
Immobilization of Cyclooxygenase-2 on Silica Gel Microspheres: Optimization and Characterization



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
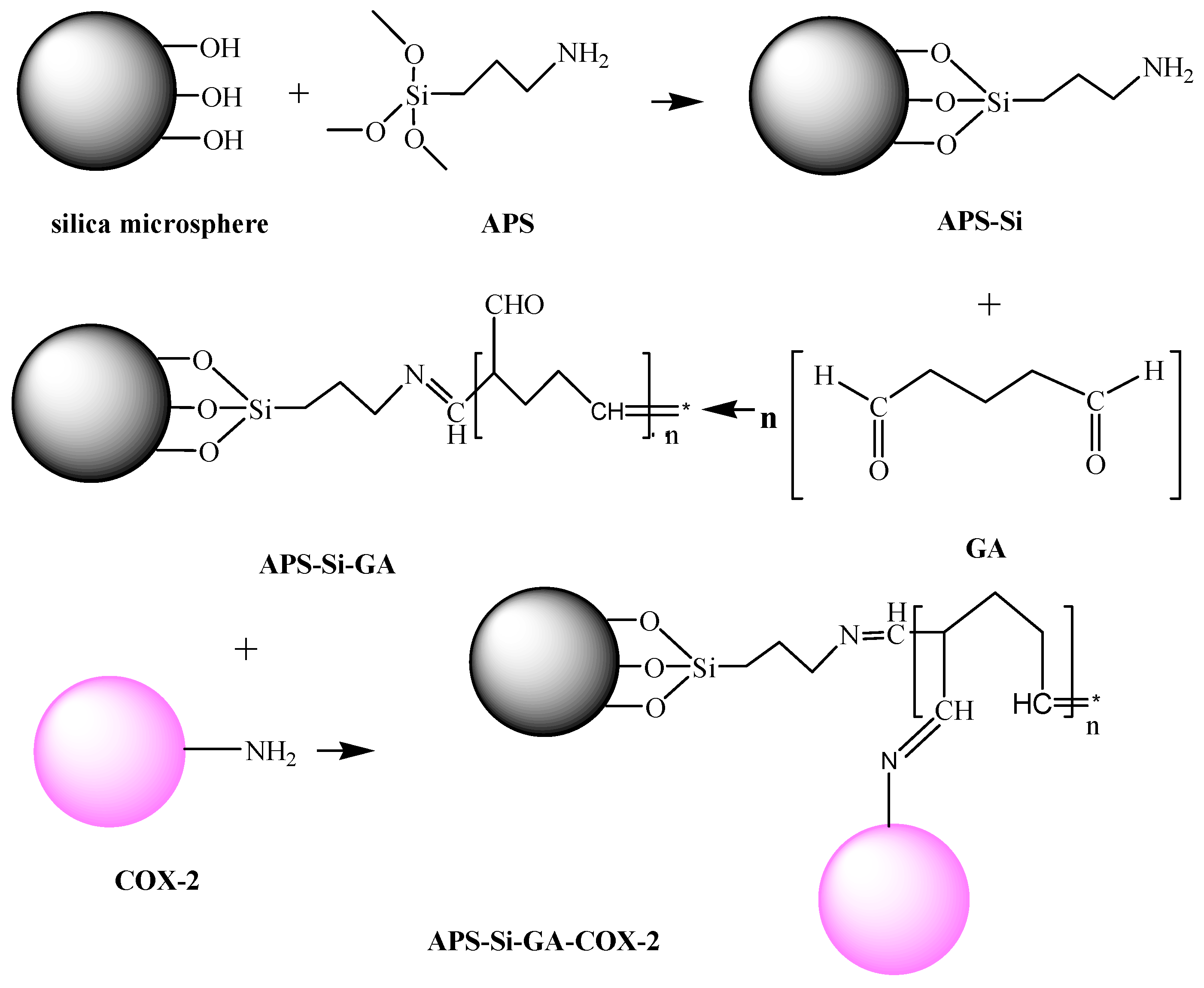{kind=link}
1. Introduction
2. Results and Discussion
2.1. Optimization of the Immobilization Conditions
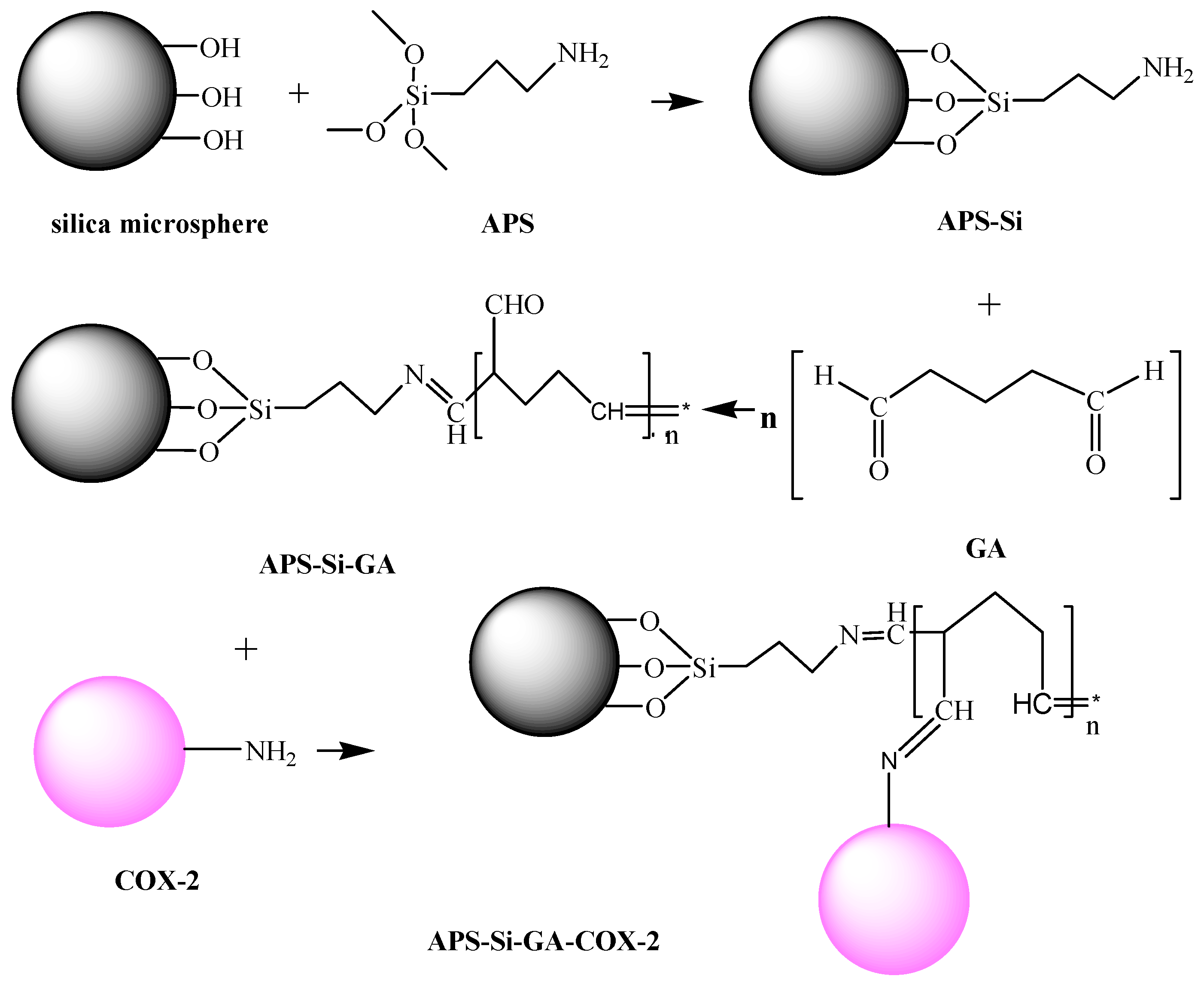
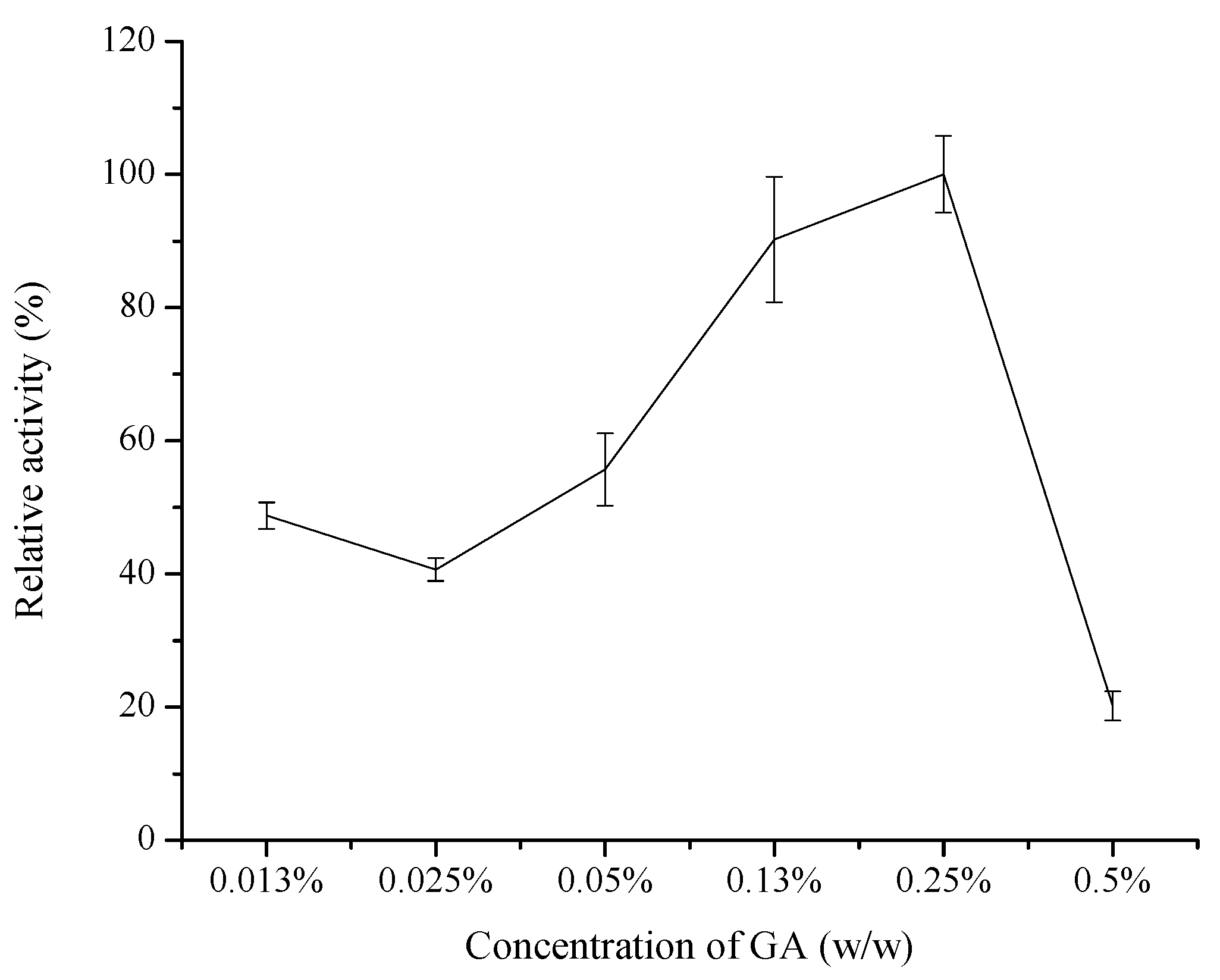
2.1.1. Effect of GA Concentration
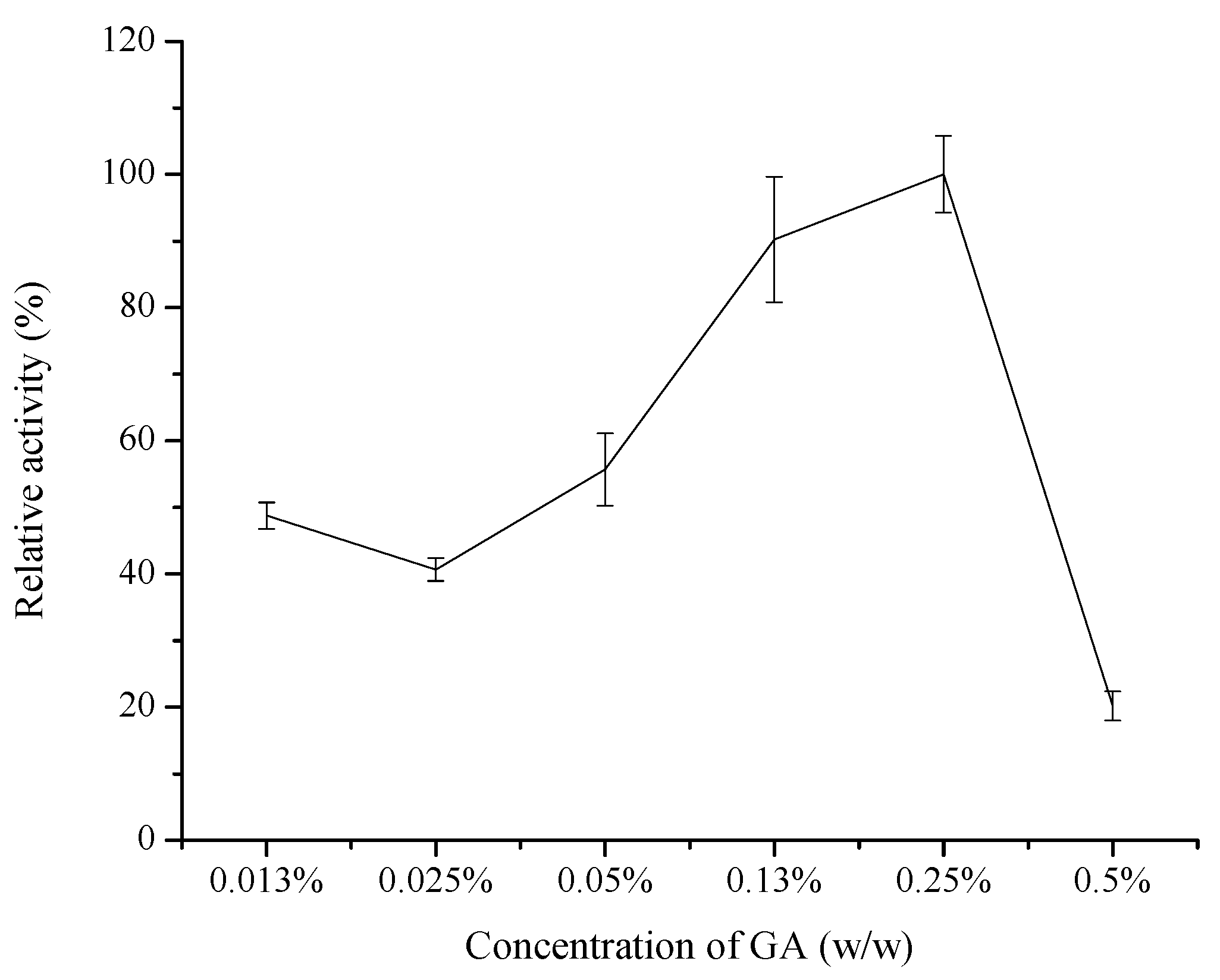
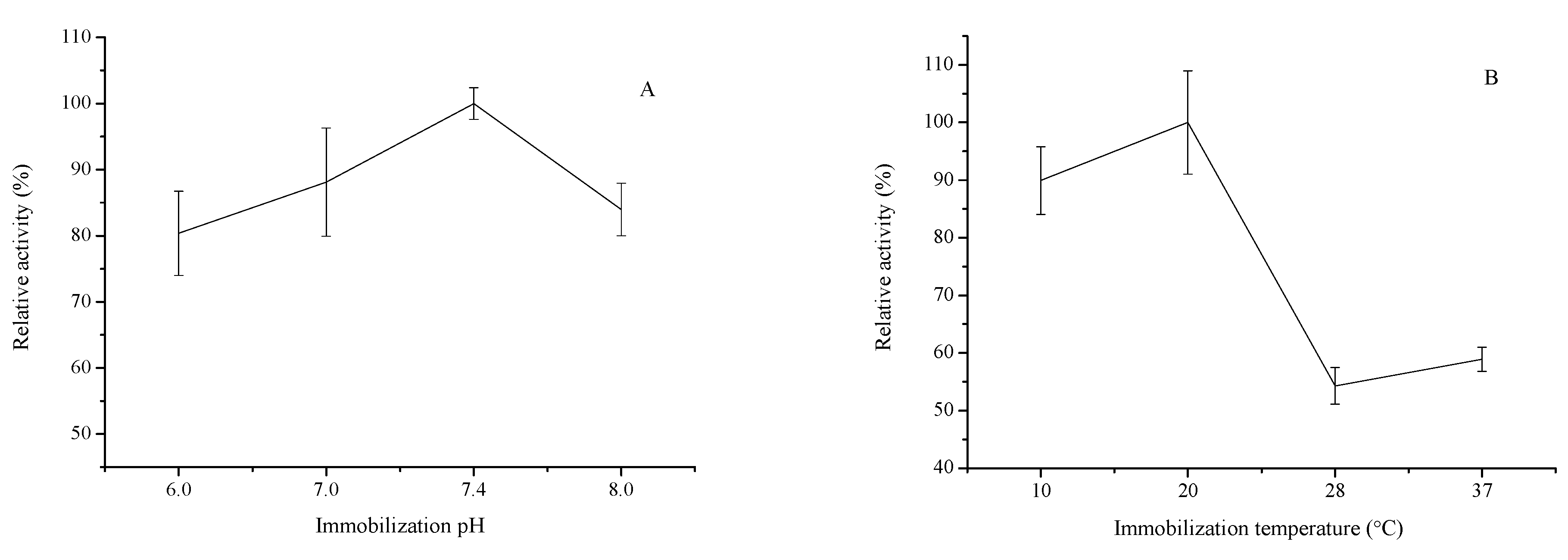
2.1.2. Effect of Reaction Solution pH
2.1.3. Effect of Reaction Temperature and Time
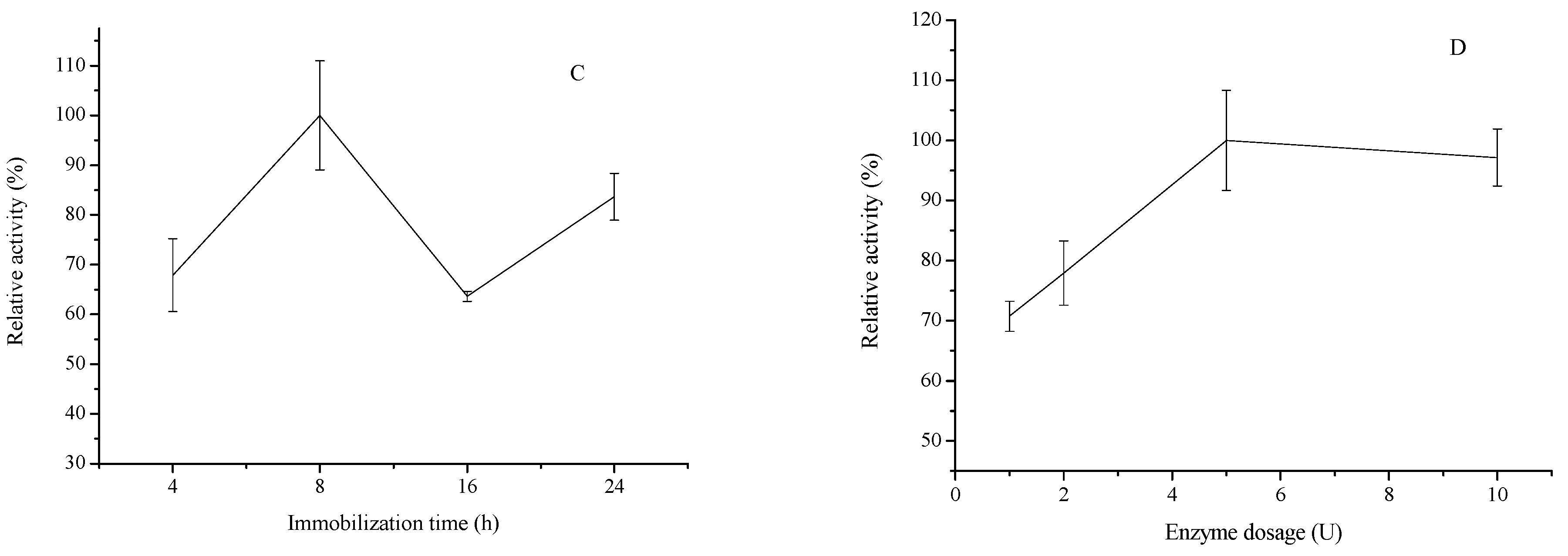
2.1.4. Effect of Enzyme Dosage
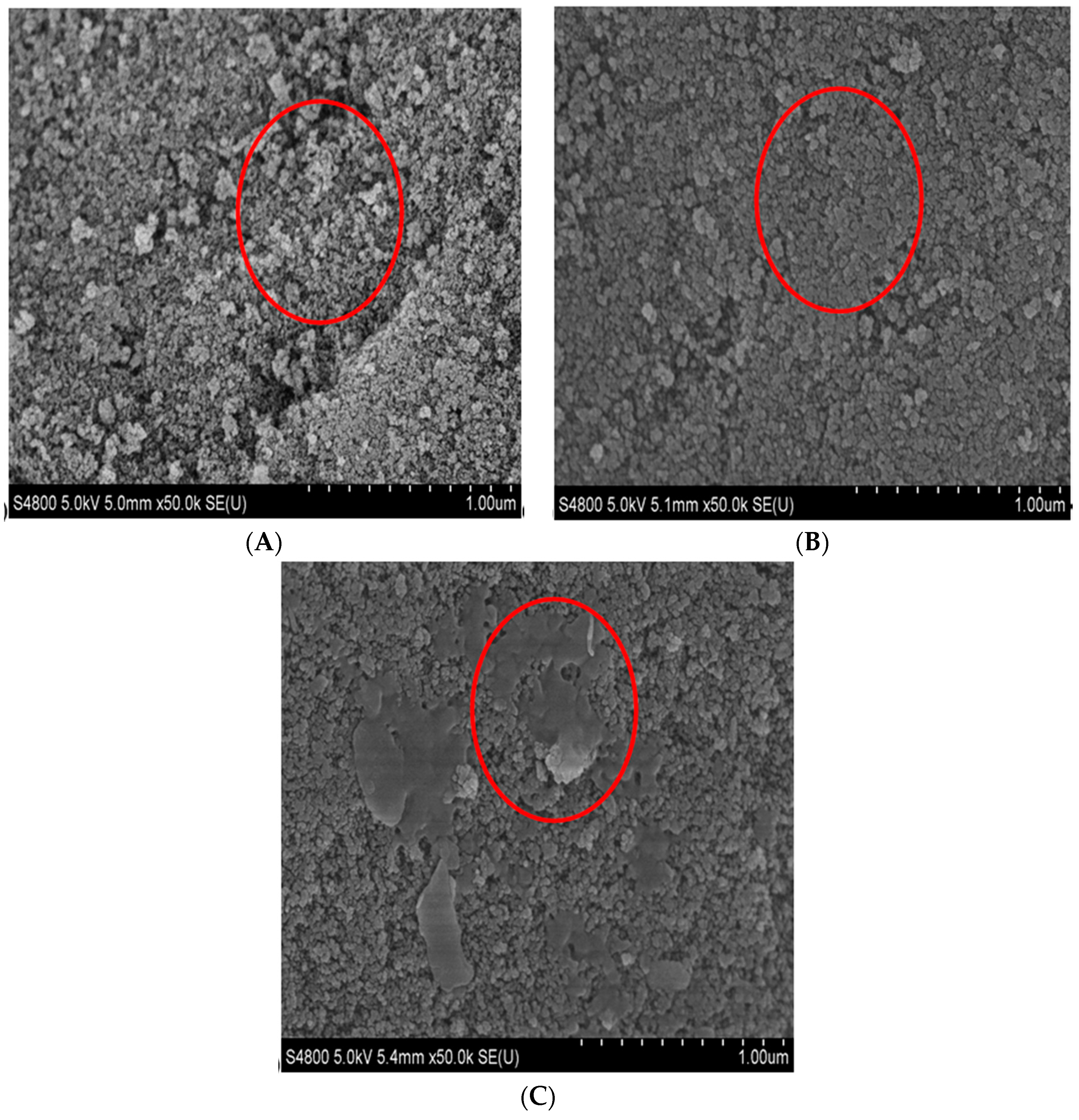
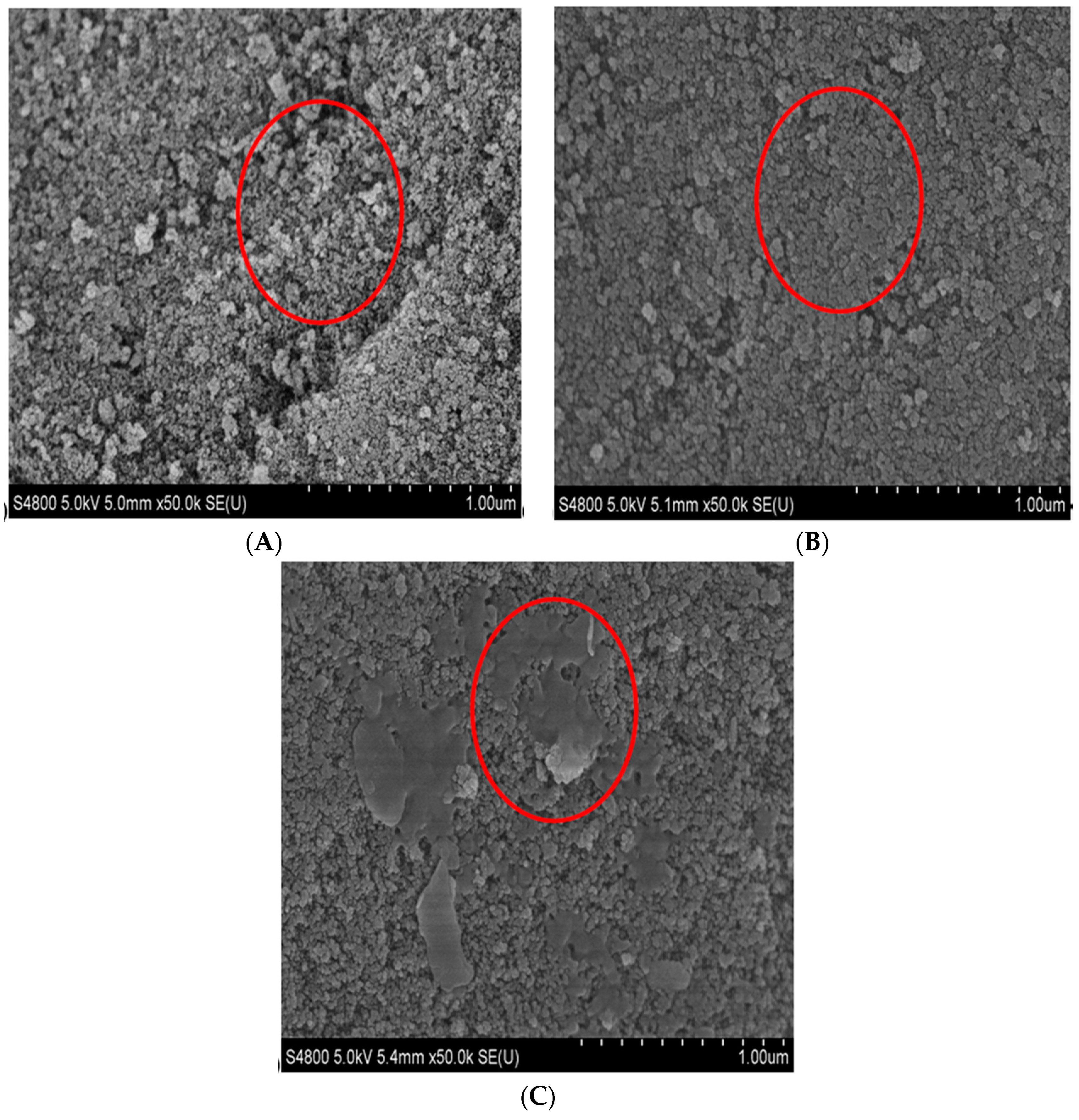
2.2. Characterization
2.3. Properties of Immobilized Cyclooxyenase-2
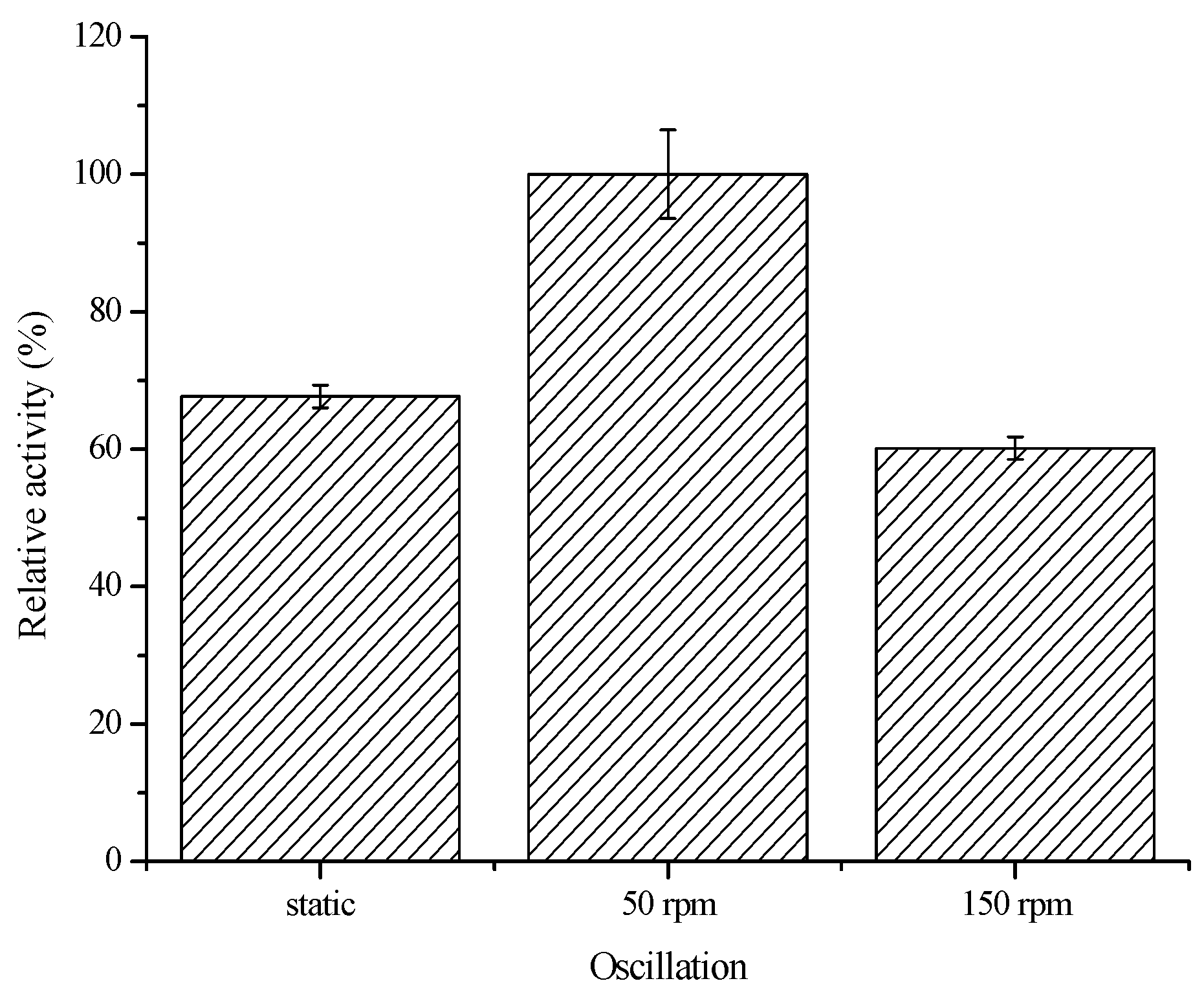
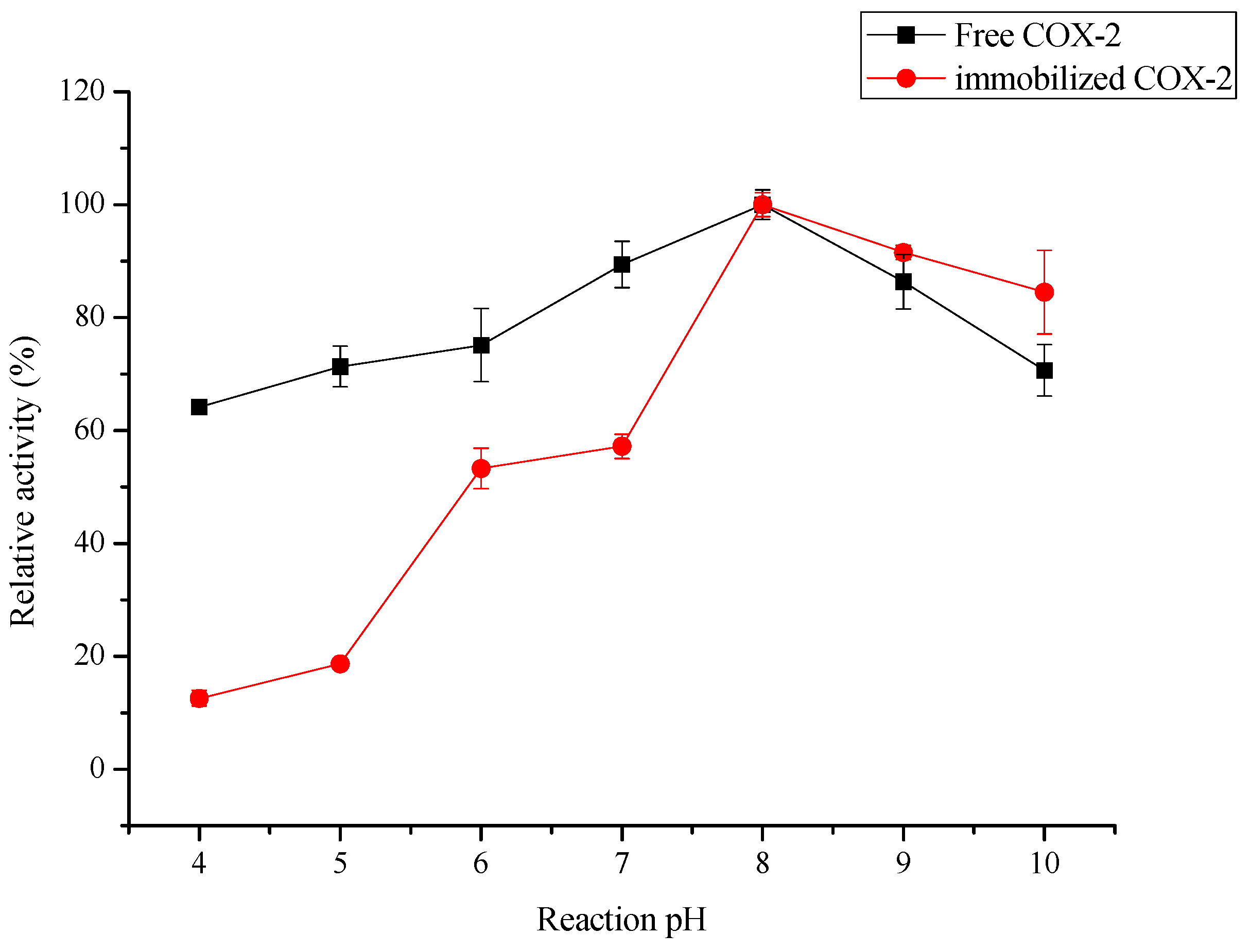
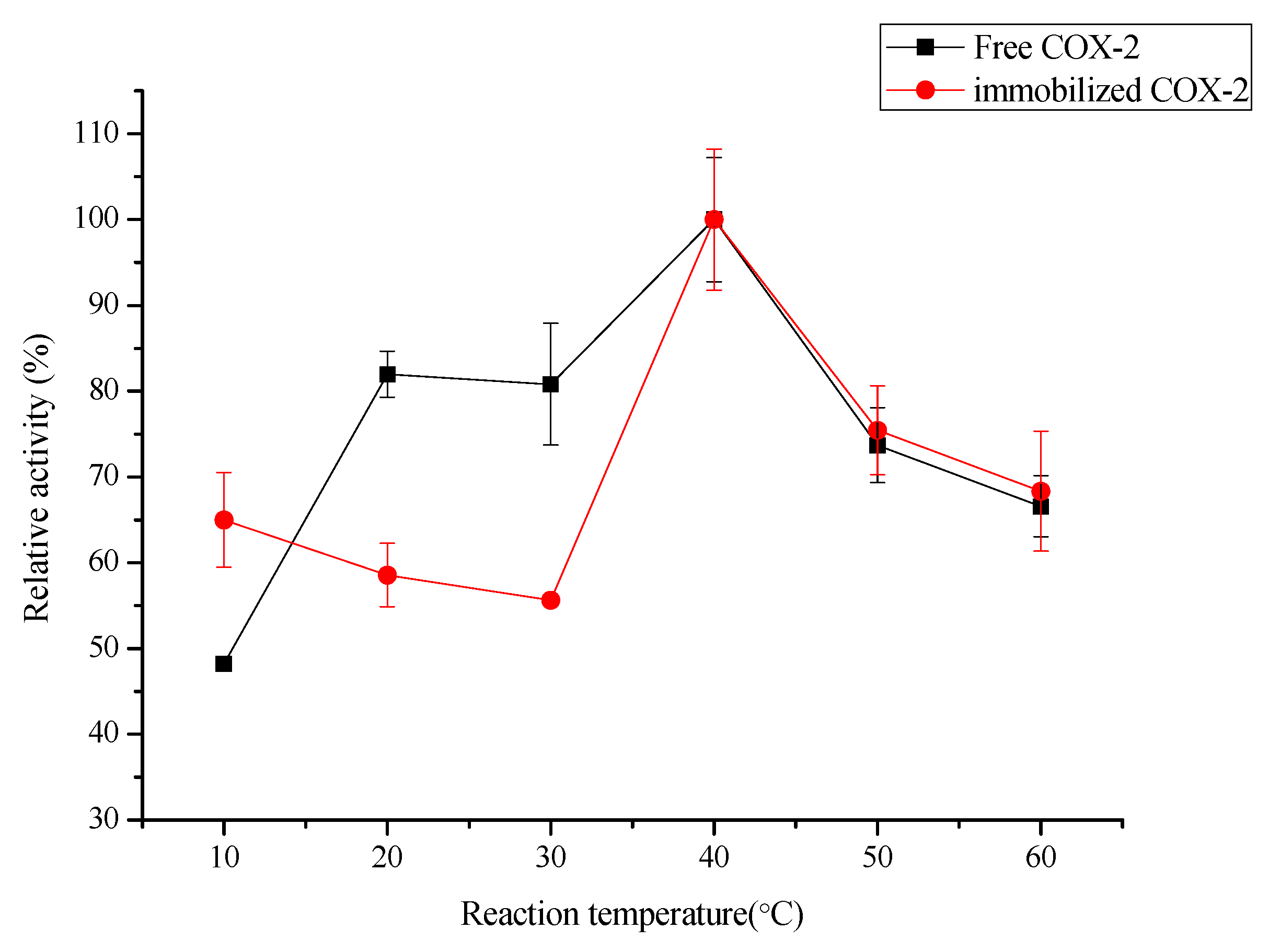
2.3.1. Effect of Oscillation, pH and Temperature on the Biocatalytic Activity of Immobilized COX-2


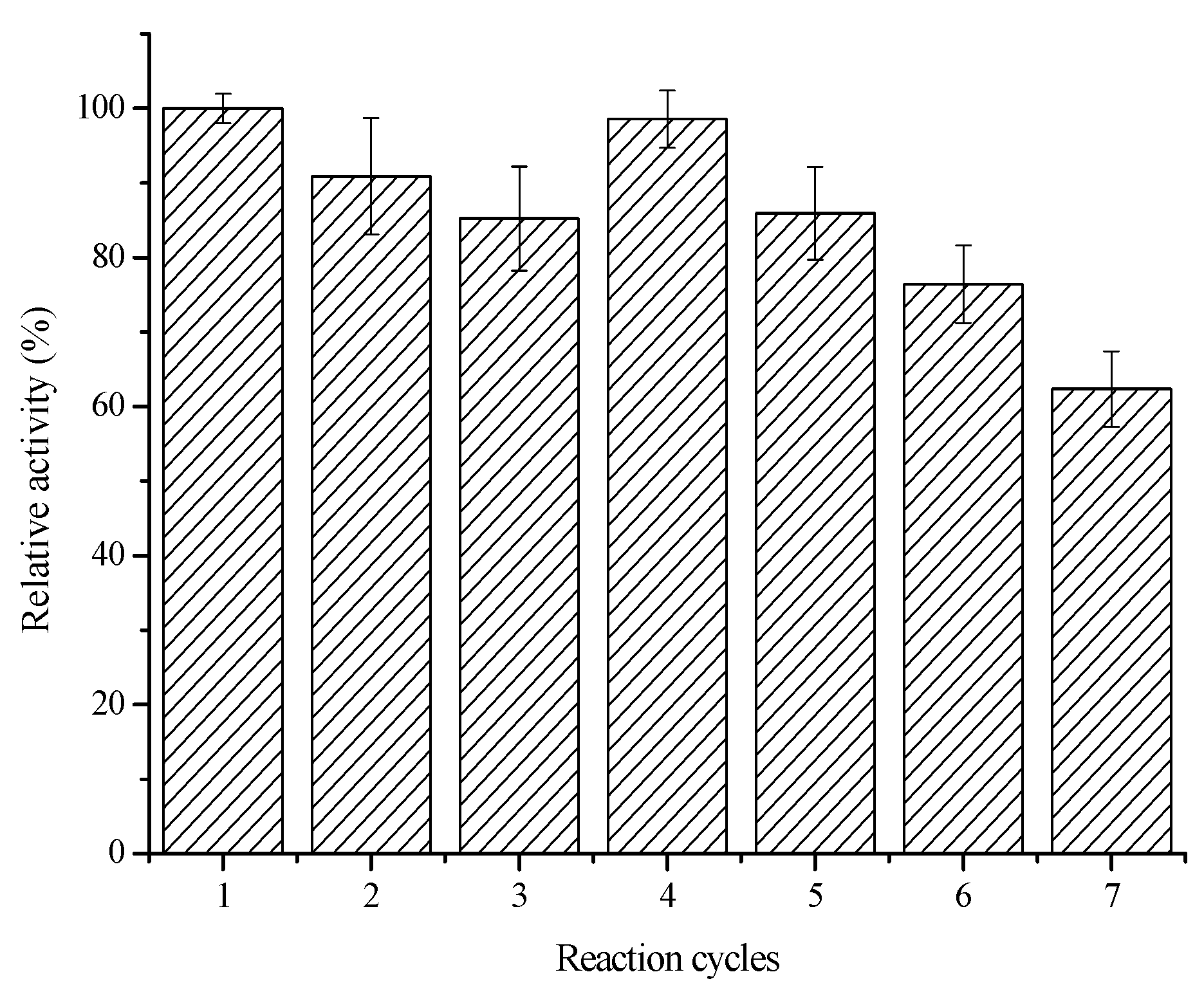
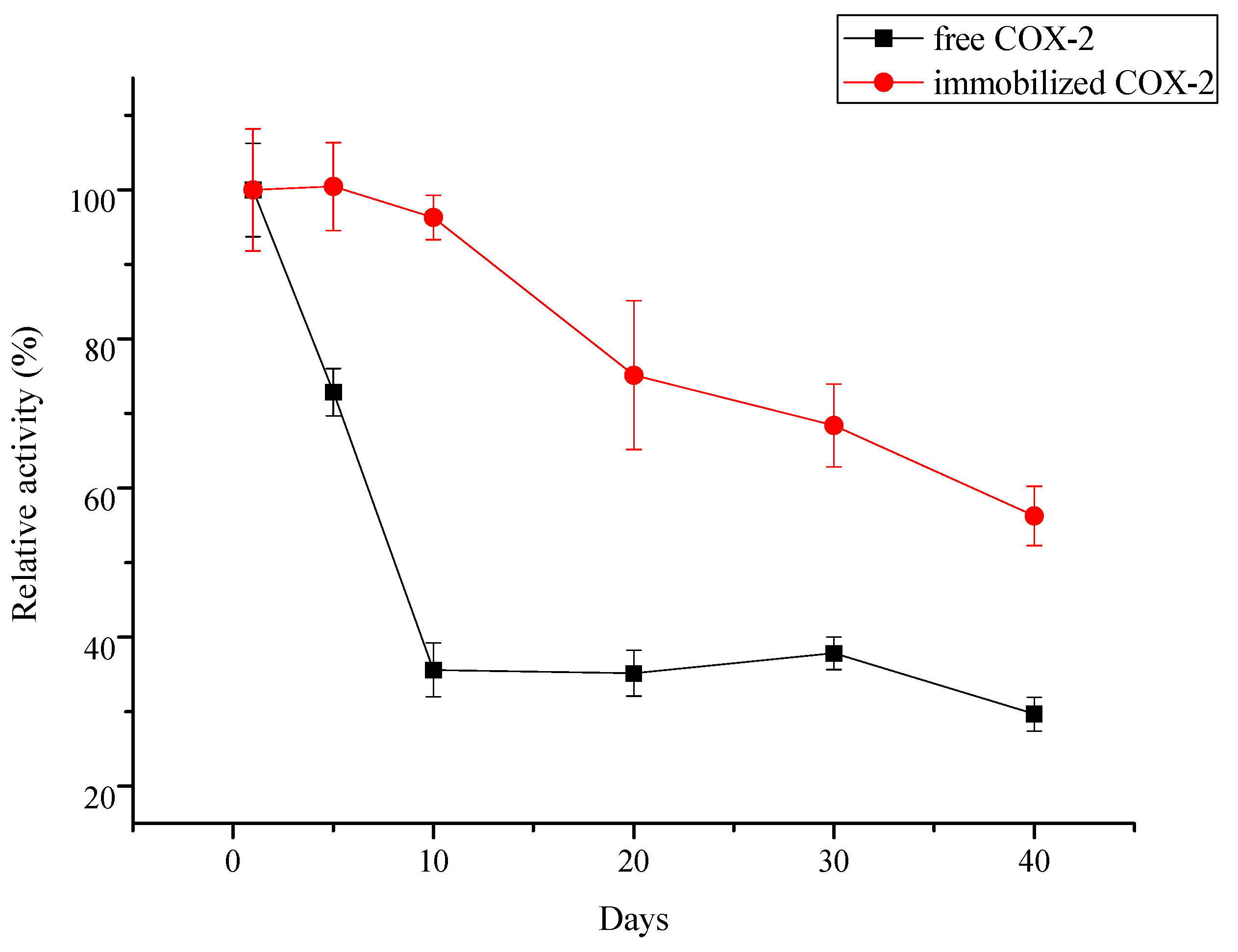
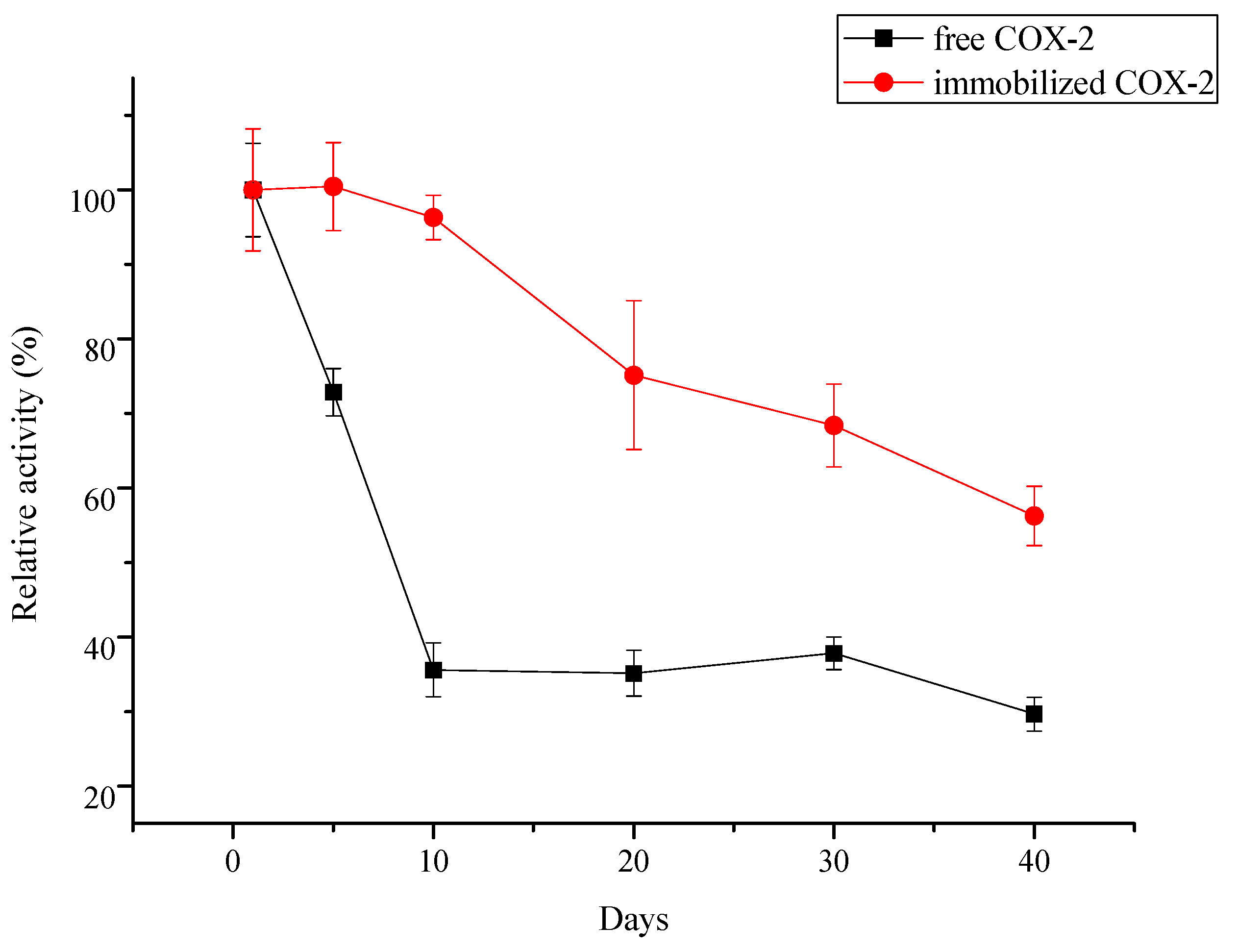
2.3.2. Reusability and Storage Stability

3. Experimental Section
3.1. Materials and Reagents
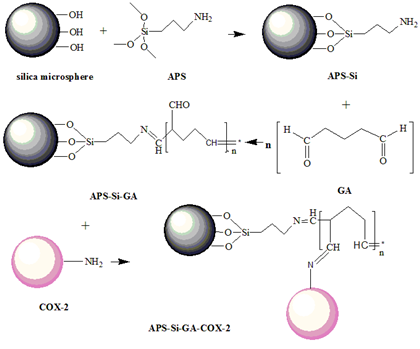
3.2. Preparation of NH2-Modified Silica Microspheres
3.3. Immobilization of Cyclooxyenase-2
3.4. Characterization
3.5. Determination of Immobilized Cyclooxyenase-2 Activity
3.6. Reusability of Immobilized Enzyme
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nikolic, D.; Habibi-Goudarzi, S.; Corley, D.G.; Gafner, S.; Pezzuto, J.M.; van Breemen, R.B. Evaluation of cyclooxygenase-2 inhibitors using pulsed ultrafiltration mass spectrometry. Anal. Chem. 2000, 72, 3853–3859. [Google Scholar] [CrossRef] [PubMed]
