
Phytochemical Compositions of Immature Wheat Bran, and Its Antioxidant Capacity, Cell Growth Inhibition, and Apoptosis Induction through Tumor Suppressor Gene

Abstract
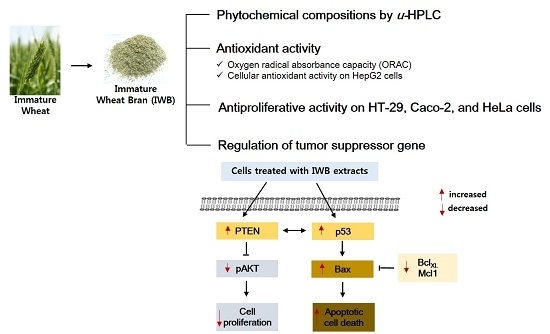
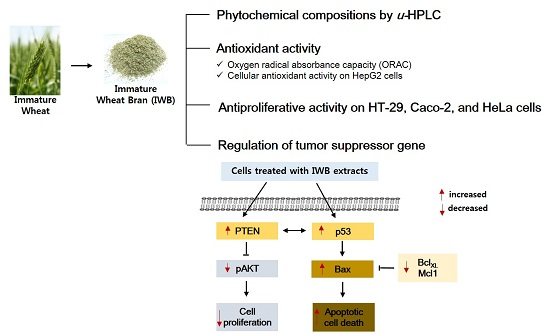
:
1. Introduction
2. Results and Discussion
2.1. Phenolic Acid Compositions of Wheat Bran Extracts
2.2. Antioxidant Capacity of Bran Extract Samples
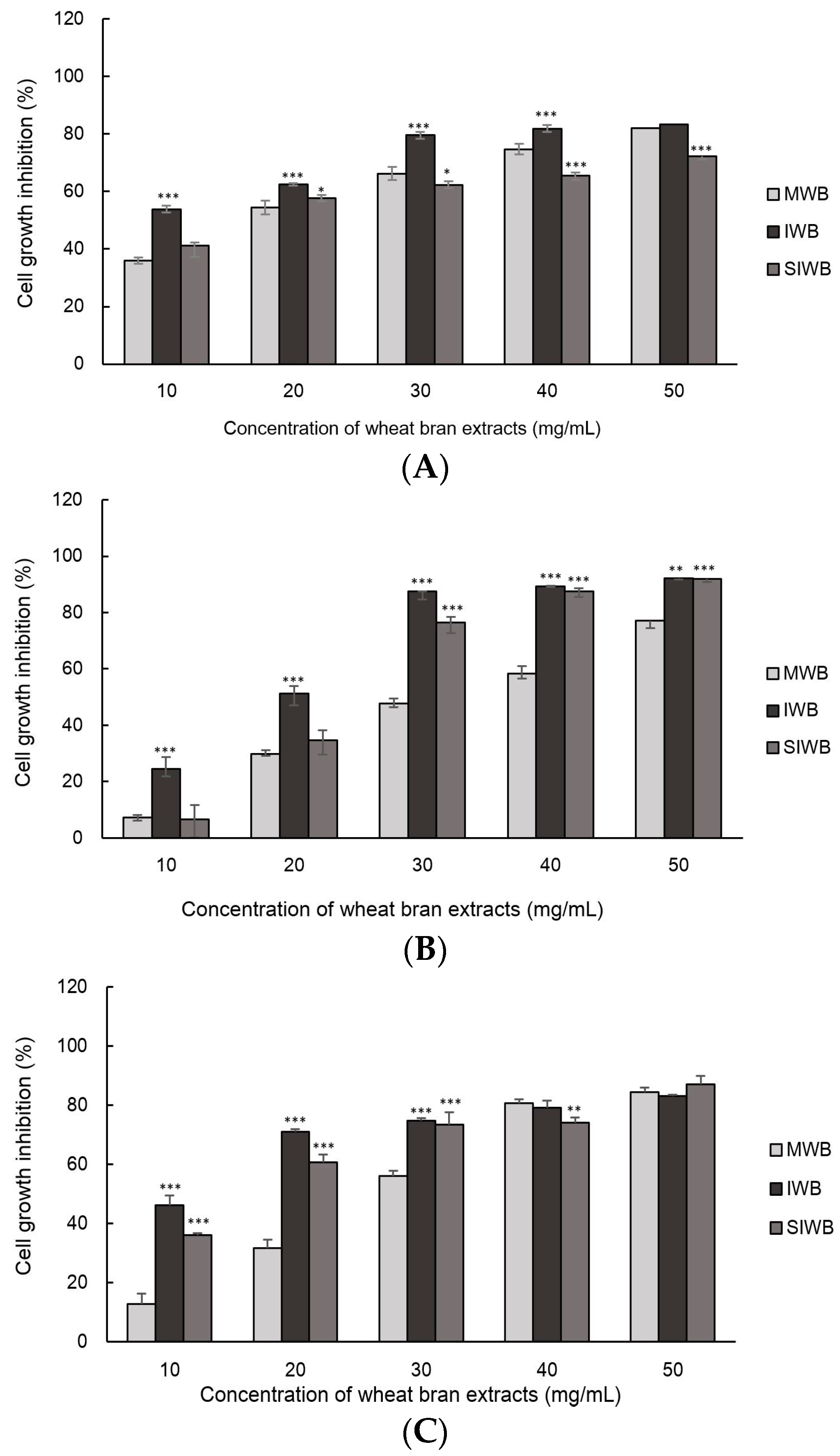
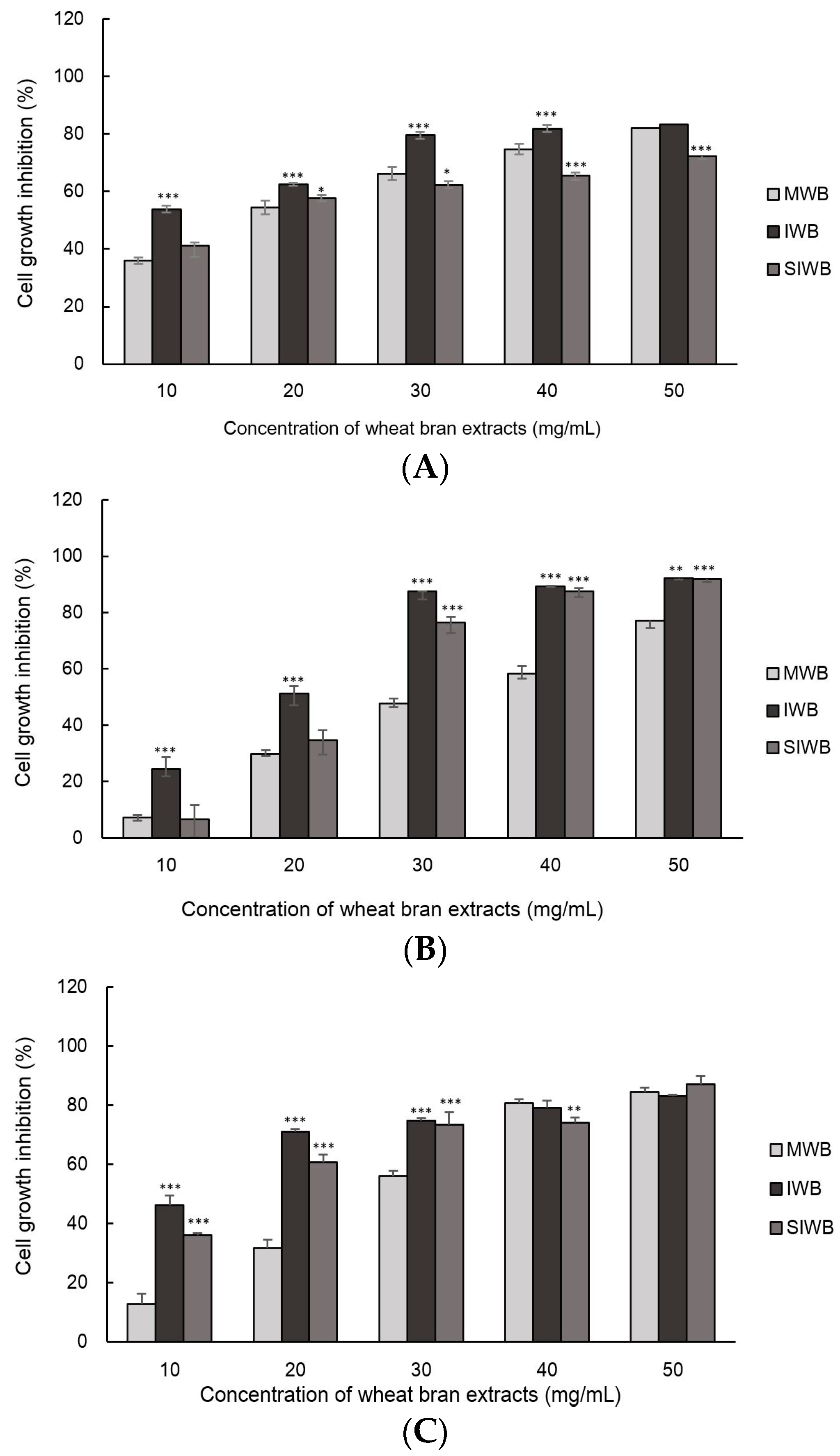
2.3. Effect of IWB Extract on Cell Growth of Human Carcinoma Cells
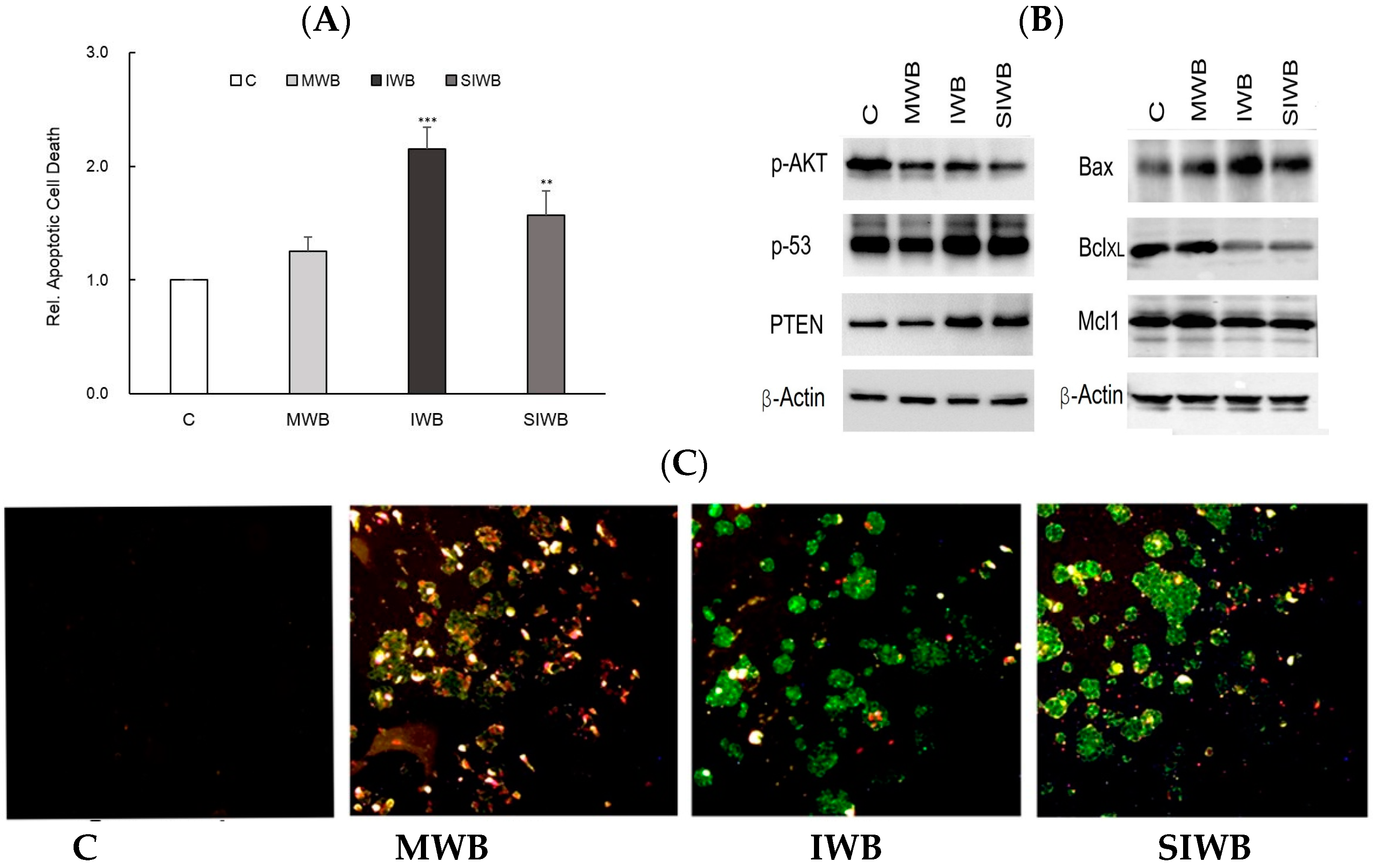
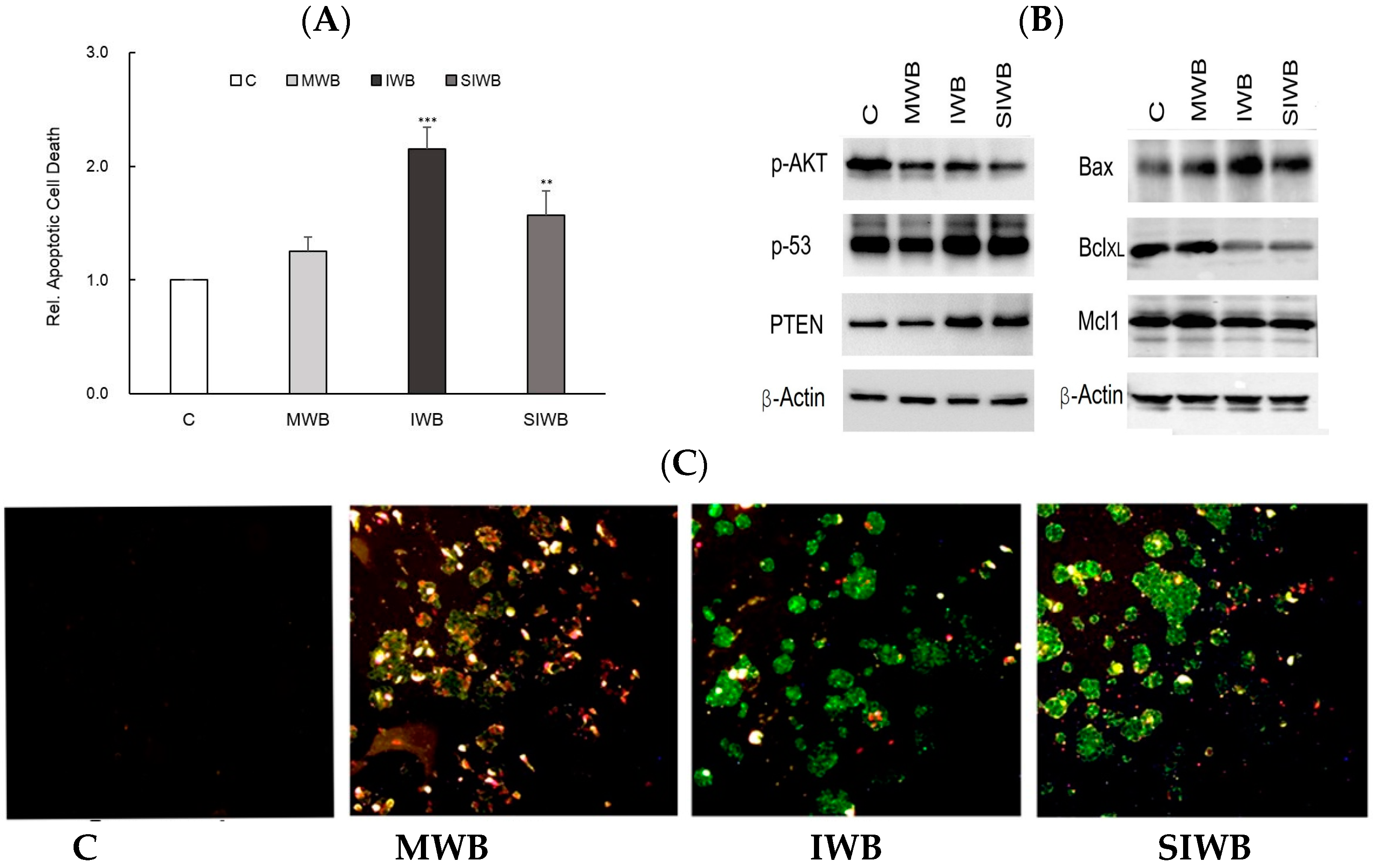
2.4. Induction of Apoptosis by Bran Extract
2.5. Correlations between Phenolic Compositions and Antioxidant, Antiprolferative, and Apoptosis Cell Death Properties in Wheat Bran Samples
3. Materials and Methods
3.1. Wheat Bran Samples
3.2. Chemicals and Reagents
3.3. Preparation of Free and Bound Extracts from Wheat Bran
3.4. Determination of Phenolic Compounds by Ultra-High Performance Liquid Chromatography
3.5. Determination of Antioxidant Capacity
3.6. Preparation of Bran Nextract for Treatment in Cell Cultures
3.7. Determination of Cellular Antioxidant Activity
3.8. Determination of Cell Growth and Apoptotic Cell Death
3.9. Western Immunoblots
3.10. Apopttic Cell Morphology by Green Fluorescent Protein
