
Water-Soluble Lignins from Different Bioenergy Crops Stimulate the Early Development of Maize (Zea mays, L.)


Abstract
:
1. Introduction
2. Results and Discussion
2.1. Extraction Yields
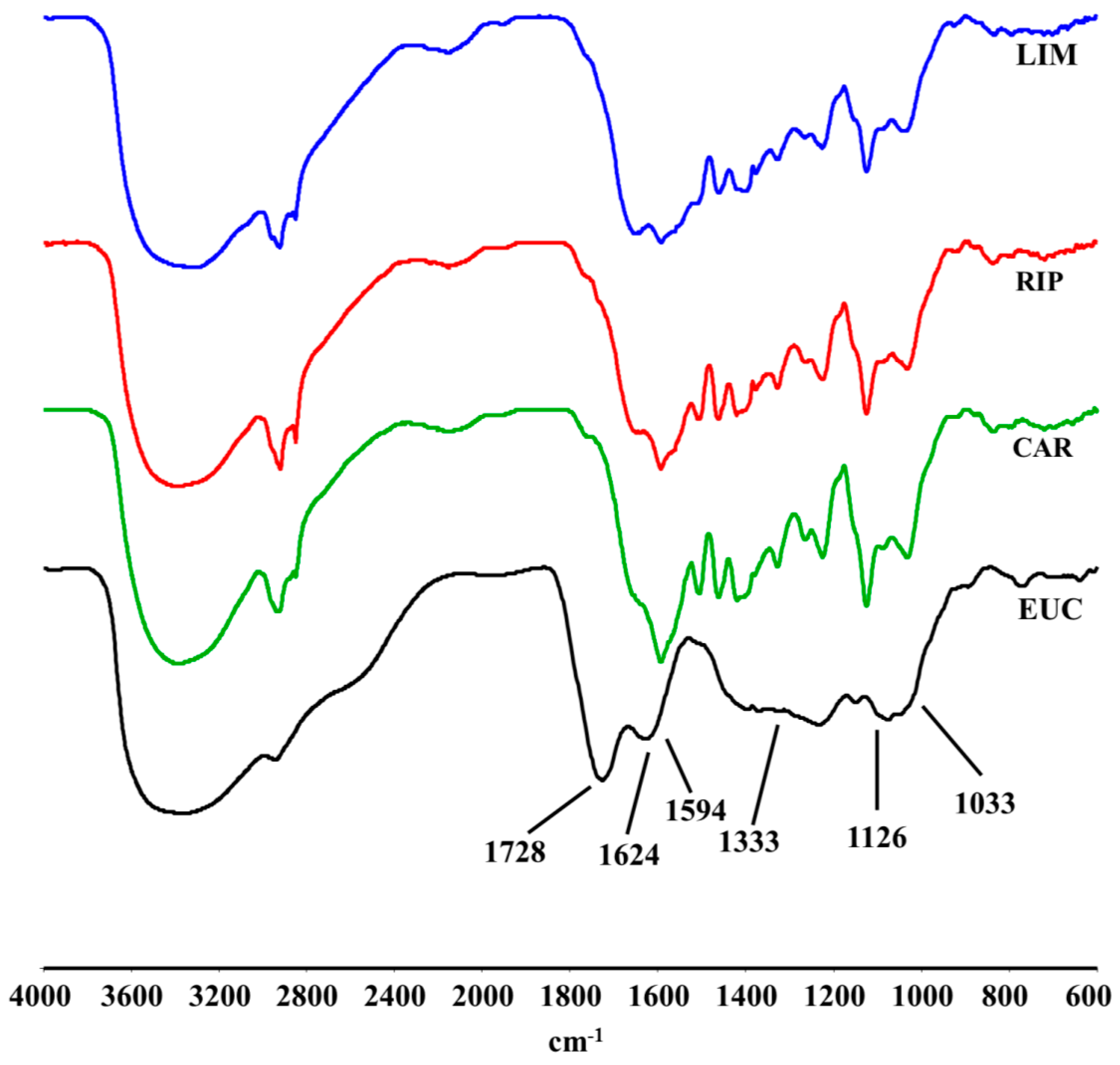
2.2. DRIFT-IR Spectra
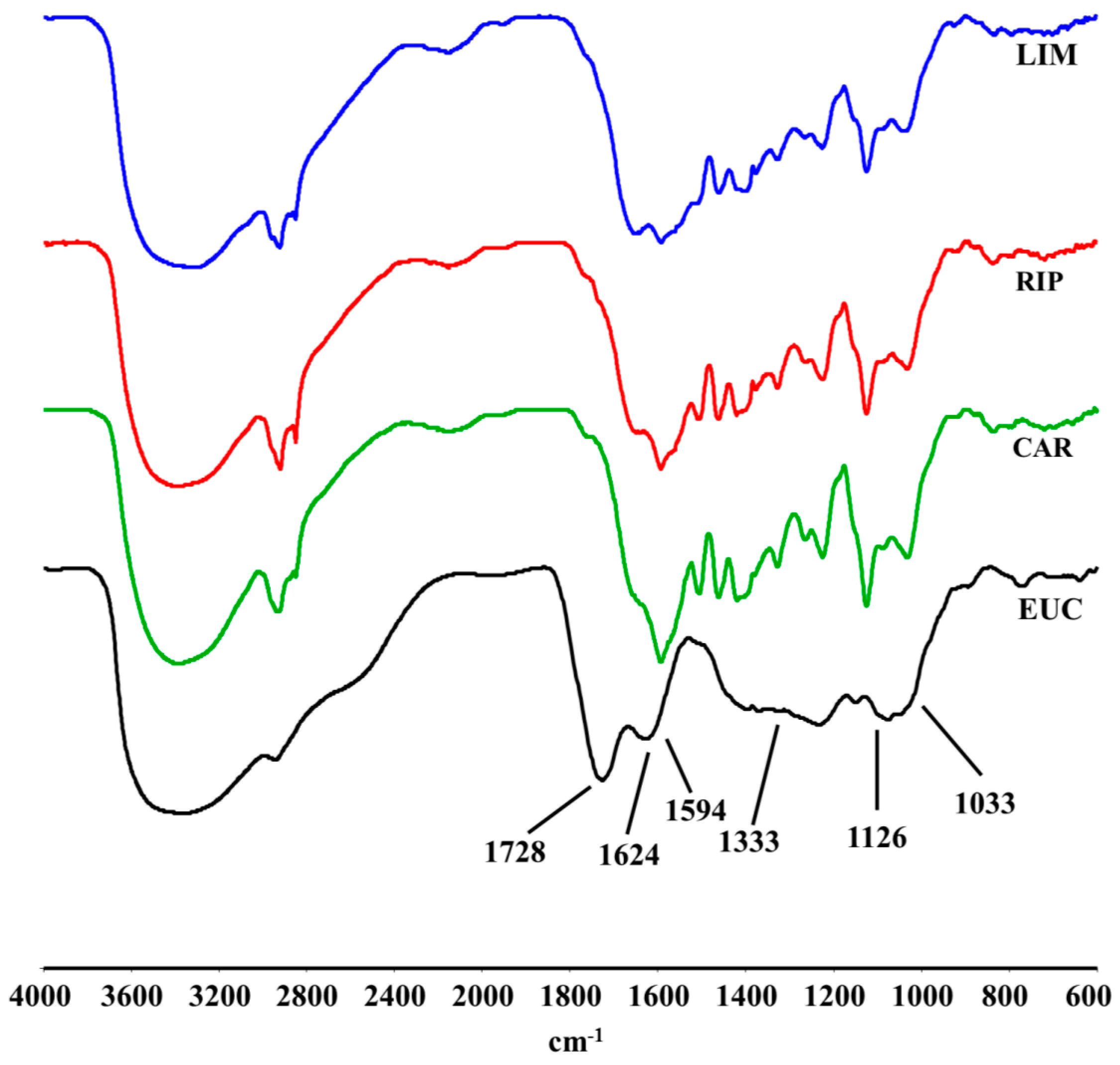
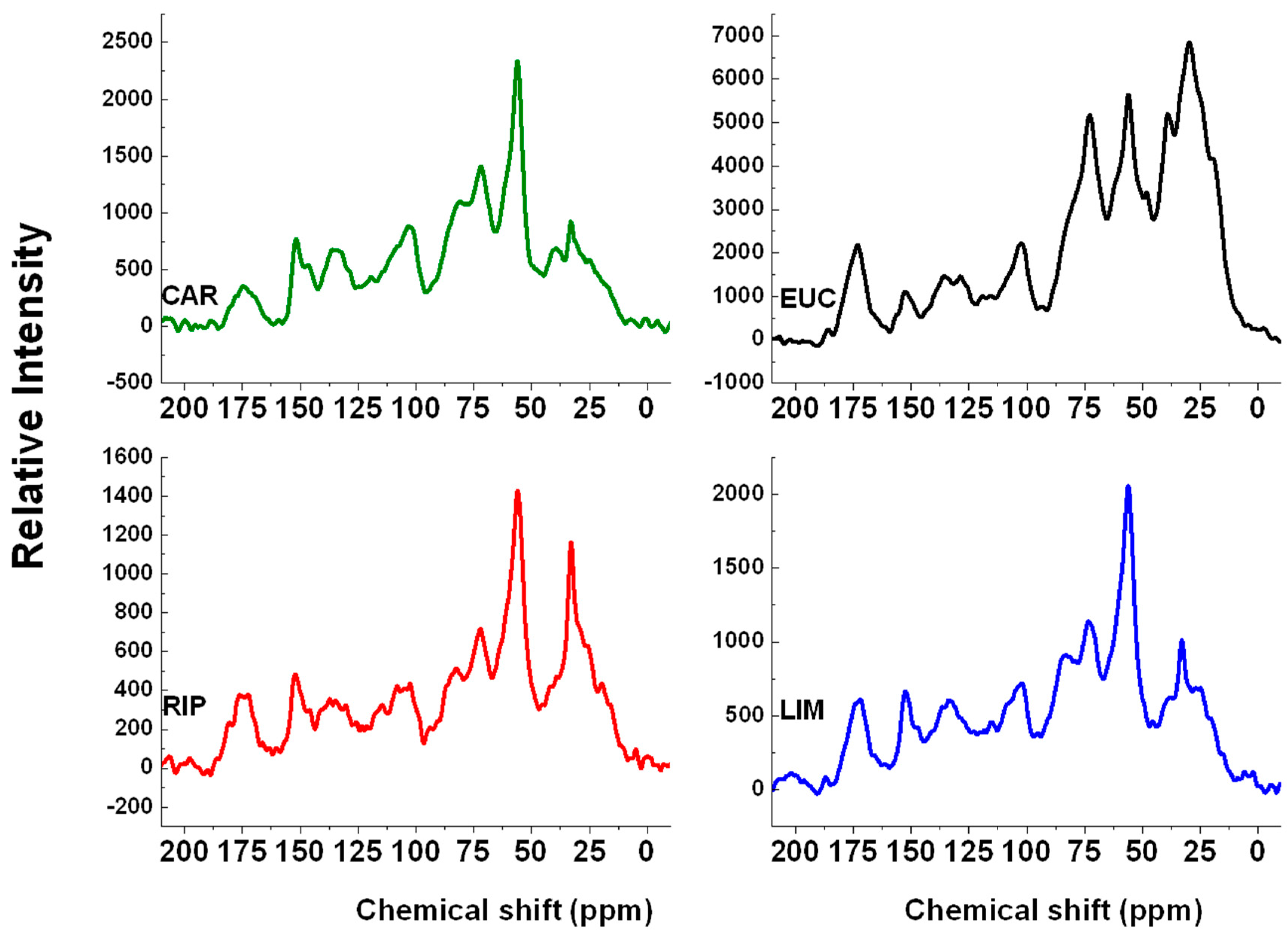
2.3. 13C-CPMAS-NMR Spectroscopy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lignin Extracts | Chemical Shift Regions | Indexes | ||||||
|---|---|---|---|---|---|---|---|---|
| 187–162 (COOH) | 162–112 (aromatic-C) | 112–93 (anomeric-C) | 93–46 (C-O, C-N) | 46–0 (Alkyl-C) | HB | HI | HB/HI | |
| CAR | 4 | 20 | 11 | 48 | 17 | 37 | 63 | 0.59 |
| EUC | 6 | 11 | 7 | 38 | 38 | 49 | 51 | 0.96 |
| RIP | 7 | 19 | 9 | 40 | 25 | 44 | 56 | 0.79 |
| LIM | 7 | 20 | 10 | 44 | 19 | 39 | 61 | 0.64 |
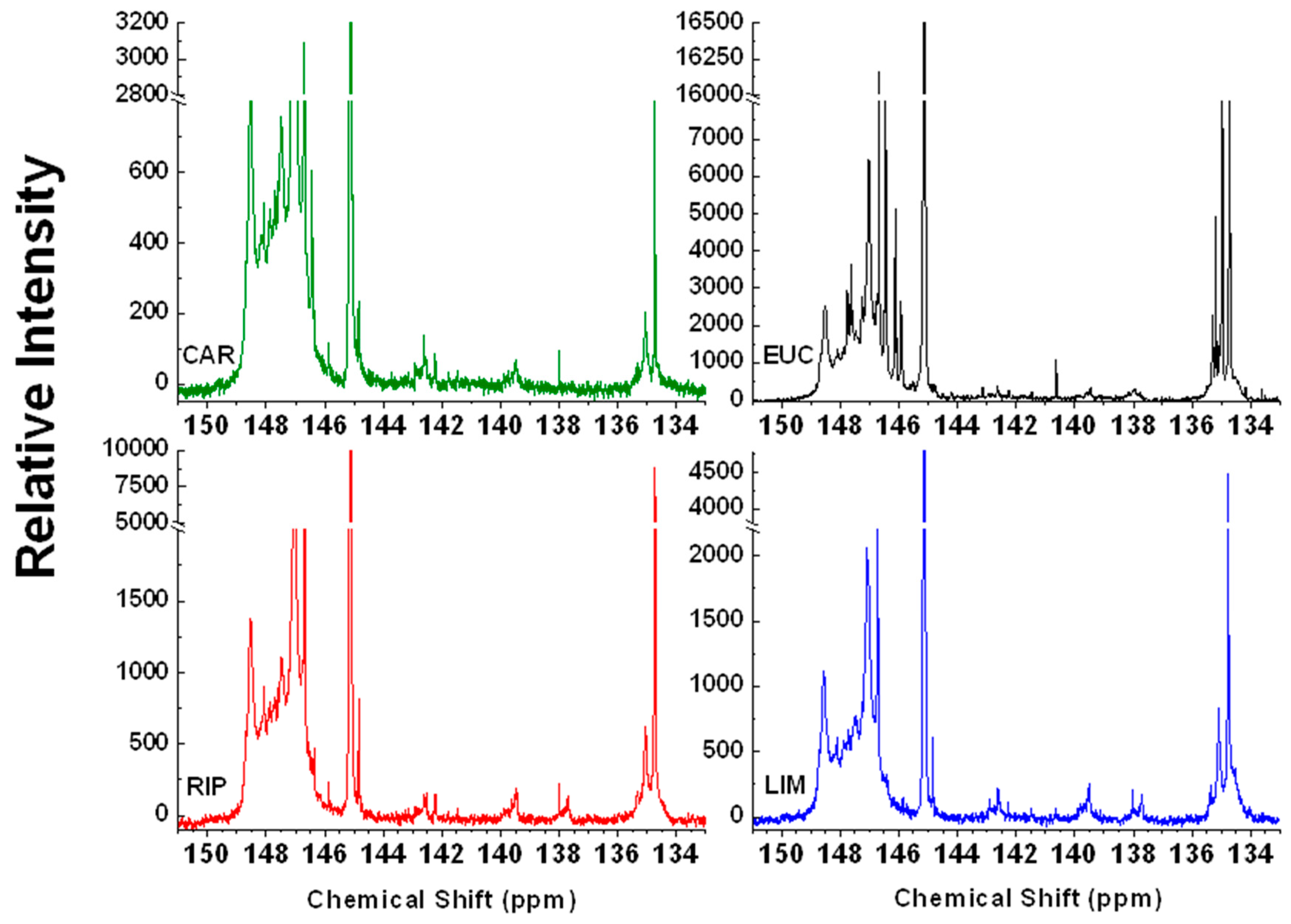
2.4. 31P-NMR Spectroscopy
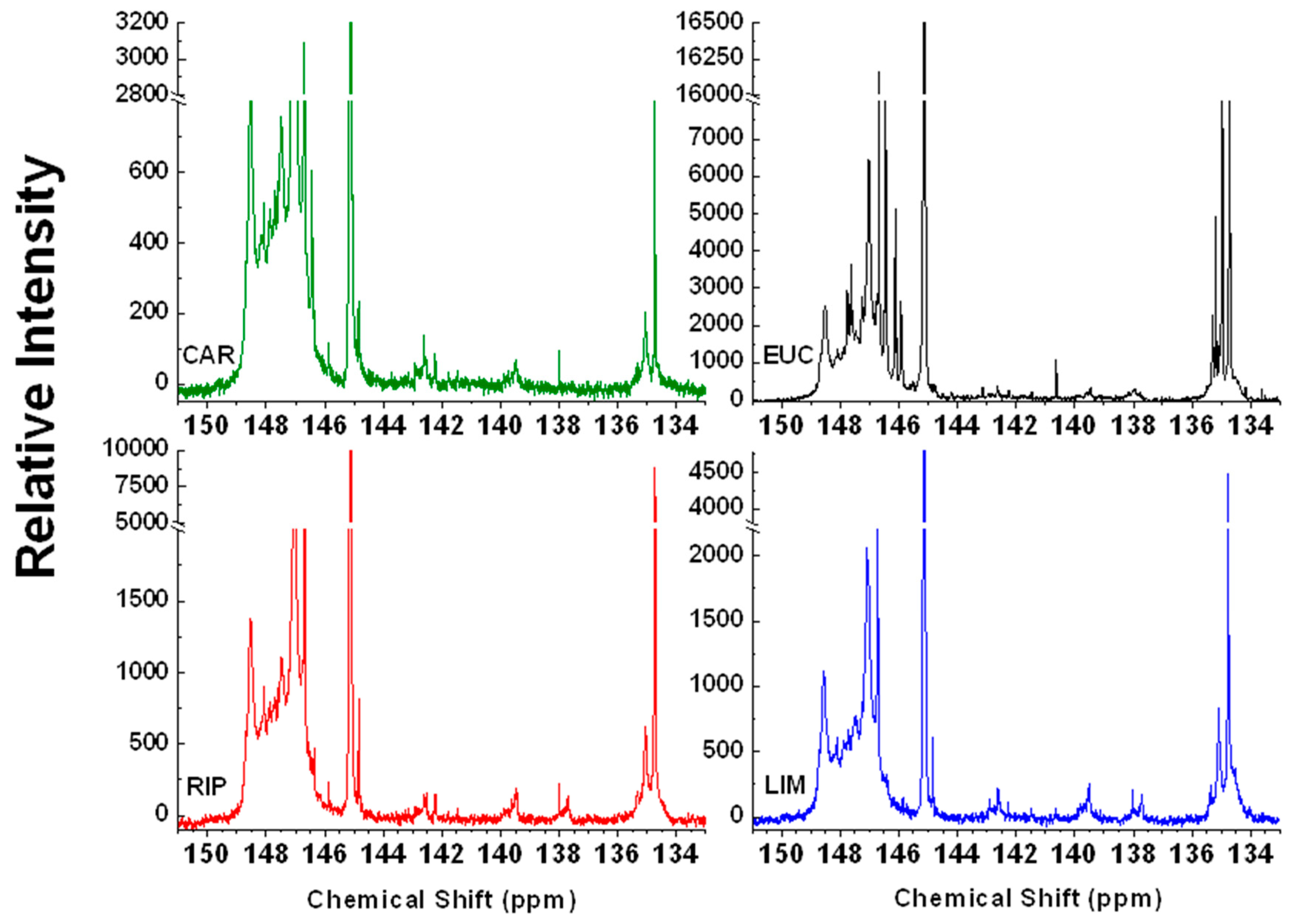
| Chemical Shift (ppm) | Assignment | CAR | EUC | RIP | LIM |
|---|---|---|---|---|---|
| 135.6–133.7 | Carboxyl groups | 0.42 | 1.41 | 1.02 | 1.23 |
| 138.6–136.9 | H-type molecules | ND | 0.07 | 0.04 | 0.07 |
| 140.2–138.4 | G-type molecules | 0.11 | 0.04 | 0.07 | 0.14 |
| 142.8–141.7 | Condensed (4-O-5′,5-5′) lignin dimers | 0.14 | 0.11 | 0.04 | 0.04 |
| 143.7–142.2 | S-type molecules | 0.14 | 0.04 | 0.07 | 0.11 |
| 143.7–136.9 | Total Phenolic Content | 0.39 | 0.25 | 0.21 | 0.35 |
| 150.8–146.3 | Aliphatic OH | 6.76 | 3.94 | 5.42 | 4.68 |
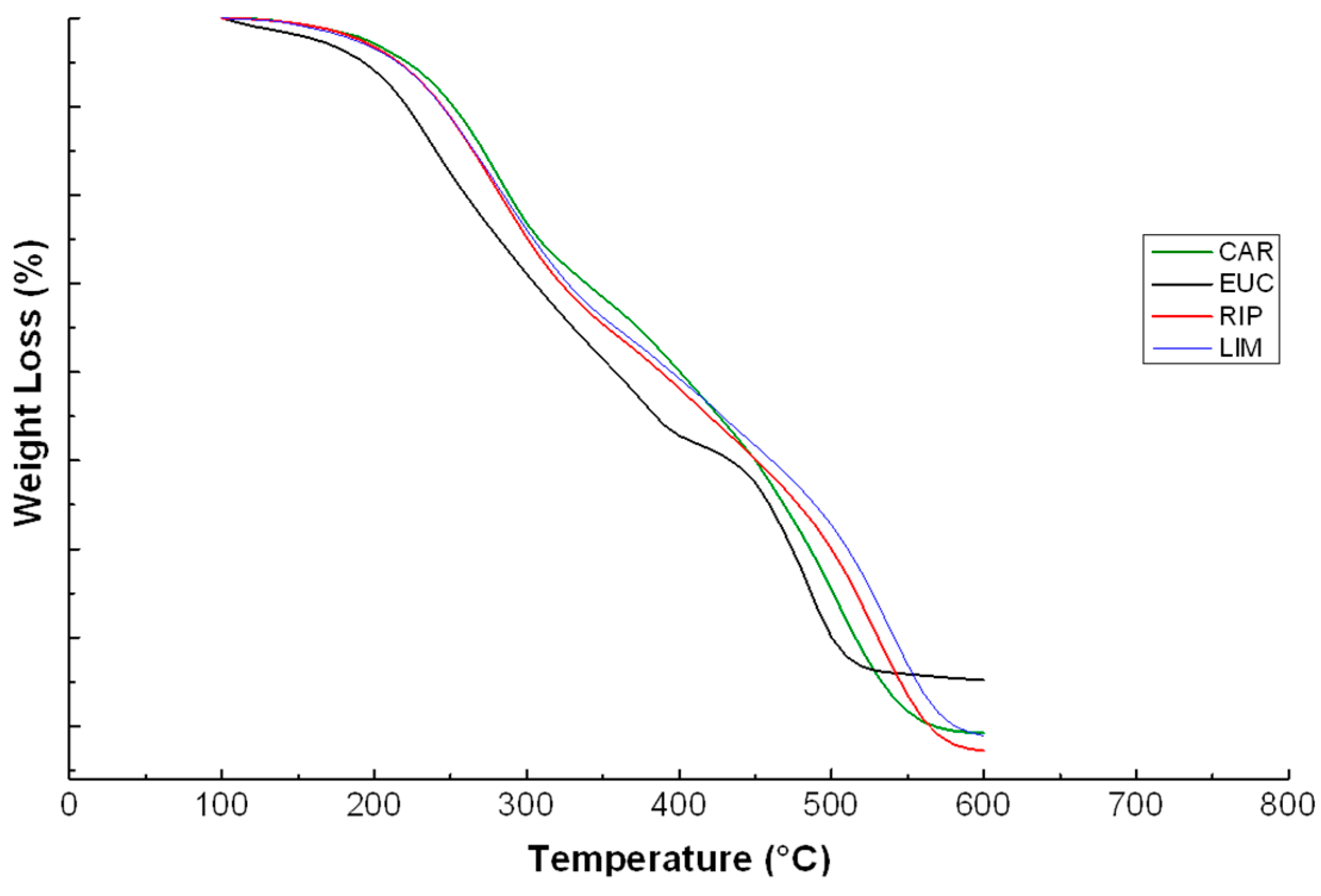
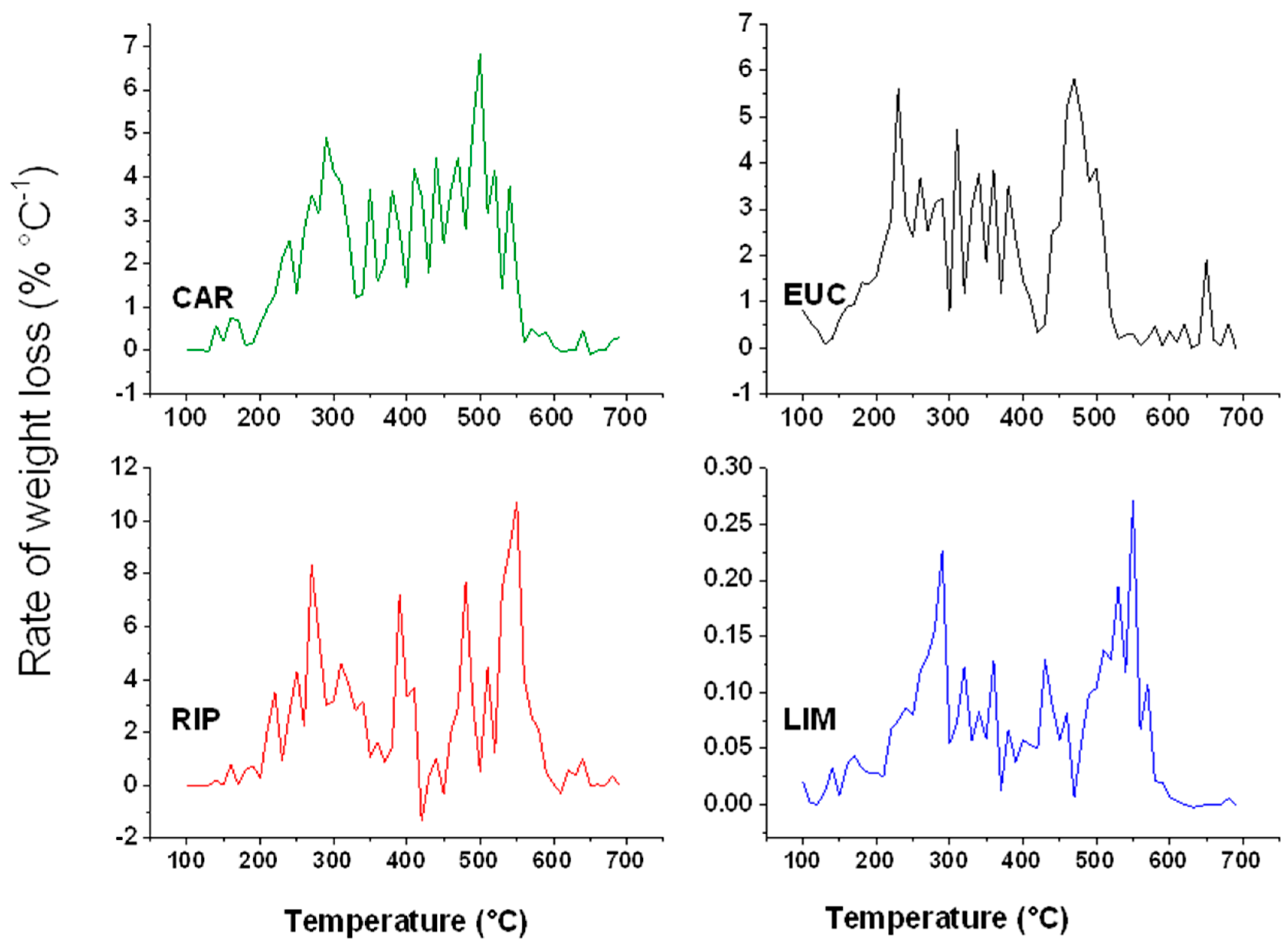
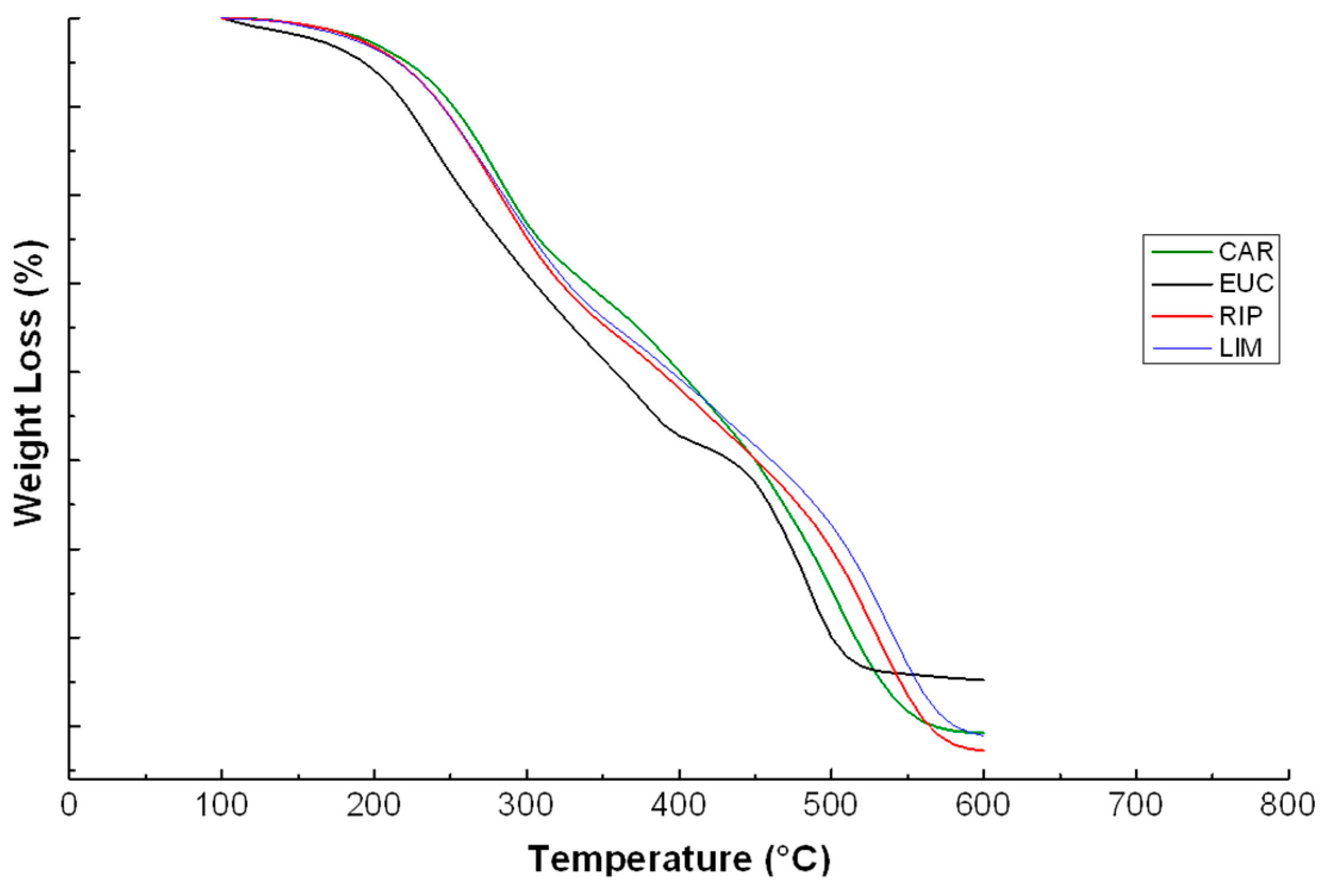
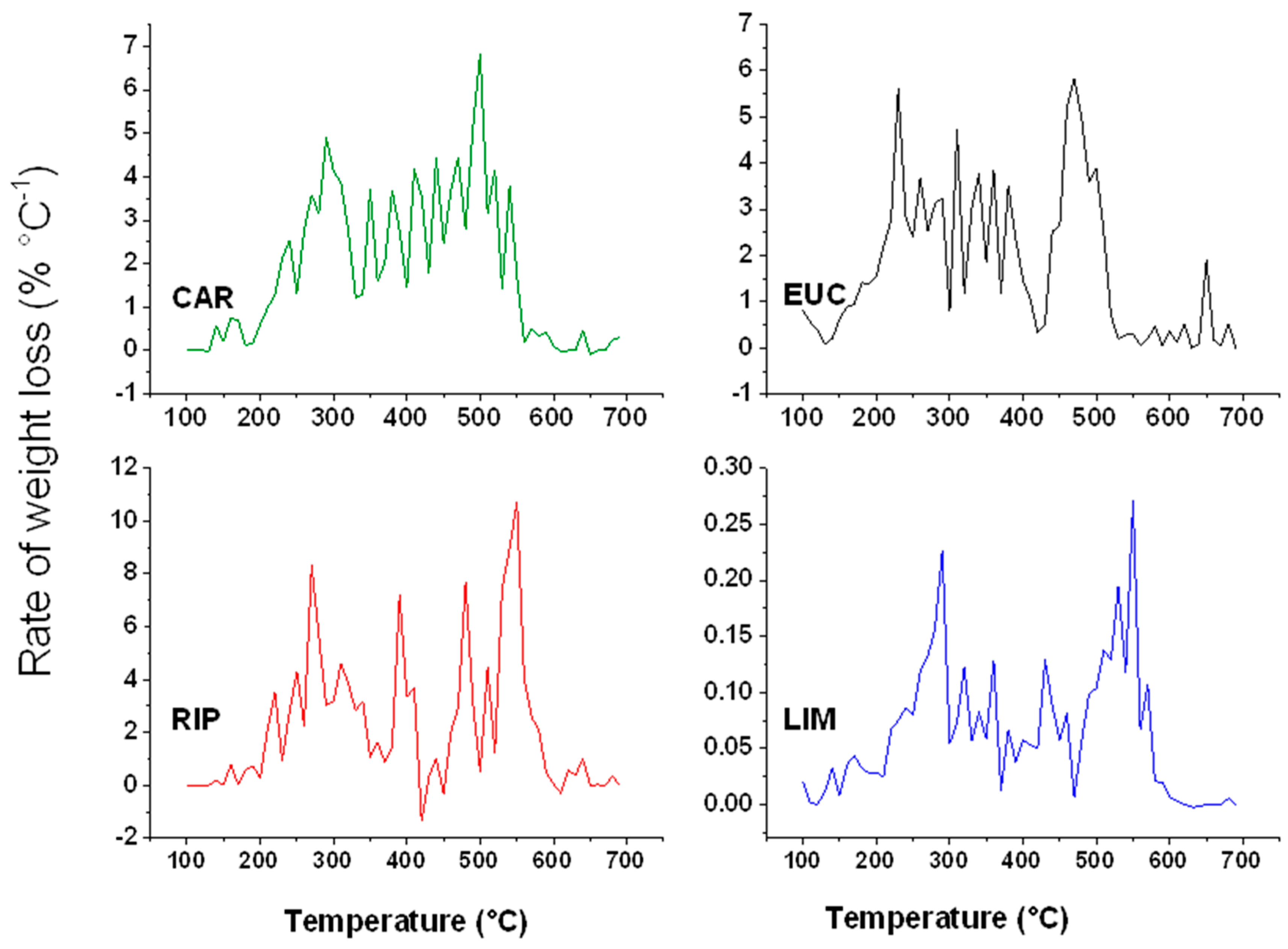
2.5. Thermal Analyses
2.6. Germination of Maize Seeds and Early Growth
| Lignin Concentration (ppm) | CAR | EUC | RIP | LIM |
|---|---|---|---|---|
| 0—Control | 6.5 (100) a | 10.8 (100) a | 5.5 (100) a | 6.4 (100) a |
| 0.1 | 5.3 (82) a | 10.8 (100) a | 6.7 (122) a,b | 6.4 (101) a |