- Seibert, K.; Zhang, Y.; Leahy, K.; Hauser, S.; Masferrer, J.; Perkins, W.; Lee, L.; Isakson, P. Pharmacological and biochemical demonstration of the role of cyclooxygenase-2 in inflammation and pain. Proc. Natl. Acad. Sci. USA 1994, 91, 12013–12017. [Google Scholar] [CrossRef] [PubMed]
- Gately, S.; Li, W.W. Multiple roles of COX-2 in tumor angiogenesis: A target for antigiogenic therapy. Semin. Oncol. 2004, 31, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Samad, T.A.; Moore, K.A.; Sapirstein, A.; Billet, S.; Allchorne, A.; Poole, S.; Bonventre, J.V.; Woolf, C.J. Interleukin-1 beta-mediated induction of COX-2 in the CNS contributes to inflammatory pain hypersensitivity. Nature 2001, 410, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Catella-Lawson, F.; McAdam, B.; Morrison, B.W.; Kapoor, S.; Kujubu, D.; Antes, L.; Lasseter, K.C.; Quan, H.; Gertz, B.J.; FitzGerald, G.A. Effects of specific inhibition of cyclooxygenase-2 on sodium balance, hemodynamics, and vasoactive eicosanoid. J. Pharmacol. Exp. Ther. 1999, 289, 735–741. [Google Scholar] [PubMed]
- Dinchuk, J.E.; Car, B.D.; Focht, R.J.; Johnston, J.J.; Jaffee, B.D.; Covington, M.B.; Contel, N.R.; Eng, V.M.; Collins, R.J.; Czerniak, P.M.; et al. Renal abnormalities and an altered inflammatory response in mice lacking cyclooxygenase II. Nature 1995, 378, 406–409. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Shi, S.; Li, S.; Yang, T. Magnetic ligand fishing combination with high-performance liquid chromatography-diode array detector-mass spectrometry to screen and characterize cyclooxygenase-2 inhibitors from green tea. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 973C, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhuang, X.; Liu, S.; Pi, Z.; Song, F.; Lin, N.; Liu, Z. Ultrahigh-performance liquid chromatography/tandem mass spectrometry method for evaluating enzyme activity and screening inhibitors of cyclooxygenase-2. Rapid Commun. Mass Spectrom. 2014, 28, 1792–1800. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Yu, R.; Tao, Y.; Nikolic, D.; van Breemen, R.B. Measurement of cyclooxygenase inhibition using liquid chromatography-tandem mass spectrometry. J. Pharm. Biomed. Anal. 2011, 54, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Dannhardt, G.; Ulbrich, H. In-vitro test system for the evaluation of cyclooxygenase-1 (COX-1) and cyclooxygenase-2 (COX-2) inhibitors based on a single HPLC run with UV detection using bovine aortic coronary endothelial cells (BAECs). Inflamm. Res. 2001, 50, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Van Breemen, R.B.; Tao, Y.; Li, W. Cyclooxygenase-2 inhibitors in ginger (Zingiberofficinale). Fitoterapia 2011, 82, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Liu, S.; Li, X.; Song, F.; Liu, Z.; Liu, S. Bioactivity fingerprint analysis of cyclooxygenase-2 ligands from radix Aconiti by ultrafiltration-UPLC-MSn. Anal. Bioanal. Chem. 2013, 405, 7437–7445. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Frenz, C.M.; Chen, M.; Wang, Y.; Li, F.; Luo, C.; Liang, N.; Yang, H.; Bohlin, L.; Wang, C. Primary virtual and in vitro bioassay screening of natural Inhibitors from flavonoids against COX-2. Chin. J. Nat. Med. 2011, 9, 156–160. [Google Scholar]
