
Methanolic Extract of Ganoderma lucidum Induces Autophagy of AGS Human Gastric Tumor Cells

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
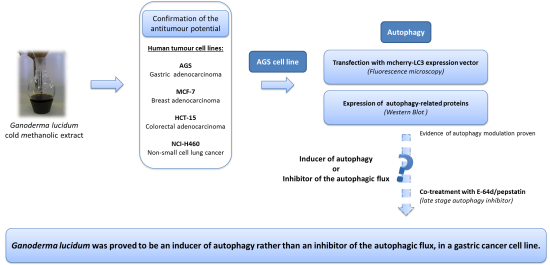
2. Results and Discussion
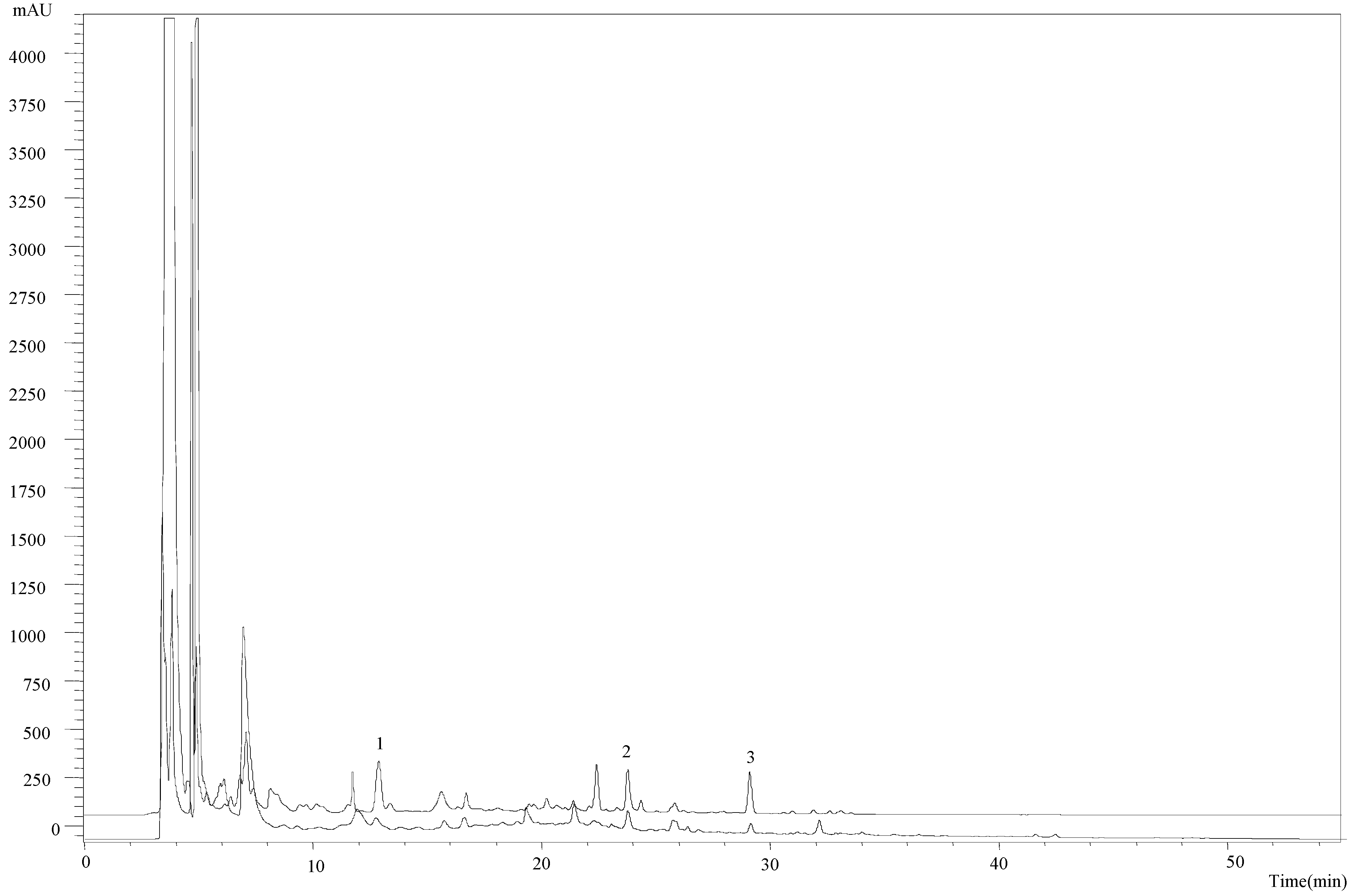
2.1. Chemical Characterization of the Methanolic Extract
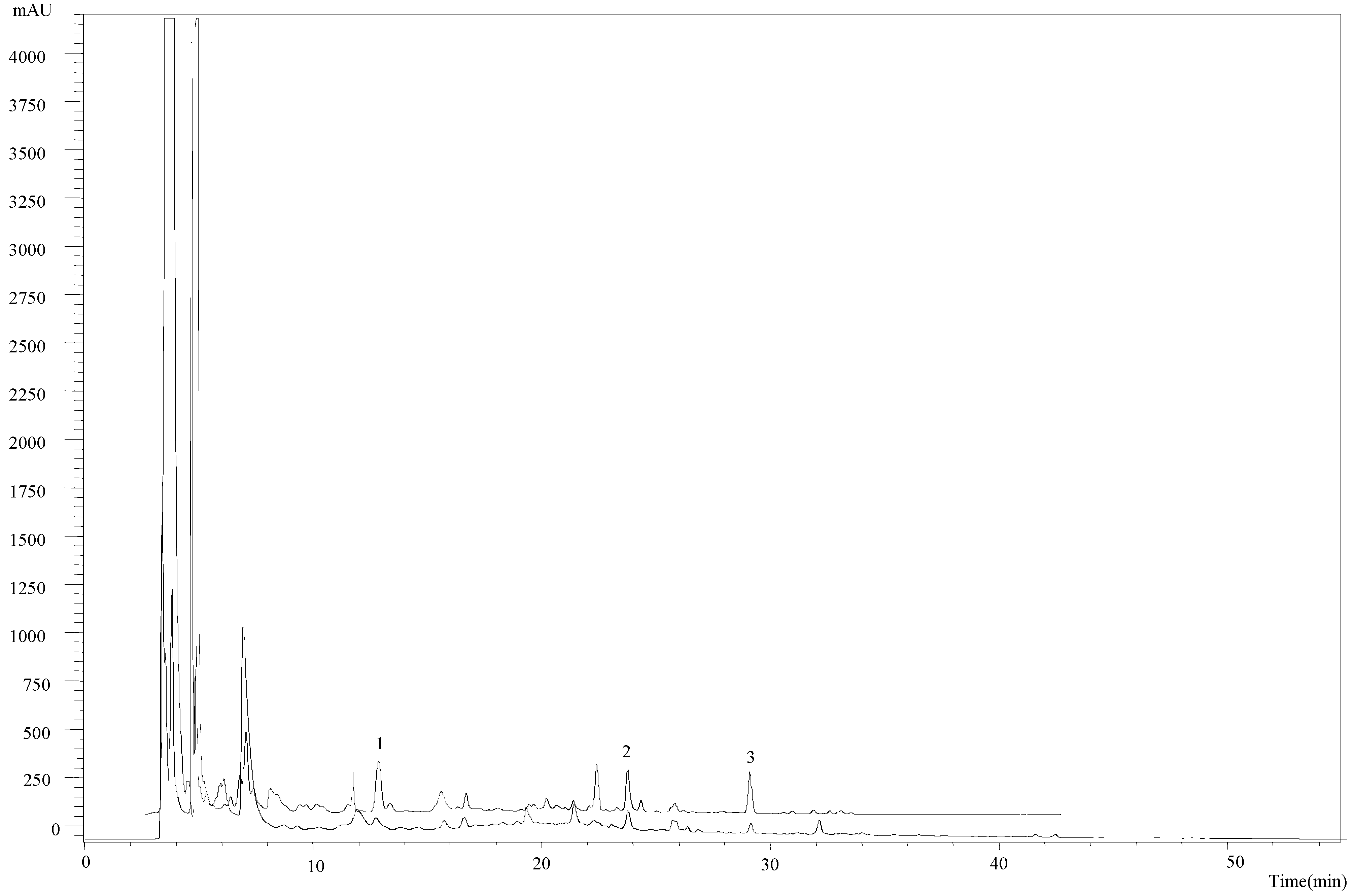