| 1 | 6.3 (98) a | 10.6 (99) a | 5.9 (107) a,b | 7.4 (116) a |
| 10 | 7.0 (108) a | 9.5 (88) a | 6.4 (117) a,b | 6.3 (99) a |
| 100 | 4.9 (76) a | 10.5 (96) a | 6.8 (124) b | 7.5 (118) a |
| Lignin Concentration (ppm) | CAR | EUC | RIP | LIM |
|---|---|---|---|---|
| 0—Control | 5.9 (100) a | 19.8 (100) a | 8.2 (100) a | 8.7 (100) a |
| 0.1 | 6.5 (111) a,b | 19.3 (98) a | 11.2 (135) b | 8.9 (102) a |
| 1 | 7.0 (118) a,b | 18.6 (94) a | 7.9 (96) a | 10.2 (118) a |
| 10 | 8.3 (140) b | 16.0 (81) a | 9.9 (120) a,b | 9.3 (107) a |
| 100 | 5.7 (97) a | 20.8 (105) a | 10.2 (123) a,b | 8.9 (102) a |
| Lignin Concentration (ppm) | CAR | EUC | RIP | LIM |
|---|---|---|---|---|
| 0—Control | 3.6 (100) a | 8.5 (100) a | 5.1 (100) a | 5.2 (100) a |
| 0.1 | 5.5 (150) b,c | 8.9 (104) a | 5.4 (102) a,b | 5.7 (110) a,b |
| 1 | 5.3 (146) b,c | 8.8 (104) a | 5.5 (104) a,b | 5.5 (106) a,b |
| 10 | 6.3 (172) c | 8.1 (95) a | 5.6 (107) a,b | 5.4 (104) a,b |
| 100 | 4.6 (127) a,b | 8.4 (98) a | 5.9 (113) b | 6.0 (117) b |
3. Experimental
3.1. Biomasses
3.2. Alkaline Oxidative Hydrolysis
3.3. Diffuse Reflectance Infrared Fourier Transform Spectroscopy (DRIFT-IR)
3.4. Solid State-Nuclear Magnetic Resonance (NMR) Spectroscopy
3.5. Synthesis of 2-Chloro-4,4,5,5-Tetramethyldioxaphospholane
3.6. 31P-Nuclear Magnetic Resonance
3.7. Thermal Analysis
3.8. Germination and Seedling Emergence Assays
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cetinkaya, M.; Karaosmanoglu, F. Optimization of base-catalyzed transesterification reaction of used cooking oil. Energy Fuels 2004, 18, 1888–1895. [Google Scholar] [CrossRef]
- Nagendramma, P.; Kaul, S. Development of ecofriendly/biodegradable lubricants: An overview. Renew. Sustain. Energy Rev. 2012, 16, 764–774. [Google Scholar] [CrossRef]
- Ma, E.; Cervera, Q.; Sanchez, G.M.M. Integrated utilization of orange peel. Bioresour. Technol. 1993, 44, 61–63. [Google Scholar] [CrossRef]