3.11. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO Cereal Supply and Demand Brief, Food and Agriculturre Organization of the United Nations. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 7 June 2016).
- Verma, B.; Hucl, P.; Chibbar, R.N. Phenolic content and antioxidant properties of bran in 51 wheat cultivars. Cereal Chem. 2008, 85, 544–549. [Google Scholar] [CrossRef]
- Anderson, J.W.; Hanna, T.J.; Peng, X.J.; Kryscio, R.J. Whole grain foods and heart disease risk. J. Am. Coll. Nutr. 2000, 19 (Suppl. 3), S291–S299. [Google Scholar] [CrossRef]
- Chatenoud, L.; Tavani, A.; La Vecchia, C.; Jacobs, D.R.; Negri, E.; Levi, F.; Franceschi, S. Whole grain food intake and cancer risk. Int. J. Cancer 1998, 77, 24–28. [Google Scholar] [CrossRef]
- Liu, S.M.; Manson, J.E.; Stampfer, M.J.; Hu, F.B.; Giovannucci, E.; Colditz, G.A.; Hennekens, C.H.; Willett, W.C. A prospective study of whole-grain intake and risk of type 2 diabetes mellitus in us women. Am. J. Public Health 2000, 90, 1409–1415. [Google Scholar] [PubMed]
- Lu, Y.J.; Lv, J.L.; Yu, L.; Fletcher, A.; Costa, J.; Yu, L.L.; Luthria, D. Phytochemical composition and antiproliferative activities of bran fraction of ten maryland-grown soft winter wheat cultivars: Comparison of different radical scavenging assays. J. Food Compos. Anal. 2014, 36, 51–58. [Google Scholar] [CrossRef]