- Chen, Y.C. De novo design of novel selective COX-2 inhibitors: From virtual screening to pharmacophore analysis. J. Taiwan Inst. Chem. Eng. 2009, 40, 55–69. [Google Scholar] [CrossRef]
- Laufer, S.; Zechmeister, P.; Klein, T. Development of an in vitro test system for the evaluation of cyclooxygenase-2 inhibitors. Inflamm. Res. 1999, 48, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, S.A.; Khan, J.A.; Al-Bar, O.A.; El-Shishtawy, R.M. Immobilization of Trichoderma harzianum α-Amylase on Treated Wool: Optimization and Characterization. Molecules 2014, 19, 8027–8038. [Google Scholar] [CrossRef] [PubMed]
- Iyer, P.V.; Ananthanarayan, L. Enzyme stability and stabilization-Aqueous and non-aqueous environment. Process Biochem. 2008, 43, 1019–1032. [Google Scholar] [CrossRef]
- Chang, Q.; Tang, H. Immobilization of Horseradish Peroxidase on NH2-Modified Magnetic Fe3O4/SiO2 Particles and Its Application in Removal of 2,4-Dichlorophenol. Molecules 2014, 19, 15768–15782. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lafuente, R. Stabilization of multimeric enzymes: Strategies to prevent subunit dissociation. Enzyme Microb. Technol. 2009, 45, 405–418. [Google Scholar] [CrossRef]
- Mateo, C.; Palomo, J.M.; Fernandez-Lorente, G.; Guisan, J.M.; Fernandez-Lafuente, R. Improvement of enzyme activity, stability and selectivity via immobilization techniques. Enzyme Microb. Technol. 2007, 40, 1451–1463. [Google Scholar] [CrossRef]
- Fernández-Lafuente, R. Editorial: Special issue-enzyme immobilization. Molecules 2014, 19, 20671–20674. [Google Scholar] [CrossRef] [PubMed]