- Li, Q.; Siles, J.A.; Thompson, I.A. Succinic acid production from orange peel and wheat straw by batch fermentations of Fibrobacter succinogenes S85. Appl. Microbiol. Biotechnol. 2010, 88, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.K.; Pfaltzfraff, L.A.; Herrero-Davila, L.; Mubofu, E.B.; Abderrahim, S.; Clark, J.H.; Koutinas, A.; Kopsahelis, N.; Stamatelatou, K.; Dickson, F.; et al. Food waste as a valuable resource for the production of chemicals, materials and fuels. Current situation and global perspective. Energy Environ. Sci. 2013, 6, 426–464. [Google Scholar] [CrossRef]
- Cherubini, F. The biorefinery concept: Using biomass instead of oil for producing energy and chemicals. Energy Convers. Manag. 2010, 51, 1412–1421. [Google Scholar] [CrossRef]
- Zakzeski, J.; Bruijnincx, P.C.A.; Jongerius, A.L.; Weckhuysen, B.M. The catalytic valorization of lignin for the production of renewable chemicals. Chem. Rev. 2010, 110, 3552–3599. [Google Scholar] [CrossRef] [PubMed]
- Holladay, J.E.; Bozell, J.J.; White, J.F.; Johnson, D. Top Value-Added Chemicals from Biomass. Volume II—Results of Screening for Potential Candidates from Biorefinery Lignin; PNNL-16983; Pacific Northwest National Laboratory: Richland, WA, USA, 2007. [Google Scholar]
- Gould, J.M. Alkaline peroxide delignification of agricultural residues to enhance enzymatic saccharification. Biotechnol. Bioeng. 1984, 26, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Savy, D.; Cozzolino, V.; Nebbioso, A.; Drosos, M.; Piccolo, A. Bioactivity of water-soluble lignins from biomass for energy on emergence and early growth of maize (Zea mays, L.). Plant Soil 2015, in press. [Google Scholar]
- Ertani, A.; Schiavon, M.; Altissimo, A.; Franceschi, C.; Nardi, S. Phenol-containing organic substances stimulate phenylpropanoid metabolism in Zea mays. J. Plant Nutr. Soil Sci. 2011, 174, 496–503. [Google Scholar] [CrossRef]