- Laddomada, B.; Caretto, S.; Mita, G. Wheat bran phenolic acids: Bioavailability and stability in whole wheat-based foods. Molecules 2015, 20, 15666–15685. [Google Scholar] [CrossRef] [PubMed]
- Verma, B.; Hucl, P.; Chibbar, R.N. Phenolic acid composition and antioxidant capacity of acid and alkali hydrolysed wheat bran fractions. Food Chem. 2009, 116, 947–954. [Google Scholar] [CrossRef]
- Aktas, K.; Demirci, T.; Akin, N. Chemical composition and microbiological properties of tarhana enriched with immature wheat grain. J. Food Process. Preserv. 2015, 39, 3014–3021. [Google Scholar] [CrossRef]
- Iametti, S.; Bonomi, F.; Pagani, M.A.; Zardi, M.; Cecchini, C.; D’Egidio, M.G. Properties of the protein and carbohydrate fractions in immature wheat kernels. J. Agric. Food Chem. 2006, 54, 10239–10244. [Google Scholar] [CrossRef] [PubMed]
- Song, J.F.; Liu, C.Q.; Li, D.J.; Gu, Z.X. Postharvest changes in physicochemical characteristics and free amino acids content of immature vegetable soya bean (Glycine max L.) grains. Int. J. Food Sci. Technol. 2016, 51, 461–469. [Google Scholar] [CrossRef]
- Xu, J.G.; Hu, Q.P.; Wang, X.D.; Luo, J.Y.; Liu, Y.; Tian, C.R. Changes in the main nutrients, phytochemicals, and antioxidant activity in yellow corn grain during maturation. J. Agric. Food Chem. 2010, 58, 5751–5756. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Shin, J.A.; Gan, L.J.; Zhu, X.M.; Hong, S.T.; Sung, C.K.; Cho, J.W.; Ku, J.H.; Lee, K.T. Comparison of nutritional compounds in premature green and mature yellow whole wheat in Korea. Cereal Chem. 2012, 89, 284–289. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, S.S. Antioxidant and antiproliferative activities in immature and mature wheat kernels. Food Chem. 2016, 196, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Okarter, N.; Liu, R.H. Health benefits of whole grain phytochemicals. Crit. Rev. Food Sci. Nutr. 2010, 50, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic acid profiles and antioxidant activities of wheat bran extracts and the effect of hydrolysis conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, T.; Fu, X.; Abbasi, A.M.; Zheng, B.S.; Liu, R.H. Phenolics content, antioxidant and antiproliferative activities of dehulled highland barley (Hordeum vulgare L.). J. Funct. Foods 2015, 19, 439–450. [Google Scholar] [CrossRef]
- Hung, P.V. Phenolic compounds of cereals and their antioxidant capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.; Hao, Z.G.; Zhou, K.Q.; Luther, M.; Costa, J.; Yu, L.L. Carotenoid, tocopherol, phenolic acid, and antioxidant properties of maryland-grown soft wheat. J. Agric. Food Chem. 2005, 53, 6649–6657. [Google Scholar] [CrossRef] [PubMed]
