
Phaleria macrocarpa Boerl. (Thymelaeaceae) Leaves Increase SR-BI Expression and Reduce Cholesterol Levels in Rats Fed a High Cholesterol Diet

Abstract
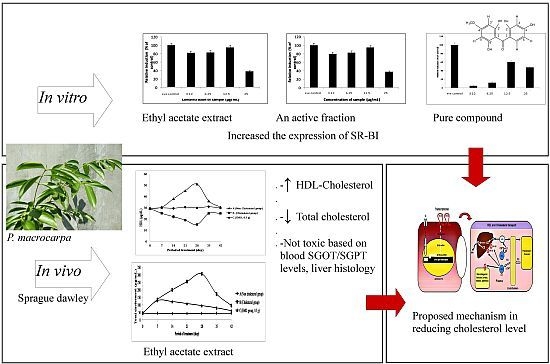
:
1. Introduction
2. Results and Discussion
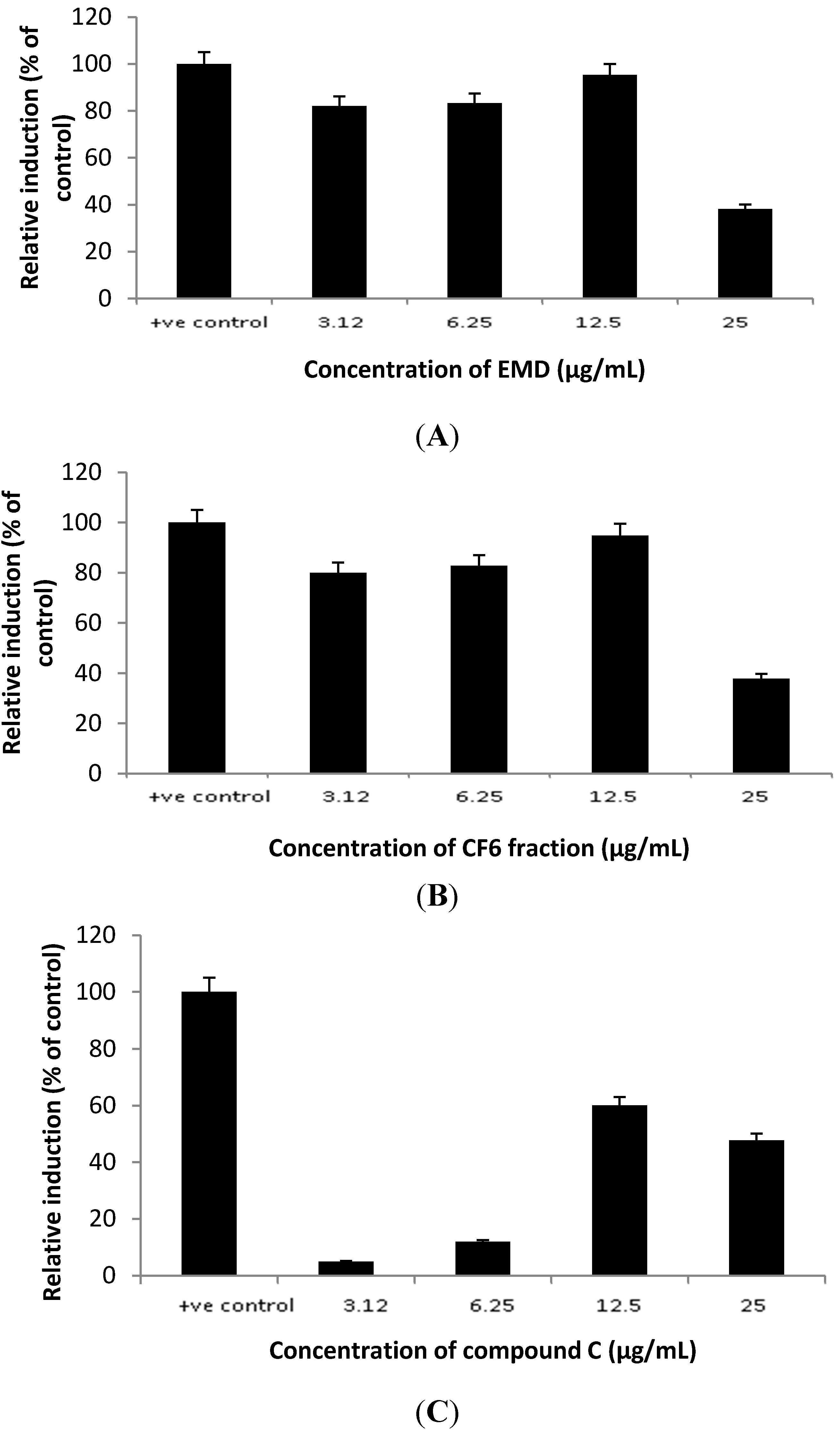
2.1. Cytotoxicity Property
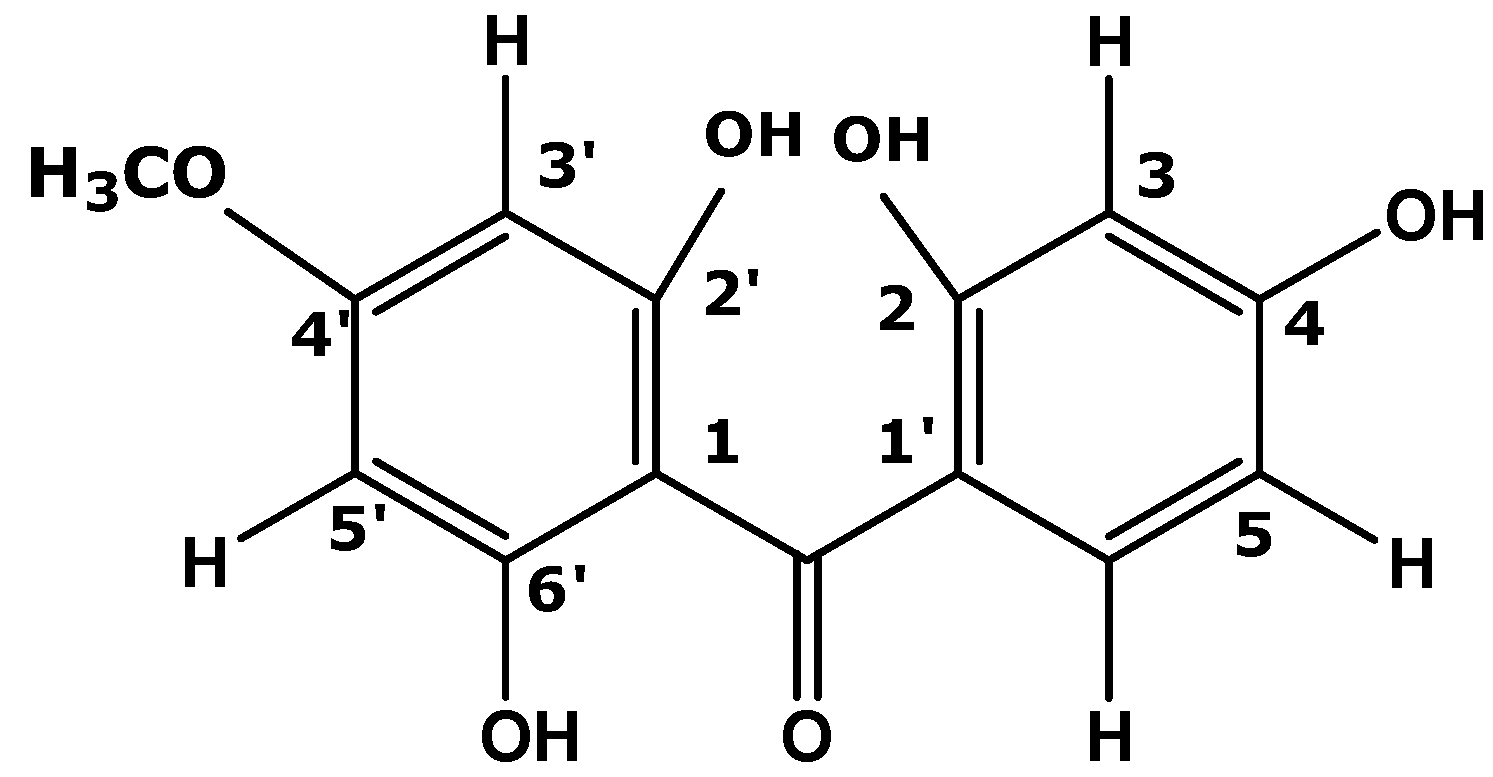
2.2. Fractionation, Compound Isolation, and Structure Elucidation of the Active Compound Isolated from P. macrocarpa Leaves

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound C from leaves of P. macrocarpa | |||
|---|---|---|---|
| Position -C | 13C-NMR (δ, ppm) | Position -H | 1H-NMR (δ, ppm, J = Hz) |
| -C=O | 196.94 | - | - |
| C-2' | 165.27 | - | - |
| C-6' | 161.62 | - | - |
| C-4 | 161.13 | - | - |
| C-1 | 132.21 | - | - |
| C-3 | 131.60 | H-3 | 7.650 (d, 8.8) |
| C-5 | H-5 | ||
| C-2 | 114.36 | H-2 | 6.887 (d, 8.8) |
| C-6 | H-6 | ||
| C-1' | 105.58 | - | - |
| C-3' | 93.64 | H-3' | 6.061 (s) |
| C-5' | H-5' | ||
| C-4'OCH3 | 54.88 | H-4' | 3.819 (s) |
| -OH | 9.854 | ||
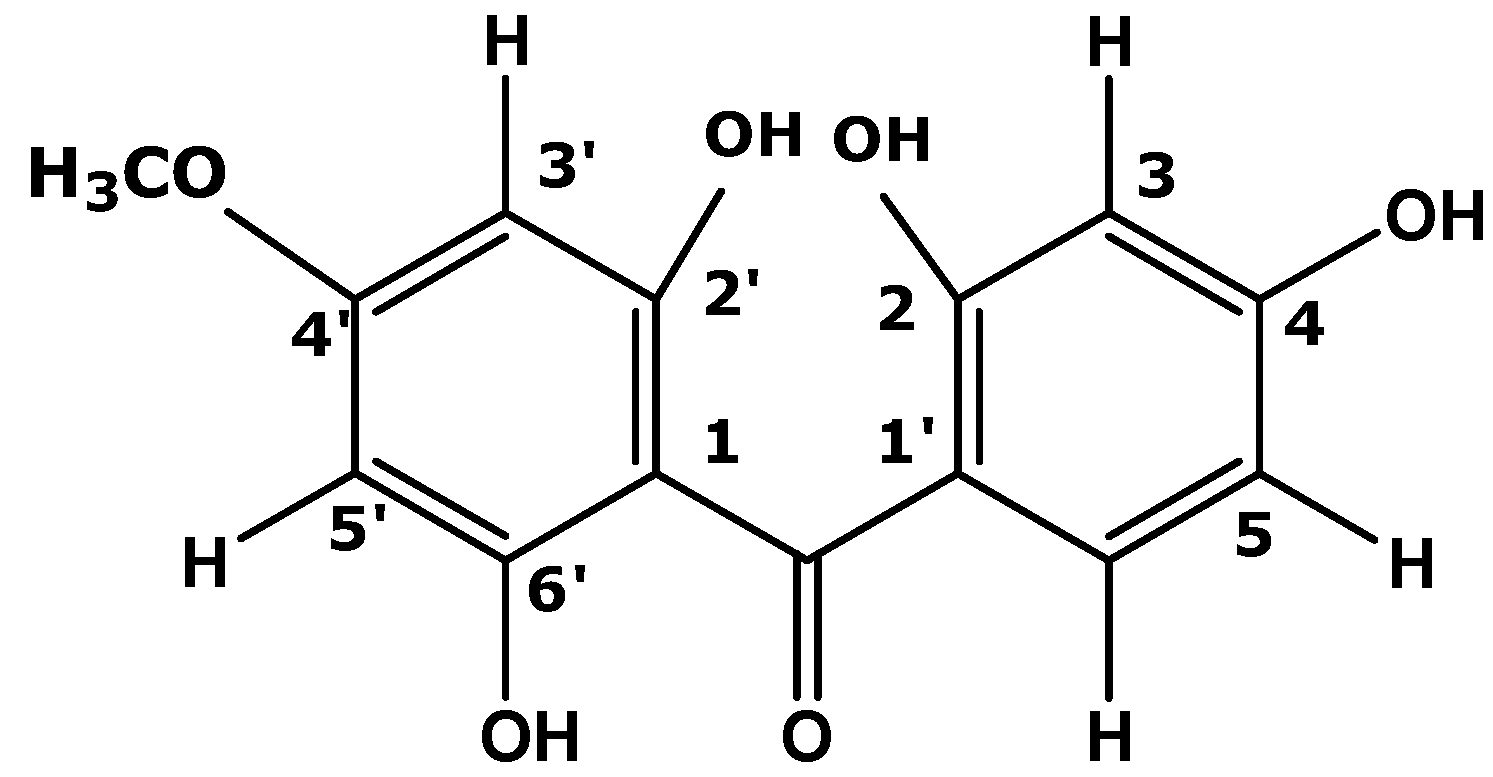
2.3. In Vitro Study: Mechanism Evaluation of P. macrocarpa Leaves Extract in Reducing Cholesterol Level by Looking the Expression of SR-B1 Genes
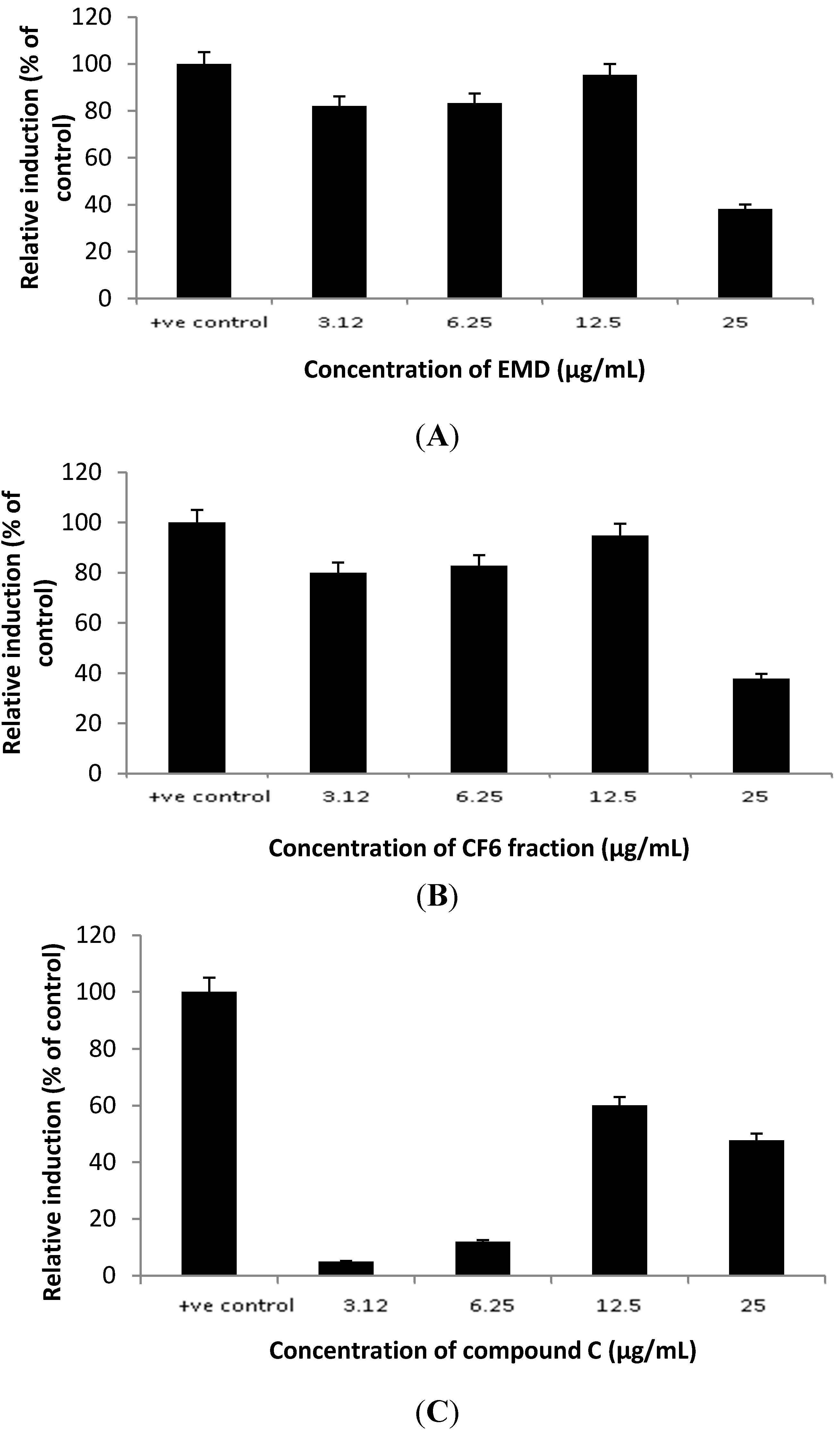
2.4. In Vivo Study: Hypocholesterolemia Effect of P. macrocarpa Leaves in Hypercholesterolemia-Induced Sprague Dawley Rats
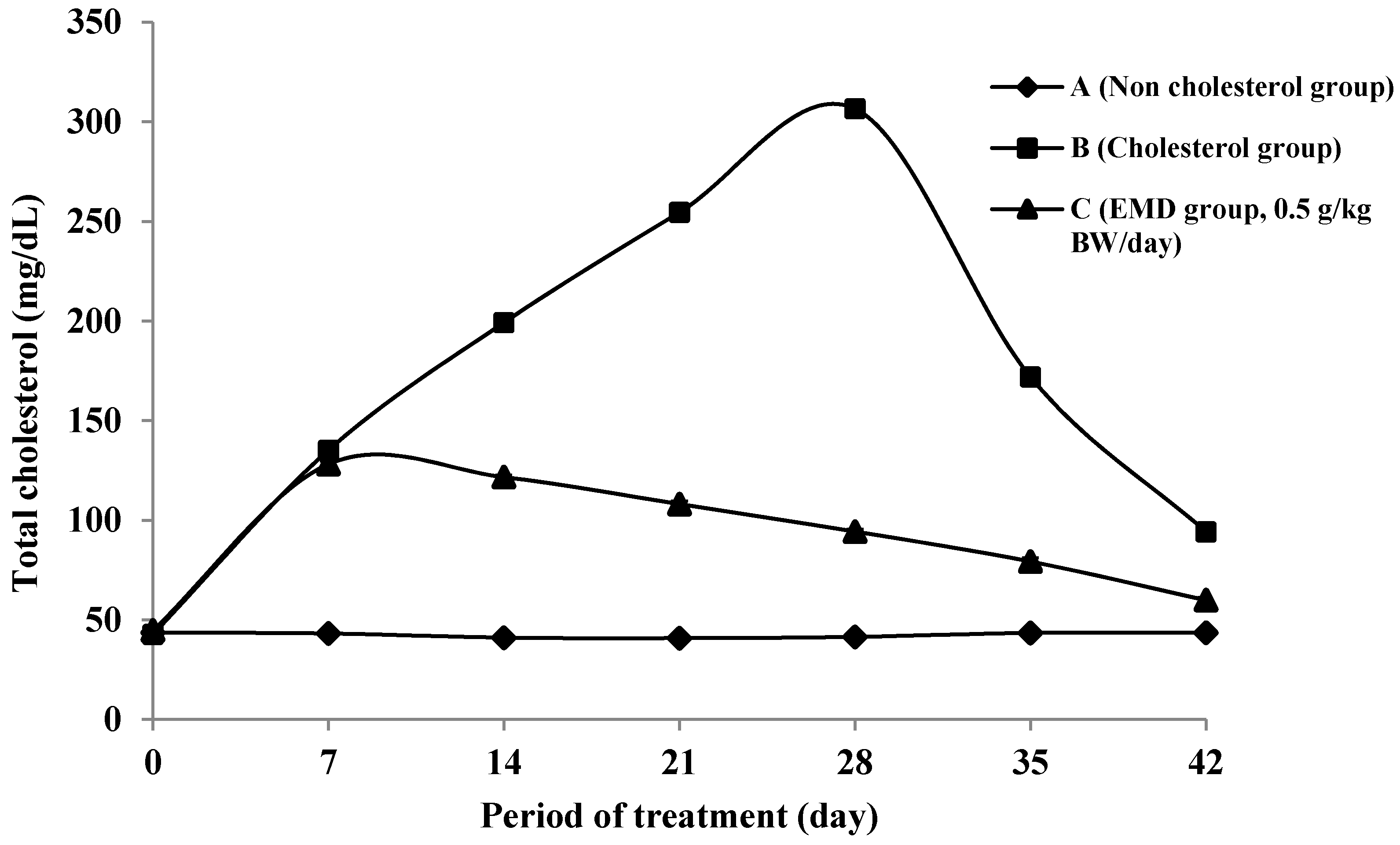
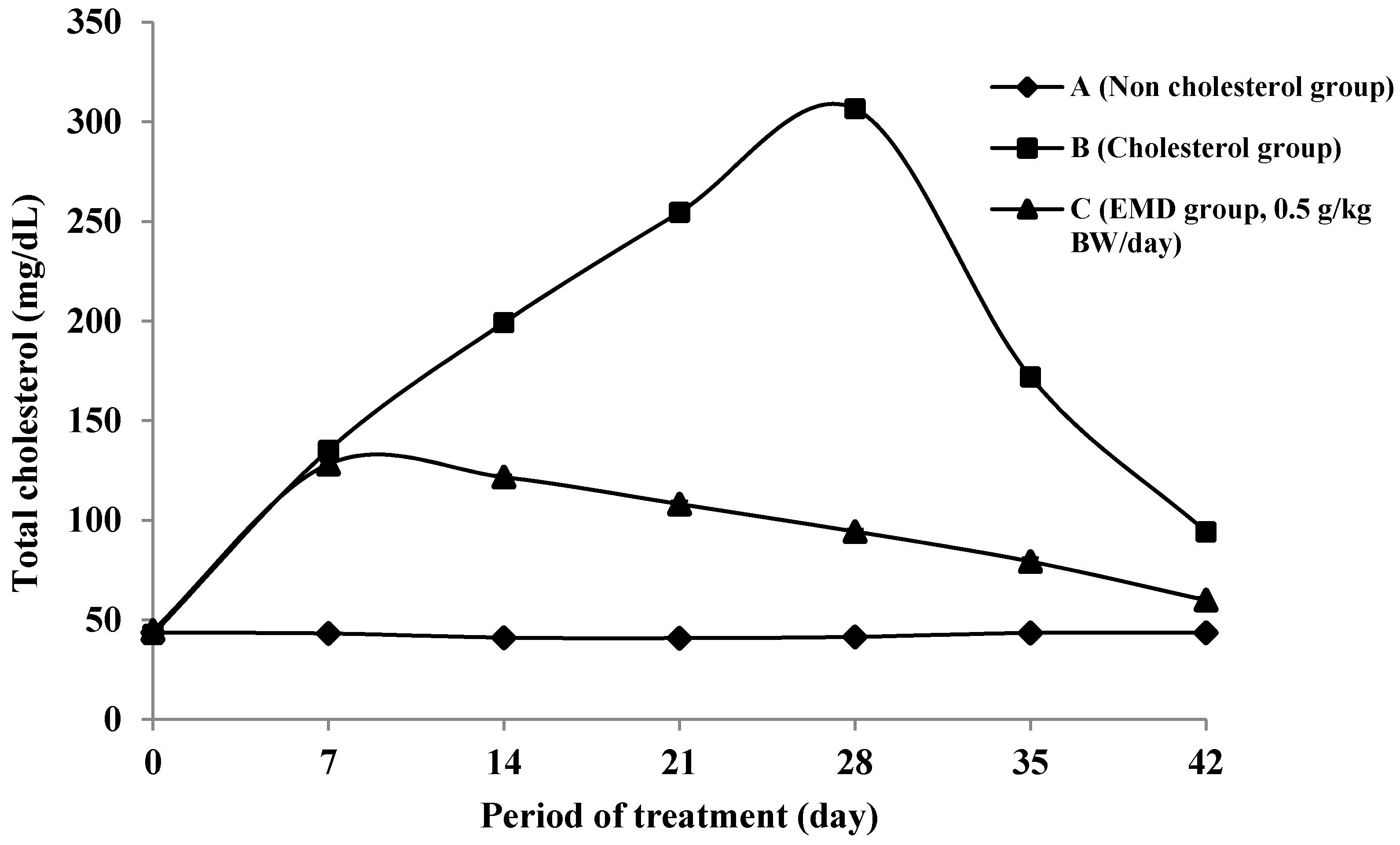
2.4.1. Total Cholesterol Levels
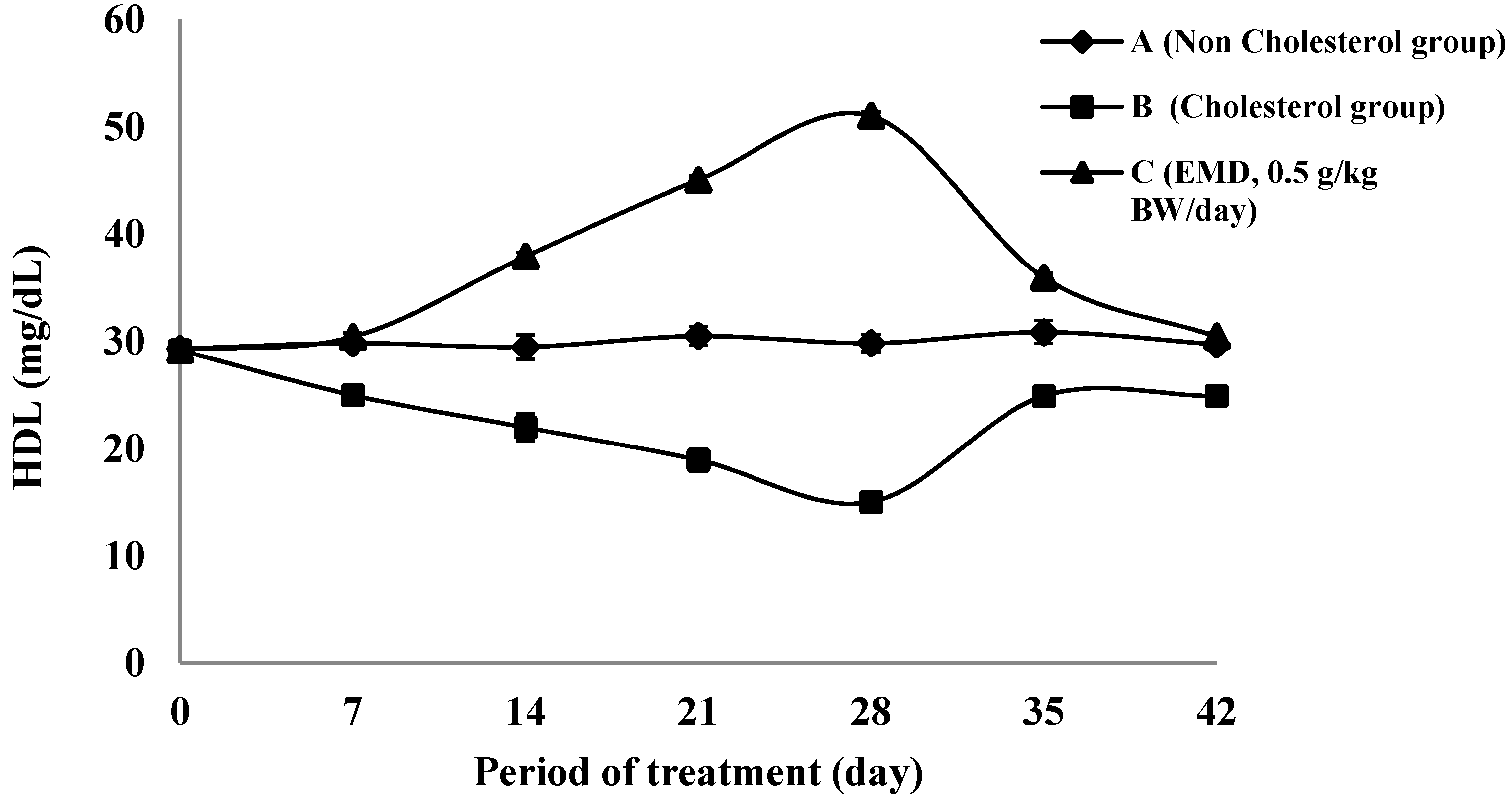
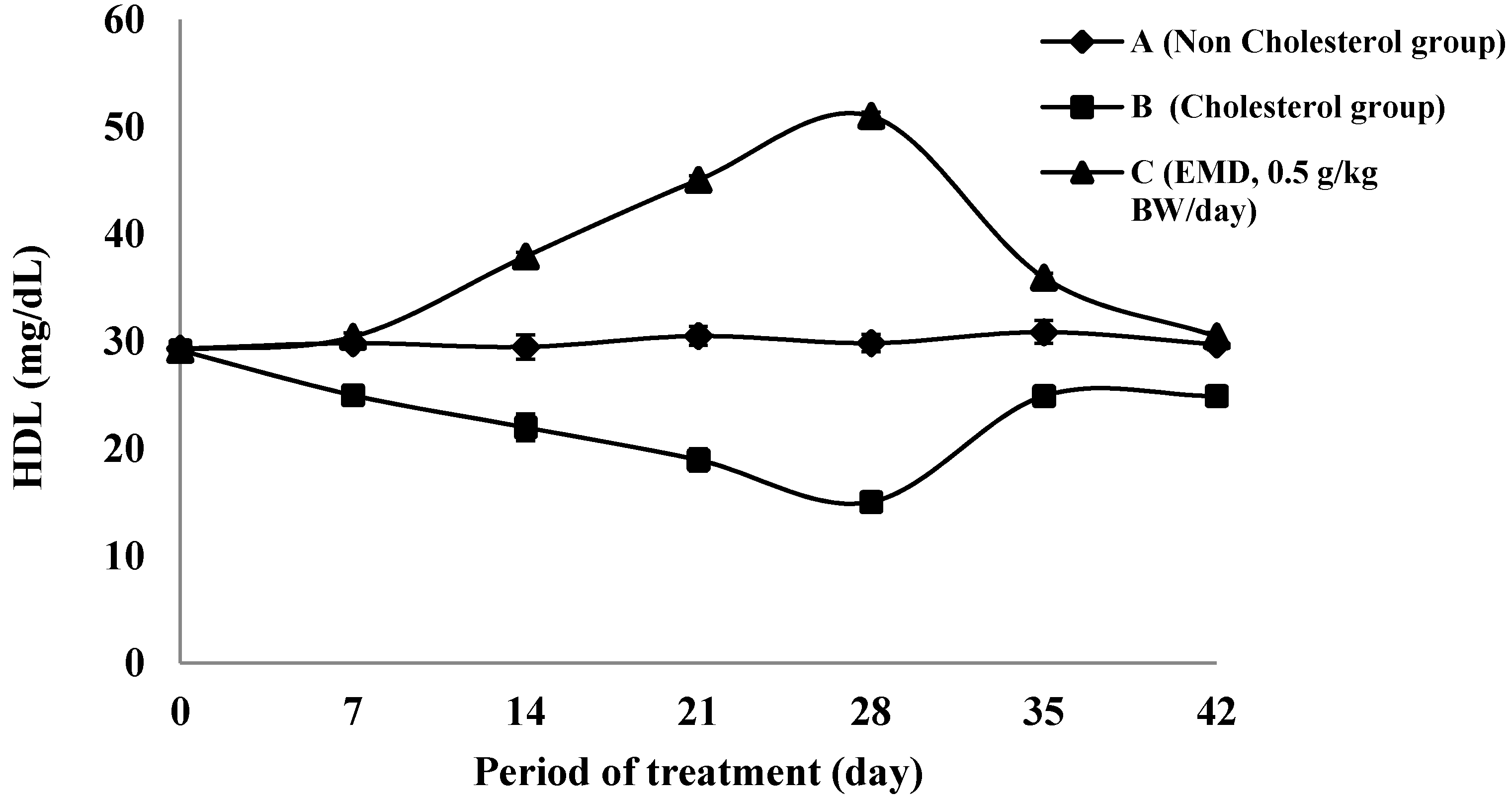
2.4.2. The HDL Cholesterol Level
2.4.3. SGOT and SGPT Activity
| Group | Activity of SGOT (U/I) for every 7 days during Treatment | ||||||
| 0 | 7 | 14 | 21 | 28 | 35 | 42 | |
| (A) | 131.73 ± 0.89 | 133.71 ± 1.32 | 133.45 ± 0.93 | 134.20 ± 0.95 | 133.86 ± 0.85 | 133.18 ± 1.26 | 133.62 ± 1.05 |
| (B) | 134.32 ± 1.14 | 146.44 ± 0.77 * | 157.84 ± 1.12 * | 192.73 ± 0.59 * | 236.73 ± 0.94 * | 179.18 ± 0.68 * | 145.51 ± 0.80 * |
| (C) | 134.65 ± 0.68 | 133.69 ± 0.97 | 133.10 ± 1.46 | 133.50 ± 0.96 | 136.51 ± 0.91 | 131.94 ± 1.20 | 132.50 ± 1.63 |
| Activity of SGPT (U/I) at every 7 days during Treatment | |||||||
| Group | 0 | 7 | 14 | 21 | 28 | 35 | 42 |