- Du Jardin, P. The Science of Plant Biostimulants-A Bibliographic Analysis. 30-CE0455515/00-96; European Commission: Brussels, Belgium, 2012; Available online: http://ec.europa.eu/enterprise/sectors/chemicals/files/fertilizers/final_report_bio_2012_en.pdf (accessed on 7 June 2015).
- Piccolo, A. The Supramolecular structure of humic substances. A novel understanding of humus chemistry and implications in soil Science. Adv. Agron. 2002, 75, 57–134. [Google Scholar]
- Canellas, L.P.; Olivares, F.L. Physiological responses to humic substances as plant growth promoter. Chem. Biol. Technol. Agric. 2014, 1, 1–11. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Zamarreño, A.M.; Aguirre, E.; Garnica, M.; Fuentes, M.; García-Mina, J.M. Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J. Plant Physiol. 2010, 167, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Francioso, O.; Tugnoli, V.; Righi, V.; Nardi, S. Effect of commercial lignosulfonate-humate on Zea mays L. metabolism. J. Agric. Food Chem. 2011, 59, 11940–11948. [Google Scholar] [CrossRef] [PubMed]
- Bălaş, A.; Popa, V.I. The influence of natural aromatic compounds on the development of Lycopersicon esculentum plantlets. BioResource 2007, 2, 363–370. [Google Scholar]
- Kesba, H.H.; El-Betagi, H.S. Biochemical changes in grape rootstocks resulted from humic acid treatments in relation to nematode infection. Asian Pac. J. Trop. Biomed. 2012, 2, 287–293. [Google Scholar] [CrossRef]
- García, A.C.; Santos, L.A.; Izquierdo, F.G.; Sperandio, M.V.; Castro, R.N.; Berbara, R.L.L. Vermicompost humic acids as an ecological pathway to protect rice plant against oxidative stress. Ecol. Eng. 2012, 47, 203–208. [Google Scholar] [CrossRef]
- Savy, D.; Piccolo, A. Physical-chemical characteristics of lignins separated from biomasses for second-generation ethanol. Biomass Bioenerg. 2014, 62, 58–67. [Google Scholar] [CrossRef]