- Okarter, N.; Liu, C.S.; Sorrells, M.E.; Liu, R.H. Phytochemical content and antioxidant activity of six diverse varieties of whole wheat. Food Chem. 2010, 119, 249–257. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F. Antioxidant properties of commercial soft and hard winter wheats (Triticum aestivum L.) and their milling fractions. J. Sci. Food Agric. 2006, 86, 477–485. [Google Scholar] [CrossRef]
- Onipe, O.O.; Jideani, A.I.O.; Beswa, D. Composition and functionality of wheat bran and its application in some cereal food products. Int. J. Food Sci. Technol. 2015, 50, 2509–2518. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Structure-activity relationships of flavonoids in the cellular antioxidant activity assay. J. Agric. Food Chem. 2008, 56, 8404–8411. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Chen, J.Y.; Xie, H.H.; Ju, X.R.; Liu, R.H. Phytochemical profiles and antioxidant activity of adlay varieties. J. Agric. Food Chem. 2013, 61, 5103–5113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.Z.; Liu, R.H. Phenolic and carotenoid profiles and antiproliferative activity of foxtail millet. Food Chem. 2015, 174, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Zand, H.; Jeddi-Tehrani, M.; Koohdani, F.; Shidfar, F.; Keshavarz, S.A. PTEN over-expression by resveratrol in acute lymphoblastic leukemia cells along with suppression of AKT/PKB and ERK1/2 in genotoxic stress. J. Nat. Med. 2015, 69, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Busca, A.; Saxena, M.; Kumar, A. Critical role for antiapoptotic bcl-xl and mcl-1 in human macrophage survival and cellular iap1/2 (ciap1/2) in resistance to hiv-vpr-induced apoptosis. J. Biol. Chem. 2012, 287, 15118–15133. [Google Scholar] [CrossRef] [PubMed]
- Woo, H.J.; Park, K.Y.; Rhu, C.H.; Lee, W.H.; Choi, B.T.; Kim, G.Y.; Park, Y.M.; Choi, Y.H. β-Lapachone, a quinone isolated from tabebuia avellanedae, induces apoptosis in HepG2 hepatoma cell line through induction of bax and activation of caspase. J. Med. Food 2006, 9, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Sevgi, K.; Tepe, B.; Sarikurkcu, C. Antioxidant and DNA damage protection potentials of selected phenolic acids. Food Chem. Toxicol. 2015, 77, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Whent, M.; Huang, H.Q.; Xie, Z.H.; Lutterodt, H.; Yu, L.; Fuerst, E.P.; Morris, C.F.; Yu, L.L.; Luthria, D. Phytochemical composition, anti-inflammatory, and antiproliferative activity of whole wheat flour. J. Agric. Food Chem. 2012, 60, 2129–2135. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, L.R.; Zhu, S.T.; Harris, P.J. Antioxidant and antigenotoxic effects of plant cell wall hydroxycinnamic acids in cultured HT-29 cells. Mol. Nutr. Food Res. 2005, 49, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the immature wheat bran is available from the authors.