| (A) | 64.46 ± 1.19 | 63.95 ± 1.61 | 64.73 ± 1.23 | 64.96 ± 1.18 | 64.85 ± 1.12 | 65.15 ± 0.94 | 64.34 ± 0.53 |
| (B) | 64.66 ± 0.77 | 97.32 ± 1.66 * | 138.25 ± 0.71 * | 175.53 ± 0.63 * | 222.991 ± 0.76 * | 130.57 ± 1.22 * | 75.57 ± 2.43 |
| (C) | 64.87 ± 0.86 | 65.98 ± 0.76 | 66.46 ± 0.94 | 64.95 ± 1.31 | 65.04 ± 1.28 | 64.87 ± 0.89 | 64.87 ± 0.96 |
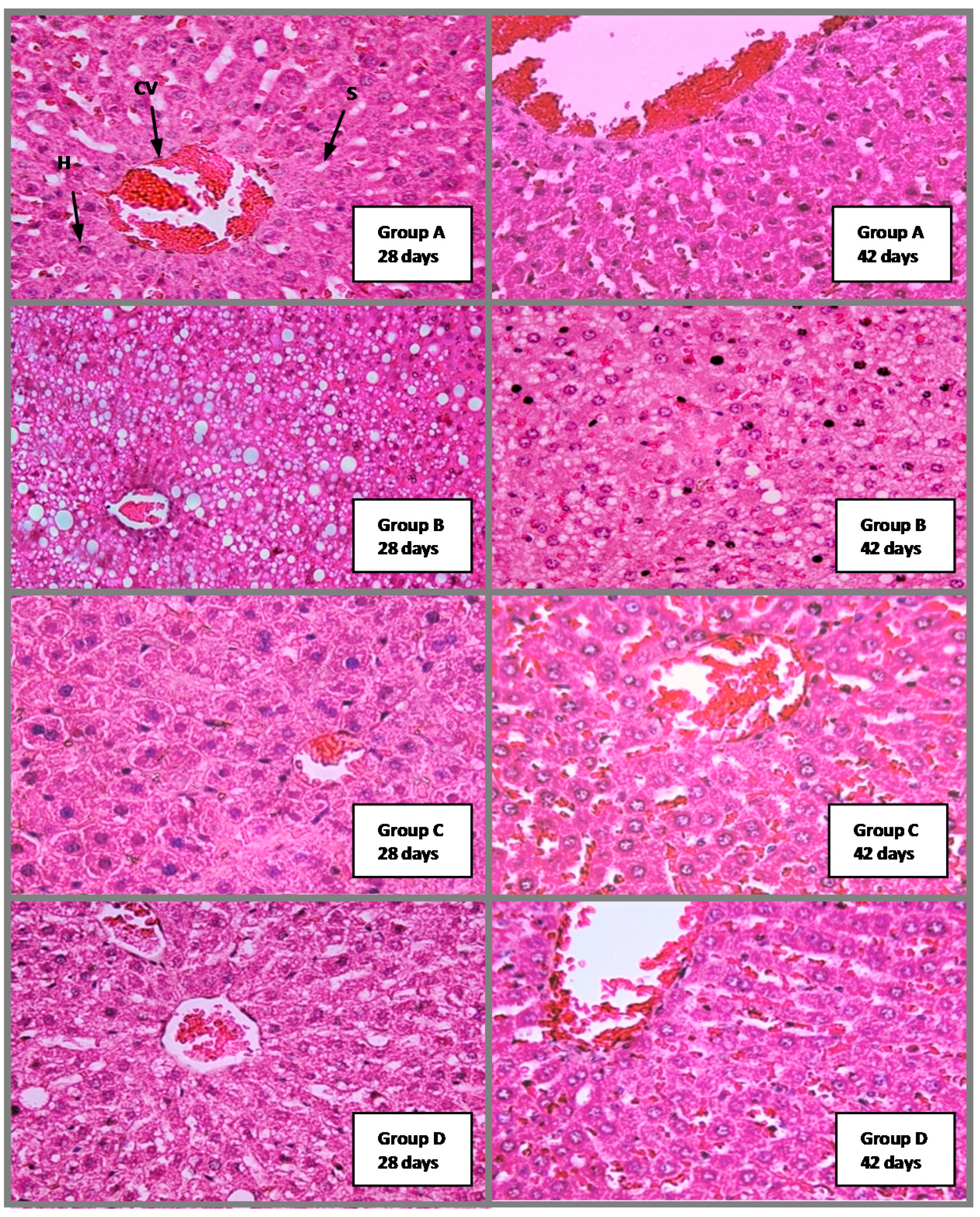
2.4.4. Histological Analysis

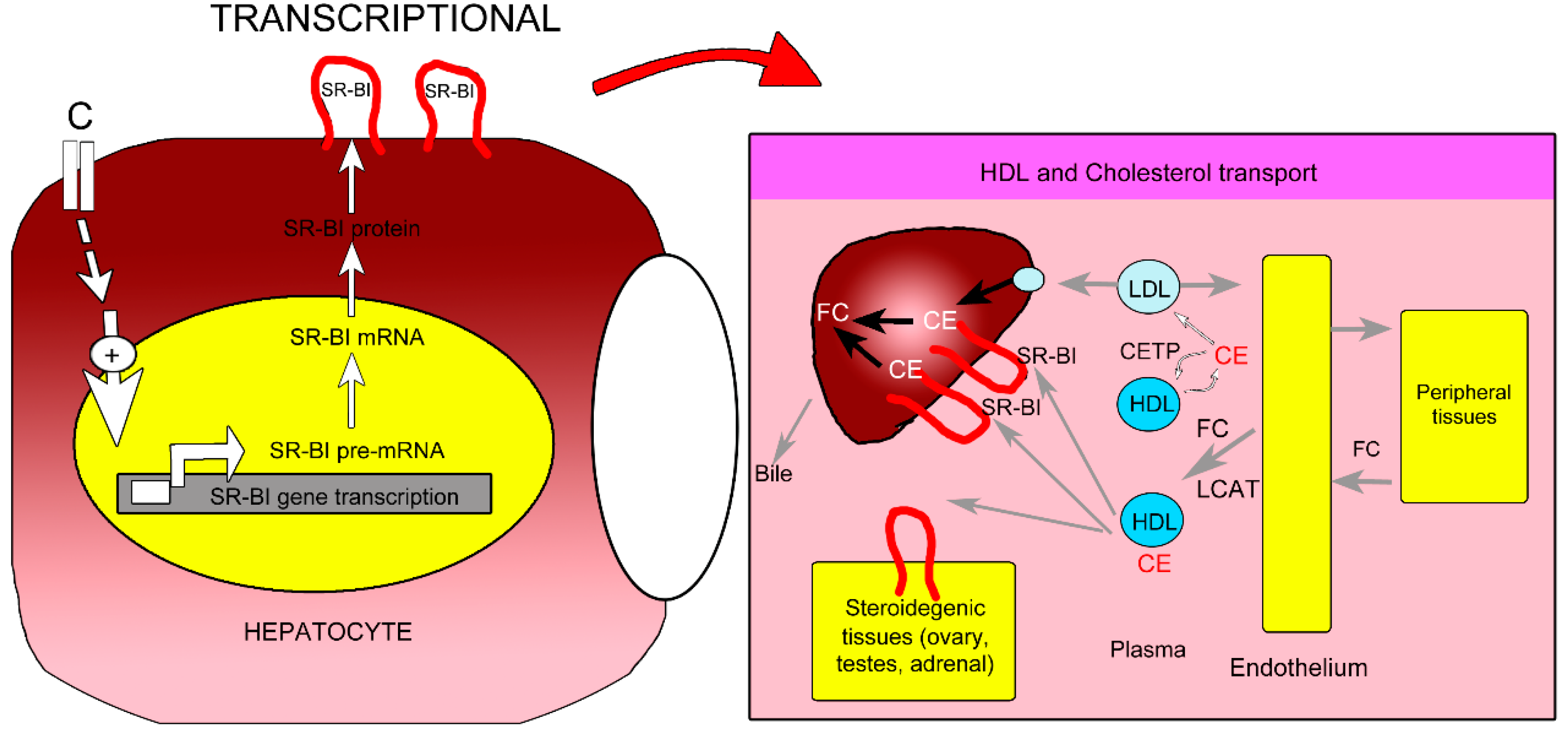
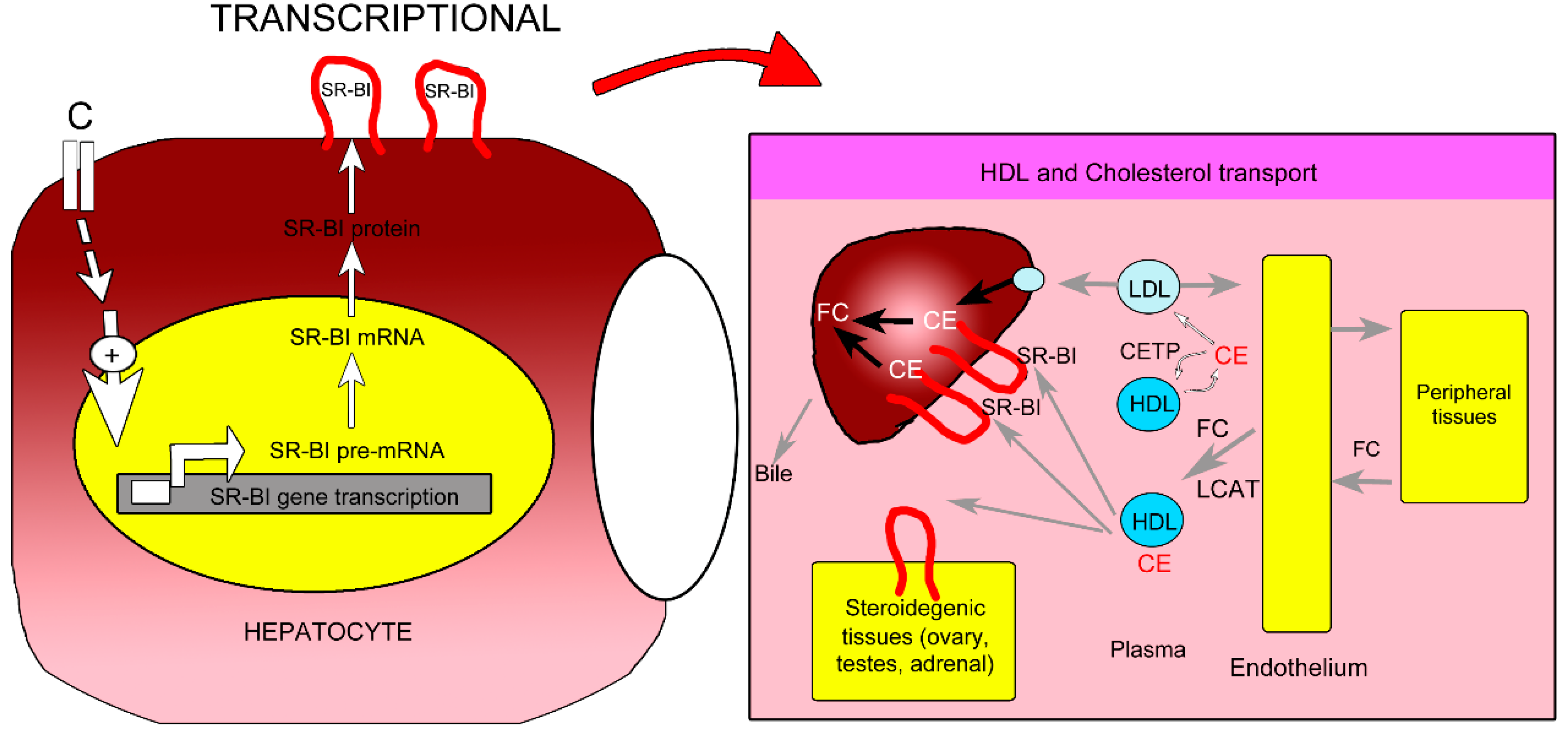
2.5. Proposed Mechanism of P. macrocarpa Leaves Effect in Reducing Cholesterol Levels (Hypocholesterolemia)
3. Experimental Section
3.1. P. macrocarpa Leave Materials
3.2. Extraction and Fractionations of P. Macrocarpa Leaves
3.3. Isolation and Structure Elucidation
3.4. In Vitro Study: Mechanism Evaluation of P. macrocarpa Leaves in Reducing Cholesterol Level by Looking the Expression of SR-BI Genes
3.4.1. Cytotoxicity Study by MTT Assay
3.4.2. Bioassay of P. macrocarpa Leaves against the Therapeutic Target for Hypercholesterolemia: SR-BI Promoter Trancriptional Activity Assay
3.4.3. Transfection of the Chimeric Reporter Plasmid into the HepG2 Cell Line
3.4.4. Treatment of the Transfected Cells with Sample
3.5. In Vivo Study: Hypocholesterolemia Effect of P. macrocarpa Leaves in Hypercholesterolemia-Induced Sprague Dawley Rats
3.5.1. Total Cholesterol Level (CHOD-PAP Method)
3.5.2. HDL Cholesterol Level
3.5.3. SGOT and SGPT Activity
3.5.4. Histological Analysis
3.6. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Salter, A.M.; Hayashi, R.; Al-seeni, M.; Brown, N.F.; Bruce, J.; Sorensen, O.; Atkinson, E.A.; Middleton, B.; Bleackey, R.C.; Brindley, D.N. Effect of hypothyroidism and high-fat feeding on mRNA concentration for the low density lipoprotein receptor and on acyl cholesterol acyl transferase activities in rat liver. J. Biochem. 1991, 276, 825–832. [Google Scholar]
- Ahmed, R.A.M.; Murao, K.; Imachi, H.; Yu, X.; Li, J.; Wong, N.C.; Ishida, T. Human scavenger receptor class b type 1 is regulated by activators of peroxisome proliferators-activated receptor gamma in hepatocytes. Endocrine 2009, 35, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Garcia, C.K.; Wyne, K.L.; Schultz, R.A.; Parker, K.L.; Hobbs, H.H. Structure and localization of the human gene encoding SR-BI/CLA-1. J. Biol. Chem. 1997, 272, 33068–33076. [Google Scholar] [CrossRef] [PubMed]
