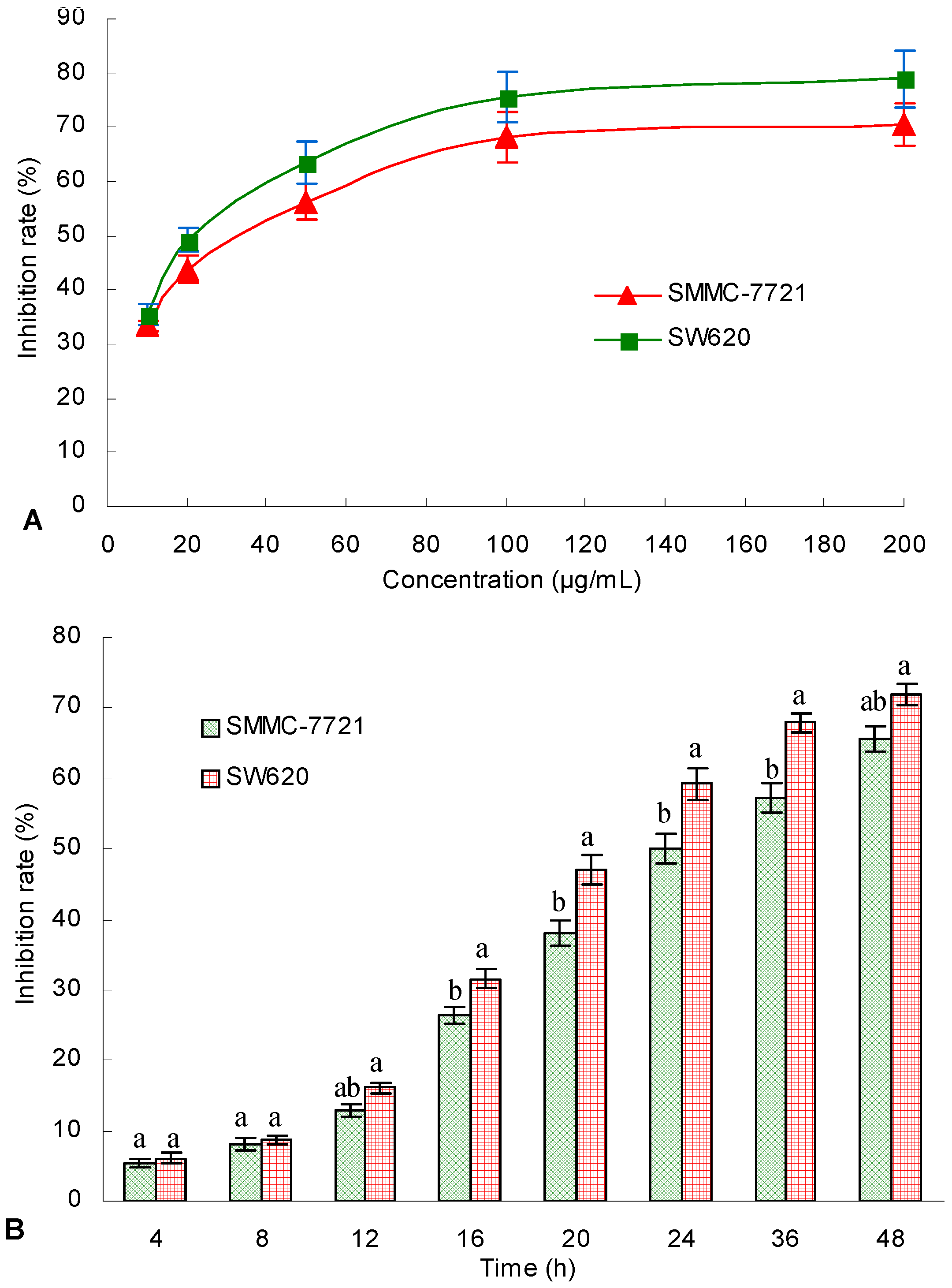
G. lingzhi was studied. For the first time, the TAE from submerged culture
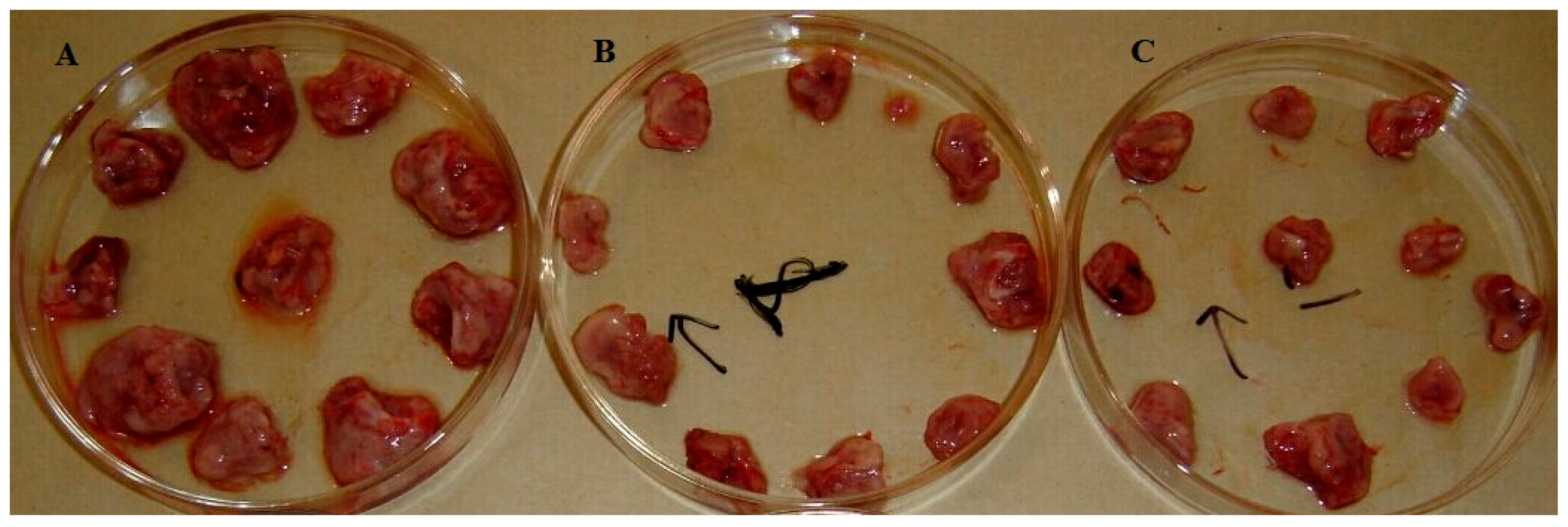
G. lingzhi mycelium was found to be cytotoxic to the human hepatoma SMMC-7721 and human colon carcinoma SW620 cell lines. Furthermore, the TAE (125 mg/kg and 250 mg/kg) from
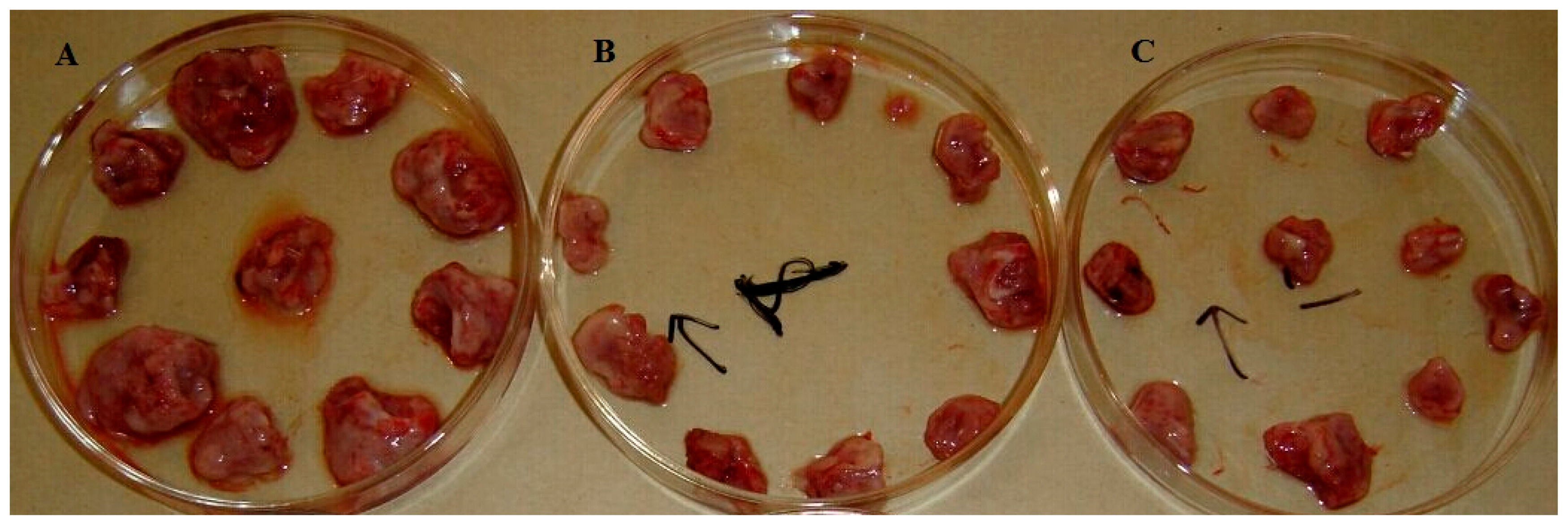
G. lingzhi mycelium obviously enhances thymus index and spleen index values in vivo, which suggests that the TA from
G. lingzhi mycelium might enhance body cell immune function at a certain dose. Hence, the antitumor effect of TA from
G. lingzhi mycelium is probably host-mediated and cytocidal. Additionally, previous studies have demonstrated that triterpene compounds, including ganoderic acids Me, T, DM and lucidenic acid F from
G. lucidum, have strong antitumor activities [
5,
28,
29,
44,
45]. Our results suggest that the TAE from the mycelium of
G. lingzhi also contains the typical TA compounds isolated from the fruiting bodies of
Ganoderma and that ganoderic acids T and Me dominantly exist in the extract. Therefore, the TAE from
G. lingzhi may have antitumor activities that are related to its high ganoderic acid T and Me contents. The identification of the other active principles and the elucidation of their mechanisms of action remain to be achieved.
In conclusion, enhanced TA production yield and productivity were attained with G. lingzhi via the optimization of the batch fermentation conditions. The TAE from G. lingzhi mycelium has strong active principle contents and possesses antitumor activity in vitro and in vivo.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}