
Multigram Synthesis and in Vivo Efficacy Studies of a Novel Multitarget Anti-Alzheimer’s Compound

 ,
,  ,
, 

Abstract
:
1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | hAChE IC50 nM | hBChE IC50 nM | BACE-1 IC50 µM | Aβ Aggreg. % Inhibition at 10 µM a | PAMPA-BBB Pe (10‒6 cm·s‒1) b (Prediction) | Ex Vivo Studies c % Inhibition Brain AChE |
|---|---|---|---|---|---|---|
| AVCRI104P4 | 2.61 | 349 | 11.0 | 29 | 11.4 (CNS+) | 59% at 5 min d |
| Donepezil | 21.4 | 7273 | 11.3 | <5 | 25.2 (CNS+) | 73% at 5 min e |
| (‒)-Huprine Y | 0.43 | 247 | Nd f | 10.2 | 18.2 (CNS+) | 97% at 20 min |
2. Results and Discussion
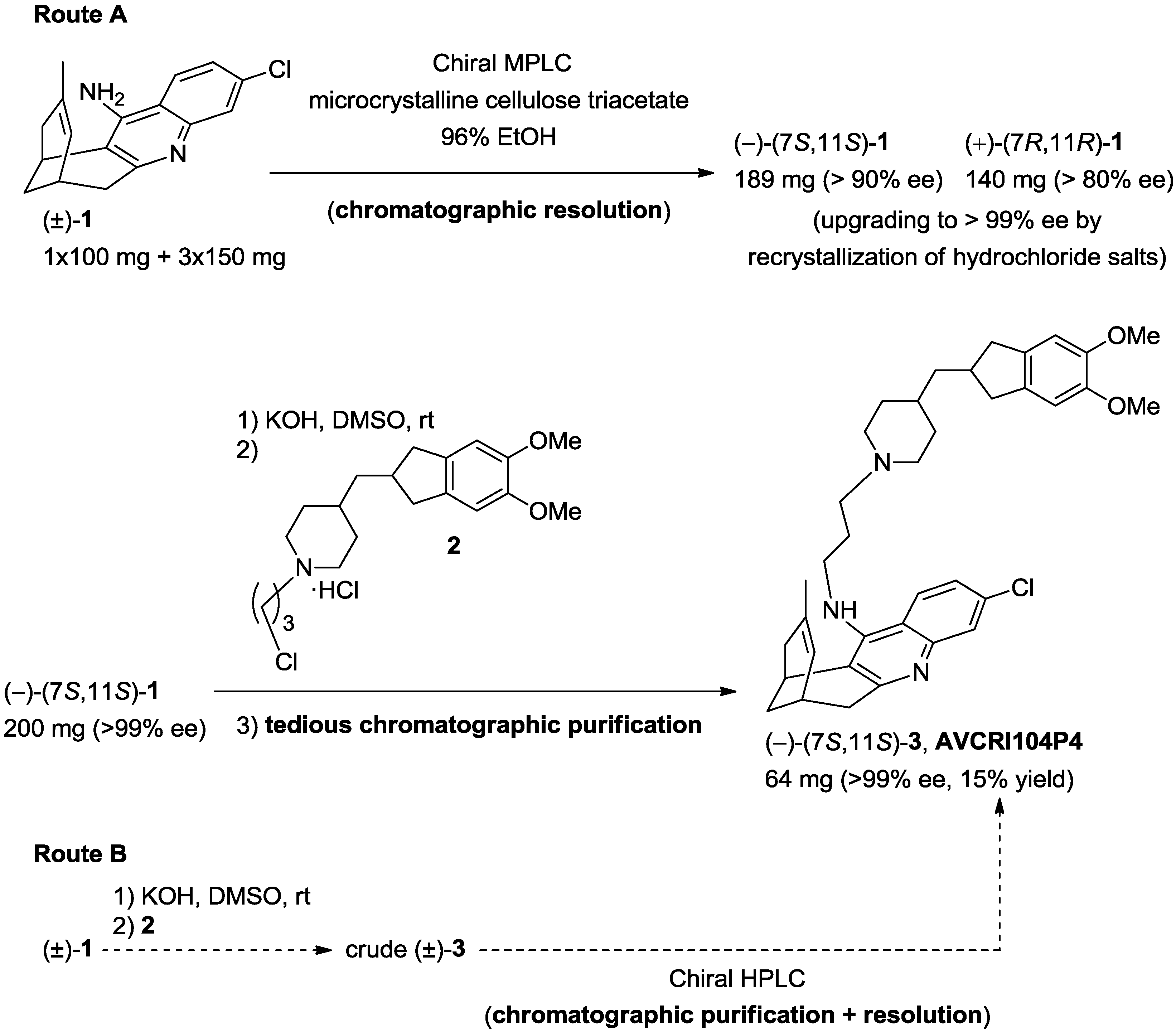
2.1. Synthesis of AVCRI104P4
2.2. In Vivo Studies in C. elegans

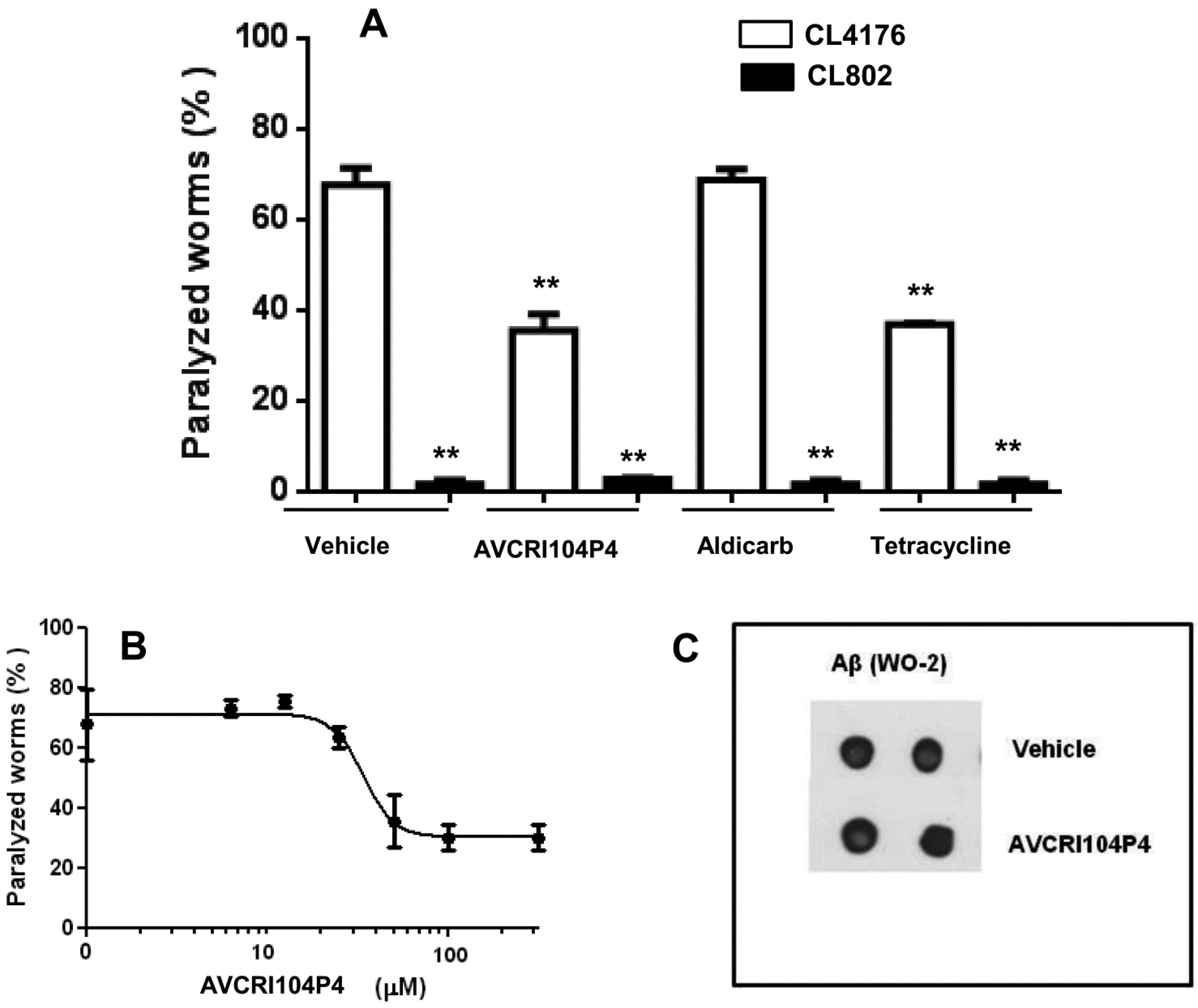
2.3. In Vivo Studies in Transgenic APPSL Mice
2.3.1. Behavioral Studies
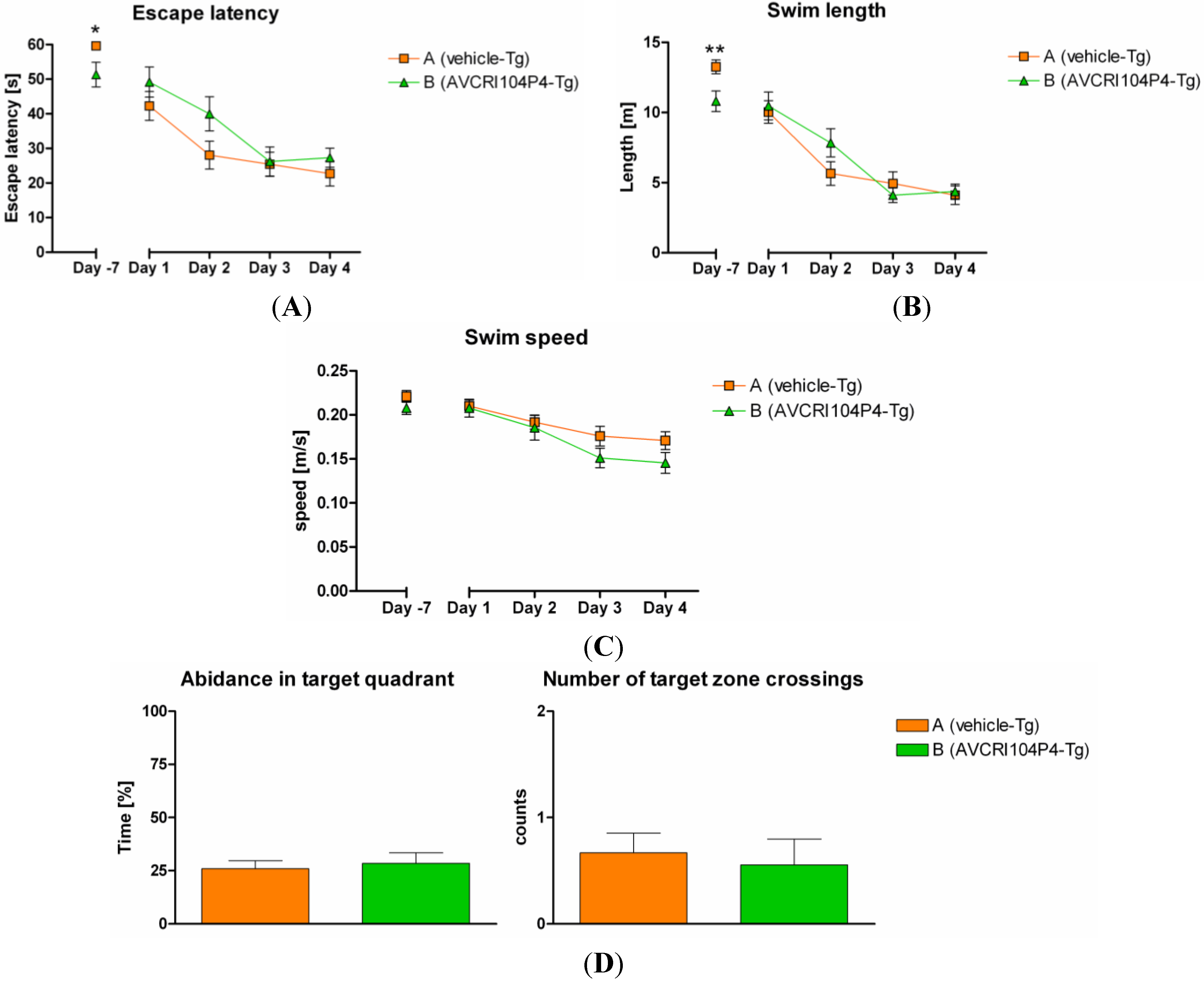
2.3.2. Biochemistry and Histology