- Kondo, A.; Teshima, T. Preparation of immobilized enzyme with high activity using affinitytag based on proteins A and G. Biotechnol. Bioeng. 1995, 46, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Fernández-Lorente, G.; Pessela, B.C.; Vian, A.; Carrascosa, A.V.; Garcia, J.L.; Fernández-Lafuente, R.; Guisan, J.M. Affinity chromatography of polyhistidine tagged enzymes—New dextran-coated immobilized metal ion affinity chromatography matrices for prevention of undesired multipoint adsorptions. J. Chromatogr. A 2001, 915, 97–106. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Zhang, Y. Characterization of the diatomite binding domain in the ribosomal protein L2 from E. coli and functions as an affinity tag. Appl. Microbiol. Biotechnol. 2013, 97, 2541–2549. [Google Scholar] [CrossRef] [PubMed]
- López-Gallego, F.; Betancor, L.; Mateo, C.; Hidalgo, A.; Alonso-Morales, N.; DellamoraOrtiz, G.; Guisán, J.M.; Fernández-Lafuente, R. Enzyme stabilization by glutaraldehyde crosslinking ofadsorbed proteins on aminated supports. J. Biotechnol. 2005, 119, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Betancor, L.; López-Gallego, F.; Hidalg, A.; Alonso-Morales, N.; Mateo, G.D.; Fernández-Lafuente, R.; Guisán, J.M. Different mechanisms of protein immobilization on glutaraldehyde activated supports: Effect of support activation and immobilization conditions. Enzyme Microb. Technol. 2006, 39, 877–882. [Google Scholar] [CrossRef]
- Monsan, P. Optimization of glutaraldehyde activation of a support for enzyme immobilization. J. Mol. Catal. 1978, 3, 371–384. [Google Scholar] [CrossRef]