- Savel, J.; Lafitte, M.; Pucheu, Y.; Pradeau, V.; Tabarin, A.; Couffinhal, T. Very low levels of HDL cholesterol and atherosclerosis, a variable relationship-are view of LCAT deficiency. Vasc. Health Risk Manag. 2012, 6, 357–361. [Google Scholar]
- Acton, S.L.; Kozarsky, K.F.; Rigotti, A. The HDL receptor SR-BI: A new therapeutic target for atherosclerosis. Mol. Med. Today 1999, 5, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, A.; Krieger, M. Getting a handle on “Good” cholesterol with the high-density lipoprotein receptor. N. Engl .J. Med. 1999, 341, 2011–2013. [Google Scholar] [CrossRef] [PubMed]
- Altaf, R.; Asmawi, M.Z.; Dewa, A.; Sadikun, A.; Umar, M.I. Phytochemistry and medicinal properties of Phaleria macrocarpa (scheff). Boerl extract. Pharmacogn. Rev. 2013, 7, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Sumastuti, R.; Sonlimar, M. Cytotoxic effect of Phaleria macrocarpa (Scheff.) boerl fruits and leaves to hela cell. Medika 2002, 2, 773–777. [Google Scholar]
- Mae, S.H.W.; Sofia, M.; Ibnu, G.G.; Mark, T.H.; Rao, K.V.; Subagus, W. Phalerin, a new benzophenoic glycoside isolated from the methanolic extract of Phaleria macrocarpa (scheff). Boerl Leaves. Maj. Farm. Indones. 2005, 16, 51–57. [Google Scholar]
- Lisdawati, V.; Kardono, L.B.S. Antioxidant activity of various extract fractions of Phaleria macrocarpa (Scheff.) Boerl. fruit and mesocarp. Media Litbang Kesehatan 2006, XIV, 1–7. [Google Scholar]
- Harahap, Y.; Syafhan, N.F.; Karsono, B. In vitro cytotoxicity test of Mahkota Dewa fruit dosage form on MCF-7 cell. Jurnal Bahan Alam Indonesia 2007, 6, 55–59. [Google Scholar]
- Zhang, S.-Y.; Zhang, Q.-H.; Zhao, W.; Zhang, X.; Zhang, Q.; Bi, Y.-F.; Zhang, Y.-B. Isolation, characterization and cytotoxic activity of benzophenone glucopyranosides from Mahkota Dewa (Phaleria macrocarpa (Scheff.) Boerl). Bioorganic Med. Chem. Lett. 2012, 22, 6862–6866. [Google Scholar] [CrossRef]