2.4. Hepatotoxicity Studies in HepG2 Cells

3. Experimental Section
3.1. Synthesis of AVCRI104P4
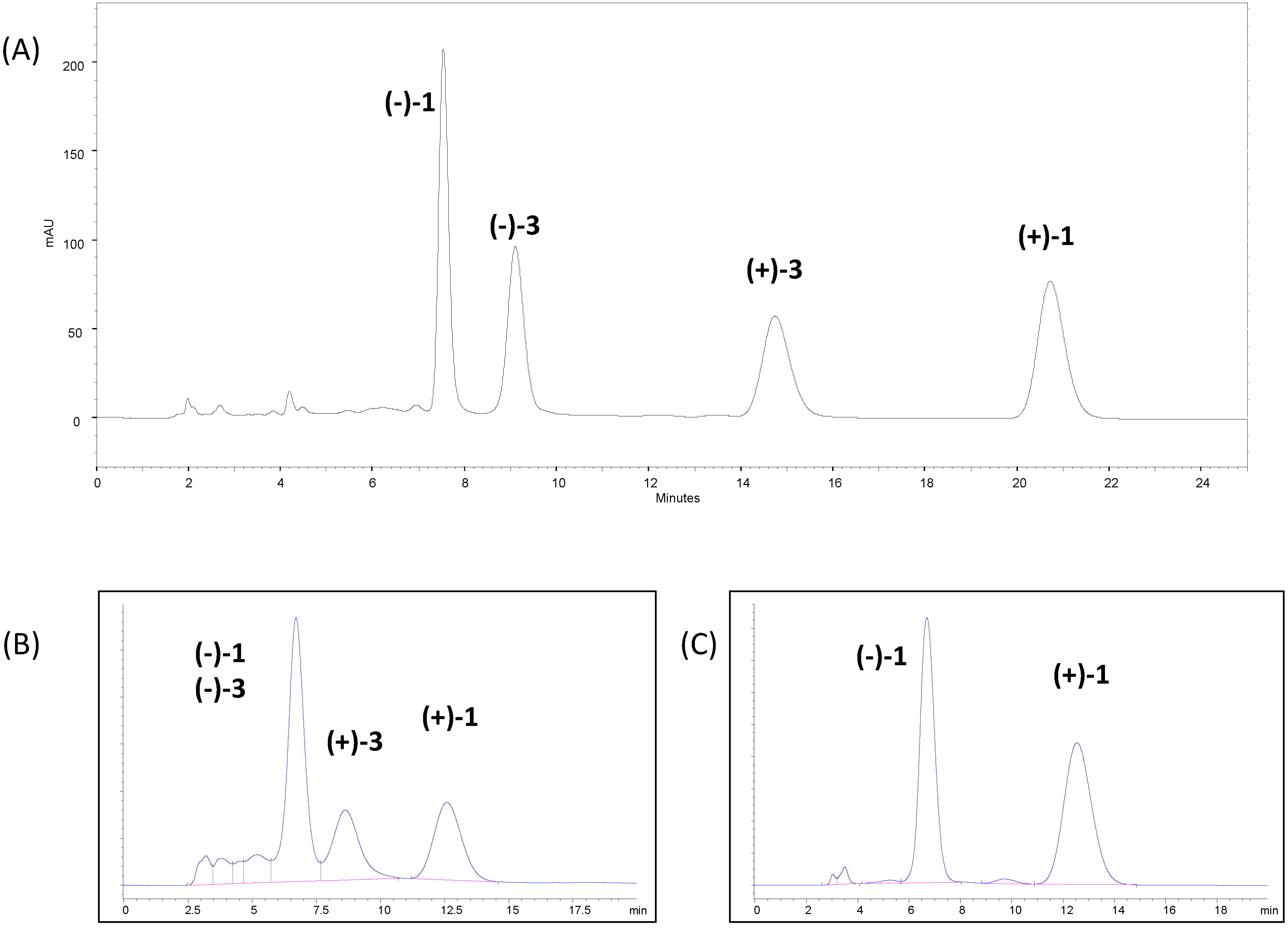
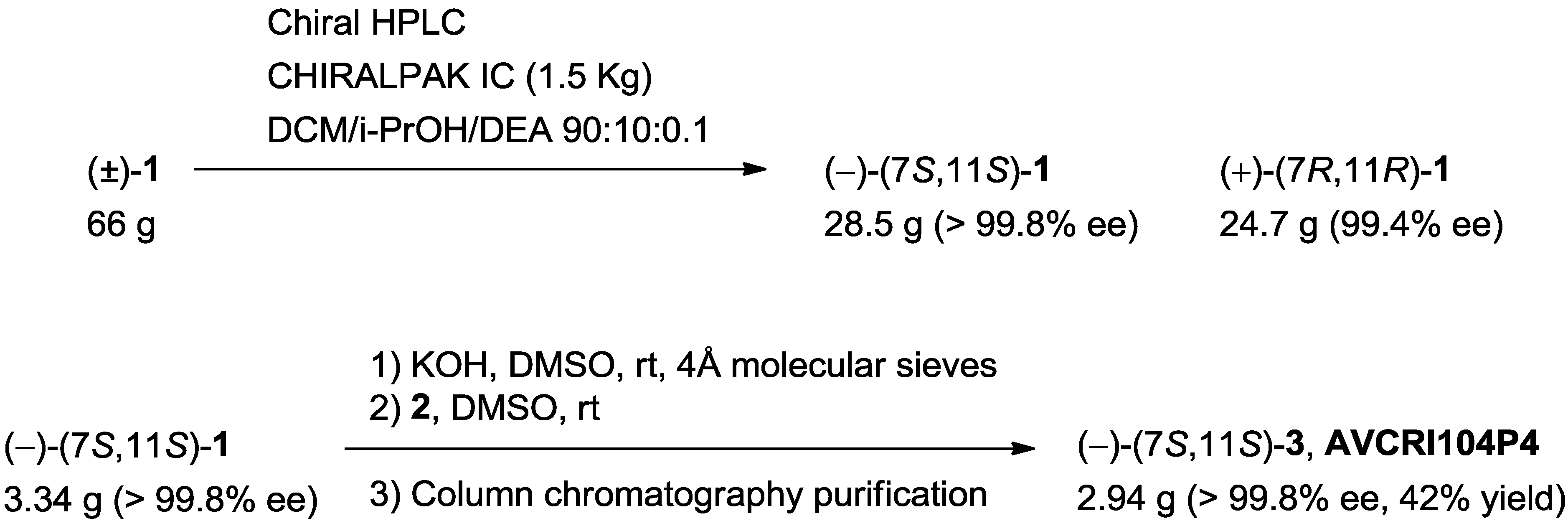
3.1.1. Chromatographic Screening
3.1.2. Chromatographic Resolution of (±)-Huprine Y at Preparative Scale
3.1.3. (–)-(7S,11S)-3-Chloro-12-[(3-{4-[(5,6-dimethoxyindan-2-yl)methyl]piperidin-1-yl}propyl) amino]-6,7,10,11-tetrahydro-9-methyl-7,11-metanocycloocta[b]quinoline, AVCRI104P4 [(–)-(7S,11S)-3]
3.2. In Vivo Studies in Caenorhabditis elegans
3.2.1. C. elegans Strains
3.2.2. Paralysis Assay
3.2.3. Aβ Expression
3.2.4. Staining of β-Amyloid
3.2.5. Statistical Analysis
3.3. In Vivo Studies in Transgenic APPSL Mice
3.3.1. Animals and Treatment
3.3.2. Behavioural Studies
3.3.3. Tissue Sampling
3.3.4. Biochemistry
Brain Protein Extraction
Determination of Aβ Levels
3.3.5. Histology
Sectioning, Immunohistochemistry and Image Analysis
Determination of Plaque Load
Imaging
Evaluation of Amyloid Depositions and Plaque Load
Statistical Analysis
3.4. Hepatotoxicity Studies in HepG2 Cells
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Prince, M.; Albanese, E.; Guerchet, M.; Prina, M. World Alzheimer Report 2014: Dementia and Risk Reduction; Alzheimer’s Disease International: London, UK, 2014; Available online: http://www.alz.co.uk (accessed on 20 February 2015).