- Barbosa, O.; Torres, R.; Ortiz, C.; Fernandez-Lafuente, R. Versatility of glutaraldehyde to immobilize lipases: Effect of the immobilization protocol on the properties of lipase B from Candida antarctica. Process Biochem. 2012, 47, 1220–1227. [Google Scholar] [CrossRef]
- Ma, D.; Guan, J.; Normandin, F.; Dénommée, S.; Enright, G.; Veres, T.; Simard, B. Multifunctional nano-architecture for biomedical applications. Chem. Mater. 2006, 18, 1920–1927. [Google Scholar] [CrossRef]
- Solís, S.; Paniagua, J.; Martínez, J.C.; Asomoza, M. Immobilization of papain on mesoporous silica: pH effect. J. Sol-Gel Sci. Technol. 2006, 37, 125–127. [Google Scholar] [CrossRef]
- Aissaoui, N.; Bergaoui, L.; Boujday, S.; Lambert, J.-F.; Méthivier, C.; Landoulsi, J. Enzyme immobilization on silane-modified surface through short linkers: Fate of interfacial phases and impact on catalytic activity. Longmuir 2014, 30, 4066–4077. [Google Scholar] [CrossRef] [PubMed]
- Monsan, P.; Puzo, G.; Mazarguil, H. Étude du mécanisme d’ établissement des liaisons glutaraldéhyde-protéines. Biochimie 1975, 57, 1281–1292. [Google Scholar] [CrossRef]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Glutaraldehyde in bio-catalysts design: A useful crosslinker and a versatile tool in enzyme immobilization. RSC Adv. 2014, 4, 1583–1600. [Google Scholar] [CrossRef]
- Migneault, I.; Dartiguenave, C.; Bertrand, M.J.; Waldron, K.C. Glutaraldehyde: Behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. Biotechniques 2004, 37, 790–796, 798–802. [Google Scholar] [PubMed]
- Okuda, K.; Urabe, I.; Yamada, Y.; Okada, H. Reaction of glutaraldehyde with amino and thiol compounds. J. Ferment. Bioeng. 1991, 71, 100–105. [Google Scholar] [CrossRef]