- Fitriani, D. Antihypocholesterolemia Screening Activity of Water Fruit Extract of Phaleria macrocarpa (Scheff.) Boerl in Mus muscullus. Undergraduate Thesis, Intitut Teknologi Bandung, Bandung, Indonesia, 2010. [Google Scholar]
- Sugiwati, S.; Leonardus, B.S.; Bintang, M. α-Glucosidase inhibitory activity and hypoglycemic effect of Phaleria macrocarpa fruit pericarp extracts by oral administration to rats. J. Appl. Sci. 2006, 6, 2312–2316. [Google Scholar] [CrossRef]
- Triastuti, A.; Choi, J.W. Protective effect of ethylacetate of Phaleria macrocarpa (Scheff.) Boerl. on oxidative stress associated with alloxan-induced diabetic rats. Jurnal Ilmiah Farmasi 2008, 5, 9–17. [Google Scholar]
- Nor Fariza, I.; Fadzurena, J.; Zunoliza, A.; Luqman Chuah, A.; Pin, K.Y.; Adawiah, I. Anti-inflammatory activity of the major compound from methanol extract of Phaleria macrocarpa leaves. J. Appl. Sci. 2012, 12, 1195–1198. [Google Scholar] [CrossRef]
- Ali, R.B.; Atangwho, I.J.; Kaur, N.; Abraika, O.S.; Ahmad, M.; Mahmud, R.; Asmawi, M.Z. Bioassay-guided anti-diabetic study of Phaleria macrocarpa fruit extract. Molecules 2012, 17, 4986–5002. [Google Scholar] [CrossRef] [PubMed]
- Kurniajasanti, R. Potensi Ekstrak Buah Mahkota Dewa (Phaleria Macrocarpa) Sebagai Antioksidan Dalam Pengaturan Profil Lipid Darah Mencit; Research Report; Faculty of Veterinary, AirLangga University: Surabaya, Indonesia, 21 March 2006. [Google Scholar]
- Chong, S.C.; Dollah, M.A.; Chong, P.P.; Maha, A. Phaleria macrocarpa (Scheff.) Boerl fruit aqueous extract enhances LDL receptor and PCSK9 expression in vivo and in vitro. J. Ethnopharmacol. 2011, 137, 817–827. [Google Scholar] [CrossRef] [PubMed]
- Aripin, A.; Poppy, F.A.; Raymond, R. Tjandrawinata. Isolate Compounds from Phaleria macrocarpa as Anti Cancer. WO2010064172 A2, 10 June 2010. [Google Scholar]
- Zhang, Y.-B.; Xu, X.-J.; Liu, H.-M. Chemical constituents from Mahkota dewa. J. Asian Nat. Prod. Res. 2006, 8, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Arnold, V.E.; Nofer, J.R.; Assmann, G. High density lipo protein and arteriosclerosis: Role of cholesterol efflux and reverse cholesterol transport. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Eckhardt, E.R.M.; Cai, L.; Sun, B.; Webb, N.R.; Deneys, R.W. High density lipoprotein uptake by scavenger receptor SRB1. J. Biol. Chem. 2004, 273, 4372–4381. [Google Scholar]