- Fava, F.; Totaro, G.; Diels, L.; Reis, M.; Duarte, J.; Caioca, O.B.; Poggi-Varaldo, H.M.; Ferreira, B.S. Biowaste biorefinery in Europe: Opportunities and research & development needs. New Biotechnol. 2015, 32, 100–109. [Google Scholar] [CrossRef]
- Jahan, M.S.; Nasima Chowdhury, D.A.; Khalidul Islam, M.; Iqbal Moeiz, S.M. Characterization of lignin isolated from some nonwood available in Bangladesh. Bioresour. Technol. 2007, 98, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Hergert, H.L. Lignins: Occurrence, Formation, Structure and Reaction; Sarkanen, K.V., Ludwig, C.H., Eds.; Wiley-Interscience: New York, NY, USA, 1971; pp. 268–272. [Google Scholar]
- Nada, A.-A.M.A.; Yousef, M.A.; Shaffei, K.A.; Salah, A.M. Infrared spectroscopy of some treated lignins. Polym. Degrad. Stab. 1998, 62, 157–163. [Google Scholar] [CrossRef]
- Faix, O. Fourier Transfom Infrared Spectroscopy. In Methods in Lignin Chemistry; Lin, S.Y., Dence, C.W., Eds.; Springer Verlag: Heidelberg, Germany, 1992; pp. 81–109. [Google Scholar]
- Hatcher, P. Chemical structural studies of natural lignin by dipolar dephasing solid-state 13C nuclear magnetic resonance. Org. Geochem. 1987, 11, 31–38. [Google Scholar] [CrossRef]
- Chaa, L.; Joly, N.; Lequart, V.; Faugeron, C.; Cavrot, J.P.; Mollet, J.-C.; Martin, P.; Morvan, H. Isolation, characterization and valorization of hemicelluloses from Aristida pungens leaves as biomaterial. Carbohydr. Polym. 2008, 74, 597–602. [Google Scholar] [CrossRef]
- Piccolo, A.; Conte, P.; Cozzolino, A. Effects of mineral and monocarboxylic acids on the molecular association of dissolved humic substances. Eur. J. Soil Sci. 1999, 50, 687–694. [Google Scholar] [CrossRef]
- Verkade, J.G.; Quin, L.D. Phosphorus-31 NMR spectroscopy in stereochemical analysis of organic compounds and metal complexes. In Methods of Stereochemical Analysis; Grayson, M., Griffith, E.J., Eds.; Wiley-VCH: Deerfield Beach, FL, USA, 1987. [Google Scholar]