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Citron, M. Alzheimer’s disease: Strategies for disease modification. Nat. Rev. Drug Discov. 2010, 9, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Pimplikar, S.W. Reassessing the amyloid cascade hypothesis of Alzheimer’s disease. Int. J. Biochem. Cell Biol. 2009, 41, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, A.; Bolognesi, M.L.; Minarini, A.; Rosini, M.; Tumiatti, V.; Recanatini, M.; Melchiorre, C. Multi-target-directed ligands to combat neurodegenerative diseases. J. Med. Chem. 2008, 51, 347–372. [Google Scholar] [CrossRef] [PubMed]
- Guzior, N.; Więckowska, A.; Panek, D.; Malawska, B. Recent development of multifunctional agents as potential drug candidates for the treatment of Alzheimer’s disease. Curr. Med. Chem. 2015, 22, 373–404. [Google Scholar] [CrossRef] [PubMed]
- Geldenhuys, W.J.; van der Schyf, C.J. Rationally designed multi-targeted agents against neurodegenerative diseases. Curr. Med. Chem. 2013, 20, 1662–1672. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Decker, M. Multi-target compounds acting in the central nervous system designed from natural products. Curr. Med. Chem. 2013, 20, 1673–1685. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Frustaci, A.; Del Bufalo, A.; Fini, M.; Cesario, A. Multitarget drugs of plants origin acting on Alzheimer’s disease. Curr. Med. Chem. 2013, 20, 1686–1693. [Google Scholar] [CrossRef] [PubMed]
- Capurro, V.; Busquet, P.; Lopes, J.P.; Bertorelli, R.; Tarozzo, G.; Bolognesi, M.L.; Piomelli, D.; Reggiani, A.; Cavalli, A. Pharmacological characterization of memoquin, a multitarget compound for the treatment of Alzheimer’s disease. PLoS One 2013, 8, e56870. [Google Scholar] [CrossRef] [PubMed]
- Antequera, D.; Bolos, M.; Spuch, C.; Pascual, C.; Ferrer, I.; Fernandez-Bachiller, M.I.; Rodríguez-Franco, M.I.; Carro, E. Effects of a tacrine-8-hydroxyquinoline hybrid (IQM-622) on Aβ accumulation and cell death: Involvement in hippocampal neuronal loss in Alzheimer’s disease. Neurobiol. Dis. 2012, 46, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Viayna, E.; Gómez, T.; Galdeano, C.; Ramírez, L.; Ratia, M.; Badia, A.; Clos, M.V.; Verdaguer, E.; Junyent, F.; Camins, A.; et al. Novel huprine derivatives with inhibitory activity toward β-amyloid aggregation and formation as disease-modifying anti-Alzheimer drug candidates. ChemMedChem 2010, 5, 1855–1870. [Google Scholar] [CrossRef]
- Sussman, J.L.; Harel, M.; Frolow, F.; Oefner, C.; Goldman, A.; Toker, L.; Silman, I. Atomic structure of acetylcholinesterase from Torpedo californica: A prototypic acetylcholine-binding protein. Science 1991, 253, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Alvarez, A.; Pérez, C.A.; Moreno, R.D.; Vicente, M.; Linker, C.; Casanueva, O.I.; Soto, C.; Garrido, J. Acetylcholinesterase accelerates assembly of amyloid-β-peptides into Alzheimer’s fibrils: Possible role of the peripheral site of the enzyme. Neuron 1996, 16, 881–891. [Google Scholar] [CrossRef] [PubMed]
- De Ferrari, G.V.; Canales, M.A.; Shin, I.; Weiner, L.M.; Silman, I.; Inestrosa, N.C. A structural motif of acetylcholinesterase that promotes amyloid beta-peptide fibril formation. Biochemistry 2001, 40, 10447–10457. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Martinez, A. Targeting beta-amyloid pathogenesis through acetylcholinesterase inhibitors. Curr. Pharm. Des. 2006, 12, 4377–4387. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Torrero, D. Acetylcholinesterase inhibitors as disease-modifying therapies for Alzheimer’s disease. Curr. Med. Chem. 2008, 15, 2433–2455. [Google Scholar] [CrossRef] [PubMed]