- Silver, D.L.; Wang, N.; Xiao, X.; Tall, A.R. High density lipoprotein (HDL) particle uptake mediated by scavenger receptor class B type I results in selective sorting of HDL cholesterol from protein and polarized cholesterol secretion. J. Biol. Chem. 2001, 276, 25287–25293. [Google Scholar] [CrossRef] [PubMed]
- Rothblatt, G.H. Cell cholesterol efflux: Integration of old and new observations provides new insights. J. Lipid Res. 1999, 40, 781–796. [Google Scholar] [PubMed]
- Kozarsky, K.F.; Donahee, M.H.; Ribotti, A.; Iqbal, S.N.; Edelman, E.R.; Krieger, M. Over expression of the HDL receptor SR-BI alters plasma HDL and bile cholesterol levels. Nature 1997, 387, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Arai, T.; Ji, Y.; Rinninger, F.; Tall, A.R. Liver-specific over expression of scavenger receptor BI decreases levels of very low density lipoprotein Apo B, low density lipoprotein Apo B, and high density lipoprotein in transgenic mice. J. Biol. Chem. 1998, 273, 32920–32926. [Google Scholar] [CrossRef] [PubMed]
- Ueda, Y.; Royer, L.; Gong, E.; Zhang, J.; Cooper, P.N.; Francone, O.; Rubi, E.M. Lower plasma levels and accelerated clearance of high density lipoprotein (HDL) and non-HDL cholesterol in scavenger receptor class B type I transgenic mice. J. Biol. Chem. 1999, 274, 7165–7171. [Google Scholar] [CrossRef] [PubMed]
- Andriani, Y.; Chew, G.-S.; Tengku Muhammad, T.S.; Habsah, M.; Latif, J.; Effendy, A.W.M. A new 4',6-dihydroxy-4-methoxybenzophenone-2-O-β-d-gentiobioside and 4',6-dihydroxy-4-methoxybenzophenone-2-O-β-d-glucoside isolated from methanolic extract of Phaleria macrocarpa (Scheff.) Boerl leaves. Int. J. Phytomedicine 2014, 6, 63–72. [Google Scholar]
- Malole, M.B.M.; Pramono, S.U. Use Animal Research in Laboratorium; Intercollegiate Center, Institut Pertanian Bogor: Bogor, Indonesia, 1989. [Google Scholar]
- [KKI] Kelompok Kerja Ilmiah Phyto Medica. Pedoman Pemeriksaan dan Pengembangan Fitofarmaka, lembaga Pengembangan Bahan Alami; PhytoMedica: Jakarta, Indonesia, 1993. [Google Scholar]
- Grundy, S.M. Multi factorial etiology of hypercholesterolemia: Implication for prevention of coronary heart disease. Arterioscler. Thromb. 1991, 11, 1619–1635. [Google Scholar] [CrossRef] [PubMed]