- Savy, D.; Mazzei, P.; Roque, R.; Nuzzo, A.; Bowra, S.; Santos, R. Structural recognition of lignin isolated from bioenergy crops by subcritical water: Ethanol extraction. Fuel Process. Technol. 2015. [Google Scholar] [CrossRef]
- Tohmura, S.; Argyropoulos, D.S. Determination of arylglycerol-β-aryl ethers and other linkages in lignins using DFRC/31P-NMR. J. Agric. Food Chem. 2001, 49, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Crestini, C.; Argyropoulos, D.S. Structural analysis of wheat straw lignin by quantitative 31P- and 2D-NMR spectroscopy. The occurrence of ester bonds and α-O-4 substructures. J. Agric. Food Chem. 1997, 45, 1212–1219. [Google Scholar] [CrossRef]
- Lourençao, A.; Neiva, D.M.; Gominho, J.; Curt, M.D.; Fernández, J.; Marques, A.V.; Pereira, H. Biomass production of four Cynara cardunculus clones and lignin composition analysis. Biomass Bioenergy 2015, 76, 86–95. [Google Scholar] [CrossRef]
- Savy, D.; Nebbioso, A.; Mazzei, P.; Drosos, M.; Piccolo, A. Molecular composition of water-soluble lignins separated from different non-food biomasses. Fuel Process. Technol. 2015, 131, 175–181. [Google Scholar] [CrossRef]
- You, T.-T.; Mao, J.; Yuan, T.; Wen, J.-L.; Xu, F. Structural elucidation of the lignins from stems and foliage of Arundo donax Linn. J. Agric. Food Chem. 2013, 61, 5361–5370. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, Y.; Kondo, R.; Sakai, K.; Imamura, H. The difference of reactivity between syringyl lignin and guaiacyl lignin in alkaline systems. Holzforschung 1995, 19, 423–428. [Google Scholar] [CrossRef]
- Sultanov, V.; Wallis, A. Reactivities of guaiacyl and syringyl lignin model phenols towards oxidation with oxygen-alkali. J. Wood Chem. Technol. 1991, 11, 291–305. [Google Scholar] [CrossRef]
- Brebu, M.; Vasile, C. Thermal degradation of wood—A review. Cellul. Chem. Technol. 2010, 44, 353–363. [Google Scholar]