- Nimni, M.E.; Cheung, D.; Strates, B.; Kodama, M.; Sheikh, K. Chemically modified collagen: A natural biomaterial for tissue replacement. J. Biomed. Mater. Res. 1987, 21, 741–771. [Google Scholar] [CrossRef] [PubMed]
- Tümtürk, H.; Sahin, F.; Demirel, G. A new method for immobilization of acetylcholinesterase. Bioprocess Biosyst. Eng. 2007, 30, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Poltorak, O.M.; Chukhrai, E.S.; Kozlenkov, A.A.; Chaplin, M.F.; Trevan, M.D. The putative common mechanism for inactivation of alkaline phosphatase isoenzymes. J. Mol. Catal. B Enzym. 1999, 7, 157–163. [Google Scholar] [CrossRef]
- Phlipenko, O.S.; Atyaksheva, L.F.; Pltorak, O.M.; Chukhrai, E.S. Dissociation and catalytic activity of oligomer forms of β-galactosidases. Russ. J. Phys. Chem. A 2007, 81, 990–994. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, A.; Torres, R.; Fernández-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Cui, J.; Li, G.; Zhang, J.; Li, D.; Huang, F.; Wei, Q. Laccase Immobilized on a PAN/Adsorbents Composite Nanofibrous Membrane for Catechol Treatment by a Biocatalysis/Adsorption Process. Molecules 2014, 19, 3376–3388. [Google Scholar] [CrossRef] [PubMed]
- Jaenicke, R.; Bohm, G. The stability of proteins in extreme environments. Curr. Opin. Struct. Biol. 1998, 8, 738–748. [Google Scholar] [CrossRef]
- Abaházi, E.; Boros, Z.; Poppe, L. Additives enhancing the catalytic properties of lipase from burkholderiacepacia immobilized on mixed-function-grafted mesoporous silica gel. Molecules 2014, 19, 9818–9837. [Google Scholar] [CrossRef] [PubMed]