- Chambers, C.M.; Ness, G.C. Dietary cholesterol regulates hepatic 3-hydroxy-3-methyl glytaril coenzyme A reductase gene expression in rats primarily at the level of translation. Arch. Biochem. Biophys. 1998, 354, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.K.; Granner, D.K.; Mayes, P.A.; Rodwell, V.W. Biokimia Harper, 24th ed.; Hartono, A., Santoso, A.H., Eds.; Appleton & Lange: Jakarta, Indonesia, 1996. [Google Scholar]
- Trigatti, B.L.; Krieger, M.; Rigotti, A. Influence of the HDL receptor SR-BI on lipoprotein metabolism and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1732–1738. [Google Scholar] [CrossRef] [PubMed]
- Rigotti, A.; Miettinen, H.; Krieger, M. The role of the high density lipoprotein receptor SR-BI in the lipid metabolism of endocrine and other tissues. Endocr. Rev. 2003, 24, 357–387. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Wang, N.; Ramakrishnan, R.; Sehayek, E.; Huszar, D.; Breslow, J.L.; Tall, A.R. Hepatic scavenger receptor BI promotes rapid clearance of high density lipoprotein free cholesterol and its transport into bile. J. Biol. Chem. 1999, 274, 33398–33402. [Google Scholar] [CrossRef] [PubMed]
- Sehayek, E.; Ono, J.G.; Shefer, S.; Nguyen, L.B.; Wang, N.; Batta, A.K.; Salen, G.; Smith, J.D.; Tall, A.R.; Breslow, J.L. Biliary cholesterol excretion: A novel mechanism that regulates dietary cholesterol absorption. Proc. Natl. Acad. Sci. USA 1998, 95, 10194–10199. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.J. Clicical Chemistry; Mosby: Edinburgh, UK, 2000. [Google Scholar]
- Girindra, A. Pathology of Biochemistry-Petunjuk Pratikum; PAU Ilmu Hayat, Bogor Agriculture Institute: Bogor, Indonesia, 1989. [Google Scholar]
- Karwani, G.; Sosodia, S. Hepato protective activity of Mimosa pudica Linn. In carbon tetra chloride induced hepatotoxicity in rats. J. Herb. Med. Toxicol. 2011, 5, 27–32. [Google Scholar]
- Rekha, R.; Hemalatha, S.; Akasakalai, K.; MadhuKrishna, C.H.; Sohil, V.B.; Sundaram, R.M. Hepatoprotective activity of Mimosa pudica leaves against Carbon tetra chloride inducedt oxicity. J. Nat. Prod. 2009, 2, 116–122. [Google Scholar]