- Hoareau, W.; Trindade, W.G.; Siegmund, B.; Castellan, A.; Follini, E. Sugar cane bagasse and curaua lignins oxidized by chlorine dioxide and reacted with furfuryl alcohol: Characterization and stability. Polym. Degrad. Stab. 2004, 86, 567–576. [Google Scholar] [CrossRef]
- Sun, R.; Lu, Q.; Sun, X.F. Physico-chemical and thermal characterization of lignins from Caligonum monogoliacum and Tamarix spp. Polym. Degrad. Stab. 2001, 72, 229–238. [Google Scholar] [CrossRef]
- Yang, H.; Yan, R.; Chen, H.; Lee, D.H.; Zheng, C. Characteristics of hemicellulose, cellulose and lignin pyrolysis. Fuel 2007, 86, 1781–1788. [Google Scholar] [CrossRef]
- Piccolo, A.; Conte, P.; Trivellone, E.; van Lagen, B.; Buurman, P. Reduced heterogeneity of a lignite humic acid by preparative HPSEC following interaction with an organic acid. Characterization of size-separates by PYR-GC-MS and 1H-NMR spectroscopy. Environ. Sci. Technol. 2002, 36, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Dobbss, L.B.; Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Peres, L.E.P.; Azevedo, M.; Spaccini, R.; Piccolo, A.; Façanha, A.R. Bioactivity of chemically transformed humic matter from vermicompost on plant root growth. J. Agric. Food Chem. 2010, 58, 3681–3688. [Google Scholar] [CrossRef] [PubMed]
- Popa, V.I.; Dumitru, M.; Volfa, I.; Anghel, N. Lignin and polyphenols as allelochemicals. Ind. Crops Prod. 2008, 27, 144–149. [Google Scholar] [CrossRef]
- Tanase, C.; Boz, I.; Stingu, A.; Volf, I.; Popa, V.I. Physiological and biochemical responses induced by spruce bark aqueous extract and deuterium depleted water with synergistic action in sunflower (Helianthus annuus L.) plants. Ind. Crops Prod. 2014, 60, 160–167. [Google Scholar] [CrossRef]
- Nardi, S.; Pizzeghello, D.; Bragazza, L.; Gedol, R. Low-molecular-weight organic acids and hormone-like activity of dissolved organic matter in two forest soils in N Italy. J. Chem. Ecol. 2003, 29, 1549–1564. [Google Scholar] [CrossRef] [PubMed]