- Di, L.; Kerns, E.H.; Fan, K.; McConnell, O.J.; Carter, G.T. High throughput artificial membrane permeability assay for blood-brain barrier. Eur. J. Med. Chem. 2003, 38, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Morphy, R.; Rankovic, Z. Designed multiple ligands. An emerging drug discovery paradigm. J. Med. Chem. 2005, 48, 6523–6543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Nguyen, D.; Franco, P. Enantiomer resolution screening strategy using multiple immobilised polysaccharide-based chiral stationary phases. J. Chromatogr. A 2008, 1191, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Franco, P.; Zhang, T. Common approaches for efficient method development with immobilised polysaccharide-derived chiral stationary phases. J. Chromatogr. B 2008, 875, 48–56. [Google Scholar] [CrossRef]
- Zhang, T.; Schaeffer, M.; Franco, P. Optimization of the chiral separation of a Ca-sensitizing drug on an immobilized polysaccharide-based chiral stationary phase. Case study with a preparative perspective. J. Chromatogr. A 2005, 1083, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Bertz, S.H. Tetramethyl 3,7-dihydroxybicyclo[3.3.1]nona-2,6-diene-2,4,6,8-tetracarboxylate: A useful companion to Meerwein’s ester. Topological analysis of bicyclo[3.3.1]nonane synthesis. J. Org. Chem. 1985, 50, 3585–3592. [Google Scholar] [CrossRef]
- Camps, P.; El Achab, R.; Font-Bardia, M.; Görbig, D.; Morral, J.; Muñoz-Torrero, D.; Solans, X.; Simon, M. Easy synthesis of 7-alkylbicyclo[3.3.1]non-6-en-3-ones by silica gel-promoted fragmentation of 3-alkyl-2-oxaadamant-1-yl mesylates. Tetrahedron 1996, 52, 5867–5880. [Google Scholar] [CrossRef]
- Ronco, C.; Jean, L.; Renard, P.-Y. Improved synthetic pathway for the derivatization of huprine scaffold. Tetrahedron 2010, 66, 7399–7404. [Google Scholar] [CrossRef]
- Camps, P.; Contreras, J.; Font-Bardia, M.; Morral, J.; Muñoz-Torrero, D.; Solans, X. Enantioselective synthesis of tacrine-huperzine A hybrids. Preparative chiral MPLC separation of their racemic mixtures and absolute configuration assignments by X-ray diffraction analysis. Tetrahedron Asymmetry 1998, 9, 835–849. [Google Scholar] [CrossRef]
- Camps, P.; Formosa, X.; Galdeano, C.; Gómez, T.; Muñoz-Torrero, D.; Scarpellini, M.; Viayna, E.; Badia, A.; Clos, M.V.; Camins, A.; et al. Novel donepezil-based inhibitors of acetyl- and butyrylcholinesterase and acetylcholinesterase-induced β-amyloid aggregation. J. Med. Chem. 2008, 51, 3588–3598. [Google Scholar] [CrossRef]
- Volovik, Y.; Carvalhal Marques, F.; Cohen, E. The nematode Caenorhabditis elegans: A versatile model for the study of proteotoxicity and aging. Methods 2014, 68, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Hassan, W.M.; Dostal, V.; Huemann, B.N.; Yerg, J.E.; Link, C.D. Identifying Aβ-specific pathogenic mechanisms using a nematode model of Alzheimer’s disease. Neurobiol. Aging 2015, 36, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Le, W. Modeling neurodegenerative diseases in Caenorhabditis elegans. Exp. Neurol. 2013, 250, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Sashidhara, K.V.; Modukuri, R.K.; Jadiya, P.; Prasad Dodda, R.; Kumar, M.; Sridhar, B.; Kumar, V.; Haque, R.; Imran Siddiqi, M.; Nazir, A. Benzofuran-chalcone hybrids as potential multifunctional agents against Alzheimer’s disease: Synthesis and in vivo studies with transgenic Caenorhabditis elegans. ChemMedChem 2014, 9, 2671–2684. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, L.; Lin, Z.; Liang, F.; Pei, Z.; Xu, J.; Gu, Q. Dehydroabietylamine derivatives as multifunctional agents for the treatment of Alzheimer’s disease. Med. Chem. Commun. 2014, 5, 1736–1743. [Google Scholar] [CrossRef]