- Ravikumar, S.; Gnanadesign, M. Hepatoprotective and antioxidant activity of a mangrove plant Lumnitzera racemosa. Asian Pac. J. Trop. Biomed. 2011, 1, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Jeong, W.-I.; Jeong, D.-H.; Do, S.-H.; Kim, Y.-K.; Park, H.-Y.; Kwon, O.-D.; Kim, T.-H.; Jeong, K.-S. Mild Hepatic fibrosis in cholesterol and sodium cholate diet-feed rats. J. Vet. Med. Sci. 2005, 67, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Buyssens, N.; Kockx, M.M.; Herman, A.G.; Lazou, J.M.; Vanden Berg, K.; Wisse, E.; Geerts, A. Control lobular liver fibrosis in the hipercholesterolemic rabbit. Hepatology 1996, 24, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A.; Rahemtulla, A.; Daly, T.; Khwaja, S.; Miao, L.; Zao, S.; Tahan, S.R. Cholesterol supplementation prevents necrosis and imflammation but enhances fibrosis in alcoholic liver disease in the rat. Hepatology 1997, 26, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.L.; Michael, M.C. Lehninger Principles of Biochemistry, 3rd Ed. ed; Worth Publisher: New York, NY, USA, 2000. [Google Scholar]
- Kobayashi, M.; Kagawa, T.; Takano, R.; Itagaki, S.; Hirano, T.; Iseki, K. Effect medium pH on the cytotoxicity of hydrophilic statins. J. Pharm. Pharm. Sci. 2007, 10, 332–339. [Google Scholar] [PubMed]
- Leiva, A.; Verdejoa, H.; Beníteza, M.L.; Martíneza, A.; Bussoa, D.; Rigotti, A. Mechanisms regulating hepatic SR-BI expression and their impact on HDL metabolism. Atherosclerosis 2011, 217, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Krieger, M. Charting the fate of the “good cholesterol”: Identification and characterization of the high-density lipoprotein receptor SR-BI. Annu. Rev. Biochem. 1999, 68, 523–558. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, J. The role of the exogenous pathway in hypercholesterolaemia. Eur. Heart J. Suppl. (Suppl. E) 2001, 3, E2–E5. [Google Scholar] [CrossRef]