- Chapman, E.G.; Piontkivska, H.; Walker, J.M.; Stewart, D.T.; Curole, J.P.; Hoeh, W.R. Extreme primary and secondary protein structure variability in the chimeric male-transmitted cytochrome c oxidase subunit II protein in freshwater mussels: Evidence for an elevated amino acid substitution rate in the face of domain-specific purifying selection. BMC Evol. Biol. 2008, 8, 165–180. [Google Scholar] [PubMed]
- Chen, J.; Shi, Q.; Chen, C.; Li, X.; Cao, W.; Zheng, L.; Wang, X. Preparation and characteristic research of immobilized acetylcholinesterase. Acta Chim. Sin. 2012, 70, 624–628. [Google Scholar] [CrossRef]
- Sample Availability: Samples of soluble and immobilized cyclooxygenase-2 are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Q.; Chen, J.; Wang, Y.; Li, Z.; Li, X.; Sun, C.; Zheng, L. Immobilization of Cyclooxygenase-2 on Silica Gel Microspheres: Optimization and Characterization. Molecules 2015, 20, 19971-19983. https://doi.org/10.3390/molecules201119670
Shi Q, Chen J, Wang Y, Li Z, Li X, Sun C, Zheng L. Immobilization of Cyclooxygenase-2 on Silica Gel Microspheres: Optimization and Characterization. Molecules. 2015; 20(11):19971-19983. https://doi.org/10.3390/molecules201119670
Chicago/Turabian StyleShi, Qian, Junhui Chen, Yanlong Wang, Zhaoyong Li, Xianguo Li, Chengjun Sun, and Li Zheng. 2015. "Immobilization of Cyclooxygenase-2 on Silica Gel Microspheres: Optimization and Characterization" Molecules 20, no. 11: 19971-19983. https://doi.org/10.3390/molecules201119670
APA StyleShi, Q., Chen, J., Wang, Y., Li, Z., Li, X., Sun, C., & Zheng, L. (2015). Immobilization of Cyclooxygenase-2 on Silica Gel Microspheres: Optimization and Characterization. Molecules, 20(11), 19971-19983. https://doi.org/10.3390/molecules201119670