{kind=link}
{kind=link}
{kind=link}
| Phenolic Acid Composition 1 | MWB | IWB | SIWB | |
|---|---|---|---|---|
| Ferulic acid (µg/g) | Free *** | 26.96 ± 2.78 a | 20.17 ± 0.38 b | 13.37 ± 0.47 c |
| Bound *** | 1767 ± 3.00 c | 3071 ± 32.8 a | 2532 ± 45.7 b | |
| Total 2,*** | 1794 ± 2.80 c | 3091 ± 32.9 a | 2545 ± 45.4 b | |
| Sinapic acid (µg/g) | Free *** | 2.46 ± 0.18 b | 3.95 ± 0.44 a | 3.51 ± 0.22 a |
| Bound | 118 ± 1.68 a | 103 ± 21.12 a | 120 ± 3.00 a | |
| Total 2 | 121 ± 1.79 a | 107 ± 21.34 a | 124 ± 2.90 a | |
| Vanillic acid (µg/g) | Free ** | 11.98 ± 0.21 a | 12.20 ± 1.95 a | 8.07 ± 0.52 b |
| Bound *** | 62.52 ± 0.21 a | 51.52 ± 1.31 b | 47.07 ± 1.61 c | |
| Total 2,*** | 74.49 ± 0.24 a | 63.73 ± 1.70 b | 55.14 ± 1.41 c | |
| p-Coumaric acid (µg/g) | Free *** | 3.22 ± 0.07 c | 5.61 ± 0.32 a | 4.68 ± 0.51 b |
| Bound *** | 51.97 ± 1.60 c | 68.98 ± 1.51 a | 58.93 ± 1.75 b | |
| Total 2,*** | 55.19 ± 1.65 c | 74.58 ± 1.38 a | 63.61 ± 1.53 b | |
| Syringic acid (µg/g) | Free | 3.38 ± 0.03 a | 3.63 ± 0.03 a | 2.39 ± 0.31 a |
| Bound ** | 26.11 ± 1.56 a | 18.01 ± 1.51 b | 19.08 ± 1.68 b | |
| Total 2,** | 29.50 ± 1.56 a | 21.64 ± 1.38 b | 21.47 ± 1.54 b | |
| Syringaldehyde (µg/g) | Free *** | 3.07 ± 0.02 c | 12.32 ± 0.33 a | 10.28 ± 0.36 b |
| Bound *** | 13.67 ± 0.14 c | 18.70 ± 0.64 a | 16.33 ± 0.48 b | |
| Total 2,*** | 16.74 ± 0.16 c | 31.02 ± 0.84 a | 26.60 ± 0.40 b | |
| Caffeic acid (µg/g) | Free | 2.38 ± 0.03 a | 2.14 ± 0.18 a | 1.99 ± 0.10 a |
| Bound | 7.87 ± 1.14 a | 9.52 ± 1.12 a | 10.14 ± 0.44 a | |
| Total 2 | 10.25 ± 1.17 a | 11.65 ± 1.01 a | 12.13 ± 0.51 a | |
| Antioxidant Capacity | MWB | IWB | SIWB | |
|---|---|---|---|---|
| ORAC 1 (µM TE/g) | Free *** | 44 ± 3.9 ab | 54 ± 8.3 a | 38 ± 3.6 b |
| Bound *** | 237 ± 4.7 b | 273 ± 13 a | 252 ± 17 ab | |
| Total 2,*** | 281 ± 4.7 b | 327 ± 11 a | 290 ± 20 b | |
| CAA 1,*** (µM QE/g) | 0.63 ± 0.07 c | 4.59 ± 0.17 a | 2.58 ± 0.47 b | |
| Bran Extracts | EC50 (mg/mL) | ||
|---|---|---|---|
| Caco-2 *** | HT-29 *** | HeLa *** | |
| MWB | 17.25 ± 0.84 a | 33.25 ± 1.07 a | 28.00 ± 2.24 a |
| IWB | 7.62 ± 0.91 c | 18.92 ± 2.09 c | 7.74 ± 1.31 c |