- Andriani, Y.; Effendy, A.W.M.; Sifzizul, T.M.T.; Habsah, M. Antibacterial, radical scavenging activities and cytotoxicity properties of Phaleria macrocarpa (Scheff.) Boerl. Leaves in HepG2 cell lines. Int. J. Pharm. Res. 2011, 2, 1700–1706. [Google Scholar]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Rieser, M.J.; Gu, Z.M.; Fang, X.P.; Zeng, L.; Wood, K.V.; McLaughlin, J.L. Five novel mono-tetrahydrofuran ring acetogenins from the seeds of Annona muricata. J. Nat. Prod. 1996, 59, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Trinder, P. Recommended method for the determination of cholesterol and triglyceride in blood. Ann. Clin. Biochem. 1969, 6, 24–27. [Google Scholar] [CrossRef]
- IFCC (International Federation of Clinical Chemistry), Scientific comitte. Recommended method for the determination of GOT and GPT in blood. J. Clin. Chem. Biochem. 1980, 18, 521–534. [Google Scholar]
- Tuffery, A.A. Laboratory Animals: An Introduction for Experiment, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1995. [Google Scholar]
- Sample Availability: Samples of the compound P. macrocarpa are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andriani, Y.; Tengku-Muhammad, T.S.; Mohamad, H.; Saidin, J.; Syamsumir, D.F.; Chew, G.-S.; Abdul Wahid, M.E. Phaleria macrocarpa Boerl. (Thymelaeaceae) Leaves Increase SR-BI Expression and Reduce Cholesterol Levels in Rats Fed a High Cholesterol Diet. Molecules 2015, 20, 4410-4429. https://doi.org/10.3390/molecules20034410
Andriani Y, Tengku-Muhammad TS, Mohamad H, Saidin J, Syamsumir DF, Chew G-S, Abdul Wahid ME. Phaleria macrocarpa Boerl. (Thymelaeaceae) Leaves Increase SR-BI Expression and Reduce Cholesterol Levels in Rats Fed a High Cholesterol Diet. Molecules. 2015; 20(3):4410-4429. https://doi.org/10.3390/molecules20034410
Chicago/Turabian StyleAndriani, Yosie, Tengku Sifzizul Tengku-Muhammad, Habsah Mohamad, Jasnizat Saidin, Desy Fitrya Syamsumir, Guat-Siew Chew, and Mohd Effendy Abdul Wahid. 2015. "Phaleria macrocarpa Boerl. (Thymelaeaceae) Leaves Increase SR-BI Expression and Reduce Cholesterol Levels in Rats Fed a High Cholesterol Diet" Molecules 20, no. 3: 4410-4429. https://doi.org/10.3390/molecules20034410
APA StyleAndriani, Y., Tengku-Muhammad, T. S., Mohamad, H., Saidin, J., Syamsumir, D. F., Chew, G.-S., & Abdul Wahid, M. E. (2015). Phaleria macrocarpa Boerl. (Thymelaeaceae) Leaves Increase SR-BI Expression and Reduce Cholesterol Levels in Rats Fed a High Cholesterol Diet. Molecules, 20(3), 4410-4429. https://doi.org/10.3390/molecules20034410