- Pizzeghello, D.; Zanella, A.; Carletti, P.; Nardi, S. Chemical and biological characterization of dissolved organic matter from silver fir and beech forest soils. Chemosphere 2006, 65, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Tomkinson, J.; Wang, S.; Zhu, W. Characterization of lignins from wheat straw by alkaline peroxide treatment. Polym. Degrad. Stab. 2000, 67, 101–109. [Google Scholar] [CrossRef]
- Hatzakis, E.; Dagounakis, G.; Dais, P. A facile NMR method for the quantification of total free and esterified sterols in virgin olive oil. Food Chem. 2010, 122, 346–352. [Google Scholar] [CrossRef]
- Zwierzak, A. Cyclic organophosphorus compounds. I. Synthesis and infrared spectral studies of cyclic hydrogen phosphites and thiophosphites. Can. J. Chem. 1967, 45, 2501–2512. [Google Scholar] [CrossRef]
- Granata, A.; Argyropouls, D.S. 2-Chloro-4,4,5,5-tetramethyl-1,3,2-dioxaphospholane, a reagent for the accurate determination of the uncondensed and condensed phenolic moieties in lignins. J. Agric. Food Chem. 1995, 43, 1538–1544. [Google Scholar] [CrossRef]
- Sample Availability: Samples of any compounds are not available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savy, D.; Cozzolino, V.; Vinci, G.; Nebbioso, A.; Piccolo, A. Water-Soluble Lignins from Different Bioenergy Crops Stimulate the Early Development of Maize (Zea mays, L.). Molecules 2015, 20, 19958-19970. https://doi.org/10.3390/molecules201119671
Savy D, Cozzolino V, Vinci G, Nebbioso A, Piccolo A. Water-Soluble Lignins from Different Bioenergy Crops Stimulate the Early Development of Maize (Zea mays, L.). Molecules. 2015; 20(11):19958-19970. https://doi.org/10.3390/molecules201119671
Chicago/Turabian StyleSavy, Davide, Vincenza Cozzolino, Giovanni Vinci, Antonio Nebbioso, and Alessandro Piccolo. 2015. "Water-Soluble Lignins from Different Bioenergy Crops Stimulate the Early Development of Maize (Zea mays, L.)" Molecules 20, no. 11: 19958-19970. https://doi.org/10.3390/molecules201119671