| SIWB | 15.25 ± 0.18 b | 23.43 ± 1.58 b | 15.25 ± 0.85 b |
| Phenolic Compositions | ORAC | CAA | Cell Growth Inhibition | Apoptotic Cell Death | |||
|---|---|---|---|---|---|---|---|
| Caco-2 | HT-29 | HeLa | |||||
| Ferulic acid | Free | −0.170 | –0.477 | −0.352 | −0.191 | −0.689 * | −0.278 |
| Bound | 0.815 ** | 0.988 *** | 0.939 *** | 0.863 *** | 0.975 *** | 0.882 *** | |
| Sinapic acid | Free | 0.645 | 0.914 *** | 0.829 ** | 0.786 * | 0.927 *** | 0.828 *** |
| Bound | −0.685 * | −0.530 | −0.615 | −0.468 | −0.356 | −0.305 | |
| Vanillic acid | Free | 0.397 | 0.054 | 0.201 | 0.188 | −0.210 | 0.103 |
| Bound | −0.428 | −0.689 * | −0.581 | −0.402 | −0.833 *** | −0.471 | |
| p-Coumaric acid | Free | 0.823 ** | 0.961 *** | 0.933 *** | 0.748 * | 0.933 *** | 0.795 *** |
| Bound | 0.779 * | 0.950 *** | 0.954 *** | 0.918 *** | 0.912 *** | 0.966 *** | |
| Syringic acid | Free | 0.460 | 0.211 | 0.297 | 0.390 | −0.085 | 0.269 |
| Bound | −0.716 * | −0.871 ** | −0.768 * | −0.637 | −0.906 *** | −0.619 | |
| Syringaldehyde | Free | 0.713 * | 0.940 *** | 0.880 ** | 0.749 * | 0.989 *** | 0.812 *** |
| Bound | 0.781 * | 0.961 *** | 0.960 *** | 0.856 ** | 0.956 *** | 0.890 *** | |
| Caffeic acid | Free | −0.301 | −0.488 | −0.481 | −0.235 | −0.675 * | −0.366 |
| Bound | 0.405 | 0.562 | 0.446 | 0.274 | 0.676 * | 0.261 | |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.J.; Yoon, W.-J.; Kim, S.S. Phytochemical Compositions of Immature Wheat Bran, and Its Antioxidant Capacity, Cell Growth Inhibition, and Apoptosis Induction through Tumor Suppressor Gene. Molecules 2016, 21, 1292. https://doi.org/10.3390/molecules21101292
Kim MJ, Yoon W-J, Kim SS. Phytochemical Compositions of Immature Wheat Bran, and Its Antioxidant Capacity, Cell Growth Inhibition, and Apoptosis Induction through Tumor Suppressor Gene. Molecules. 2016; 21(10):1292. https://doi.org/10.3390/molecules21101292
Chicago/Turabian StyleKim, Mi Jeong, Won-Jin Yoon, and Sang Sook Kim. 2016. "Phytochemical Compositions of Immature Wheat Bran, and Its Antioxidant Capacity, Cell Growth Inhibition, and Apoptosis Induction through Tumor Suppressor Gene" Molecules 21, no. 10: 1292. https://doi.org/10.3390/molecules21101292