- Cao, Y.Y.; Wang, L.; Ge, H.; Lu, X.L.; Pei, Z.; Gu, Q.; Xu, J. Salvianolic acid A, a polyphenolic derivative from Salvia miltiorrhiza bunge, as a multifunctional agent for the treatment of Alzheimer’s disease. Mol. Divers. 2013, 17, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Link, C.D. Invertebrate models of Alzheimer’s disease. Genes Brain Behav. 2005, 4, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Diomede, L.; Rigacci, S.; Romeo, M.; Stefani, M.; Salmona, M. Oleuropein aglycone protects transgenic C. elegans strains expressing Aβ42 by reducing plaque load and motor deficit. PLoS One 2013, 8, e58893. [Google Scholar] [CrossRef] [PubMed]
- Diomede, L.; Cassata, G.; Fiordaliso, F.; Salio, M.; Ami, D.; Natalello, A.; Doglia, S.M.; De Luigi, A.; Salmona, M. Tetracycline and its analogues protect Caenorhabditis elegans from β amyloid-induced toxicity by targeting oligomers. Neurobiol. Dis. 2010, 40, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Beeg, M.; Diomede, L.; Stravalaci, M.; Salmona, M.; Gobbi, M. Novel approaches for studying amyloidogenic peptides/proteins. Curr. Opin. Pharmacol. 2013, 13, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Stoilova, T.; Colombo, L.; Forloni, G.; Tagliavini, F.; Salmona, M. A new face for old antibiotics: Tetracyclines in treatment of amyloidoses. J. Med. Chem. 2013, 56, 5987–6006. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yu, H.; Li, W.M.; Cheung, M.C.; Pang, Y.P.; Gu, Z.M.; Chan, K.; Wang, Y.T.; Zuo, Z.; Han, Y.F. Preclinical characterization of intestinal absorption and metabolism of promising anti-Alzheimer’s dimer bis(7)-tacrine. Int. J. Pharm. 2008, 357, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.Y.; Yu, Z.L.; Dong, H.; Lee, N.T.K.; Wang, H.; Fong, W.F.; Han, Y.F.; Ko, K.M. Evaluation of acute bis(7)-tacrine treatment on behavioral functions in 17-day-old and 30-day-old mice, with attention to drug toxicity. Pharmacol. Biochem. Behav. 2007, 86, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; He, L.; Mak, M.; Han, Y.; Ho, C.-Y.; Zuo, Z. Synthesis, biological activity, and biopharmaceutical characterization of tacrine dimers as acetylcholinesterase inhibitors. Int. J. Pharm. 2014, 477, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wu, Z.; Butko, P.; Christen, Y.; Lambert, M.P.; Klein, W.L.; Link, C.D.; Luo, Y. Amyloid-β-induced pathological behaviors are suppressed by Ginkgo biloba extract EGb 761 and ginkgolides in transgenic Caenorhabditis elegans. J. Neurosci. 2006, 26, 13102–13113. [Google Scholar] [CrossRef] [PubMed]
- Styren, S.D.; Hamilton, R.L.; Styren, G.C.; Klunk, W.E. X-34, a fluorescent derivative of Congo red: A novel histochemical stain for Alzheimer’s disease pathology. J. Histochem. Cytochem. 2000, 48, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Havas, D.; Hutter-Paier, B.; Ubhi, K.; Rockenstein, E.; Crailsheim, K.; Masliah, E.; Windisch, M. A longitudinal study of behavioral deficits in an AβPP transgenic mouse model of Alzheimer’s disease. J. Alzheimers Dis. 2011, 25, 231–243. [Google Scholar] [PubMed]
- Sample Availability: Samples of the compound AVCRI104P4 are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sola, I.; Viayna, E.; Gómez, T.; Galdeano, C.; Cassina, M.; Camps, P.; Romeo, M.; Diomede, L.; Salmona, M.; Franco, P.; et al. Multigram Synthesis and in Vivo Efficacy Studies of a Novel Multitarget Anti-Alzheimer’s Compound. Molecules 2015, 20, 4492-4515. https://doi.org/10.3390/molecules20034492
Sola I, Viayna E, Gómez T, Galdeano C, Cassina M, Camps P, Romeo M, Diomede L, Salmona M, Franco P, et al. Multigram Synthesis and in Vivo Efficacy Studies of a Novel Multitarget Anti-Alzheimer’s Compound. Molecules. 2015; 20(3):4492-4515. https://doi.org/10.3390/molecules20034492
Chicago/Turabian StyleSola, Irene, Elisabet Viayna, Tània Gómez, Carles Galdeano, Matteo Cassina, Pelayo Camps, Margherita Romeo, Luisa Diomede, Mario Salmona, Pilar Franco, and et al. 2015. "Multigram Synthesis and in Vivo Efficacy Studies of a Novel Multitarget Anti-Alzheimer’s Compound" Molecules 20, no. 3: 4492-4515. https://doi.org/10.3390/molecules20034492