2.2. Antitumor Potential of the New Obtained Extract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ganoderma lucidum Methanolic Extract | ||||
|---|---|---|---|---|
| GI50 (µg/mL) | NCI-H460 | HCT-15 | MCF-7 | AGS |
| 77.9 ± 3.3 | 70.0 ± 2.4 | 67.2 ± 3.1 | 66.6 ± 4.4 | |
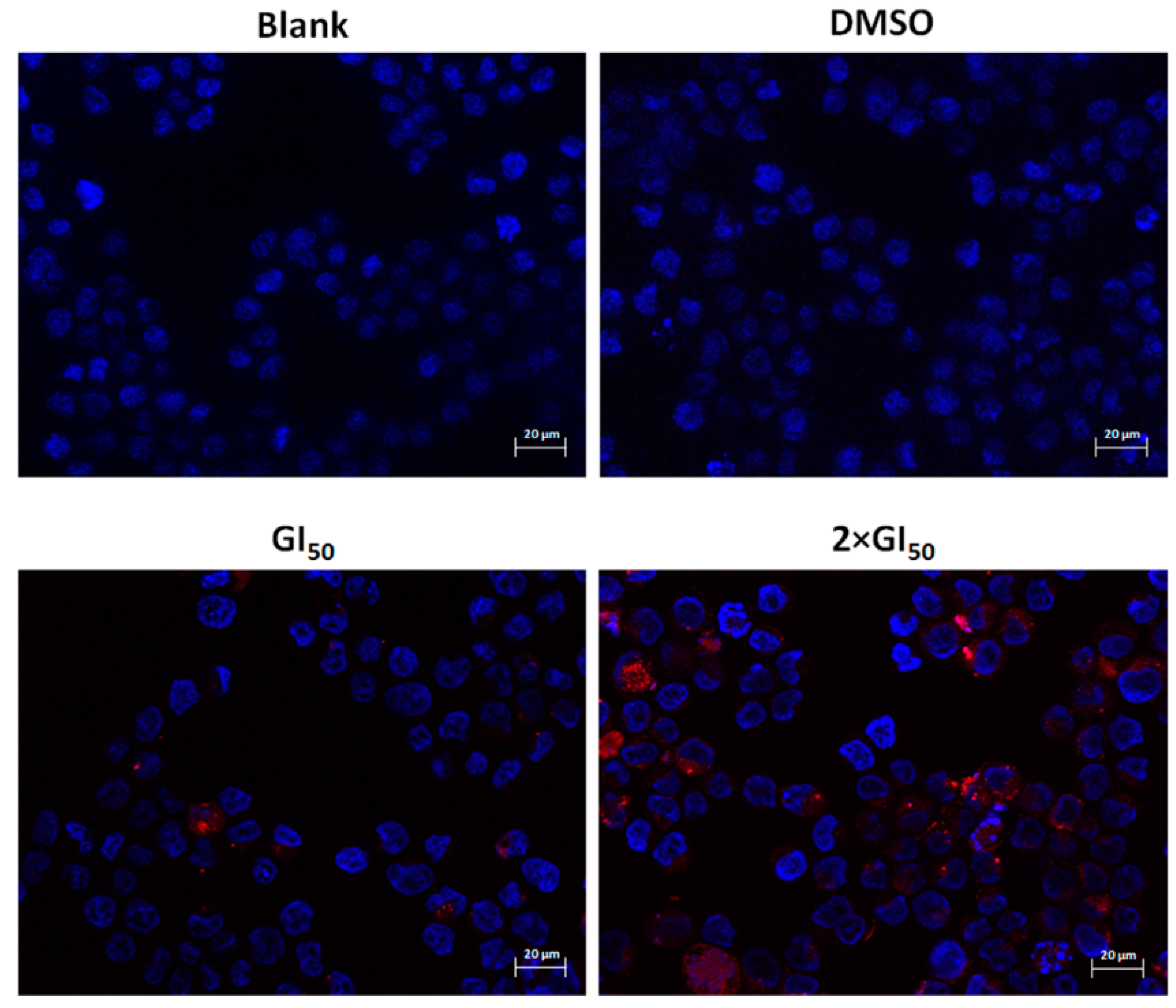
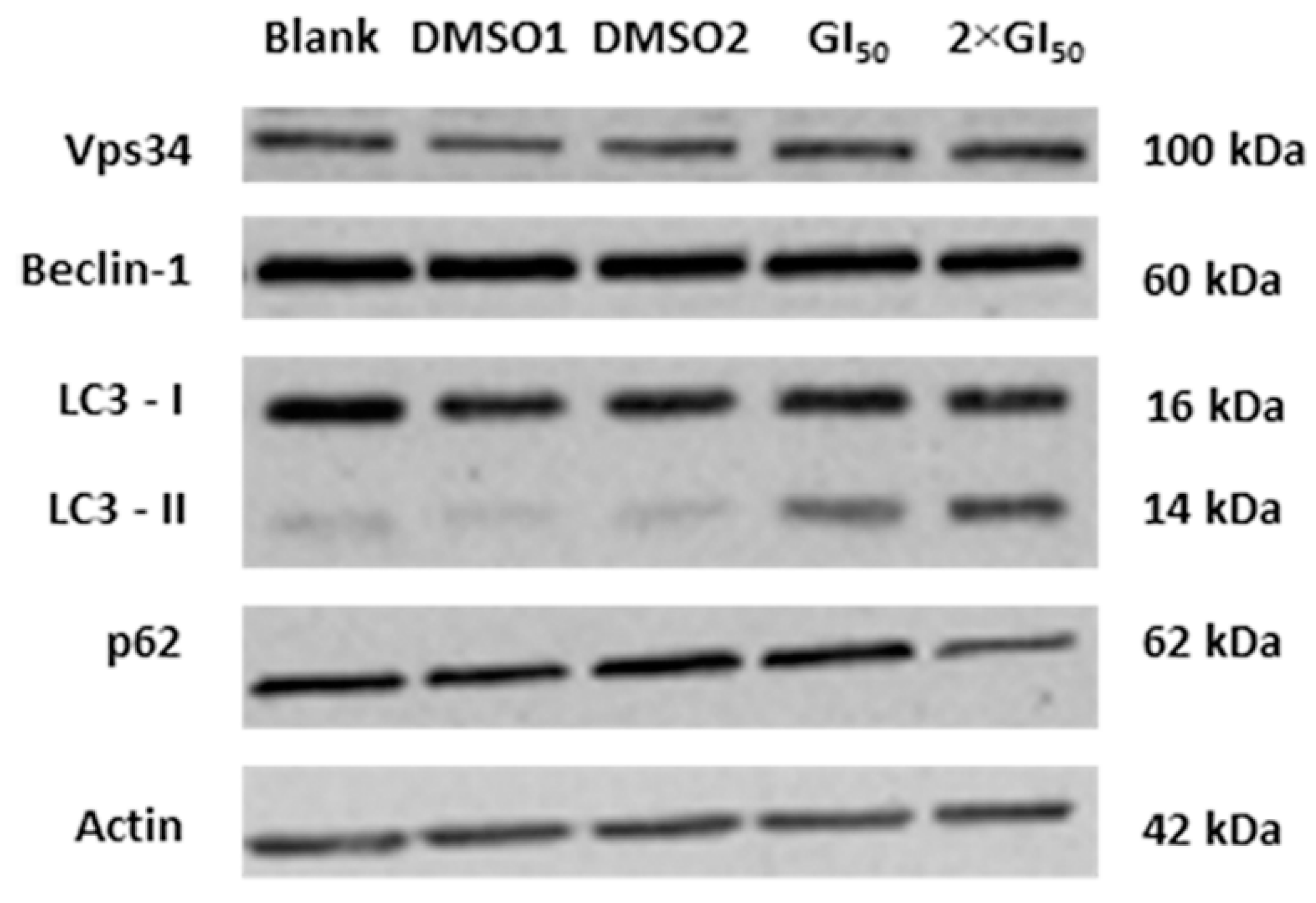
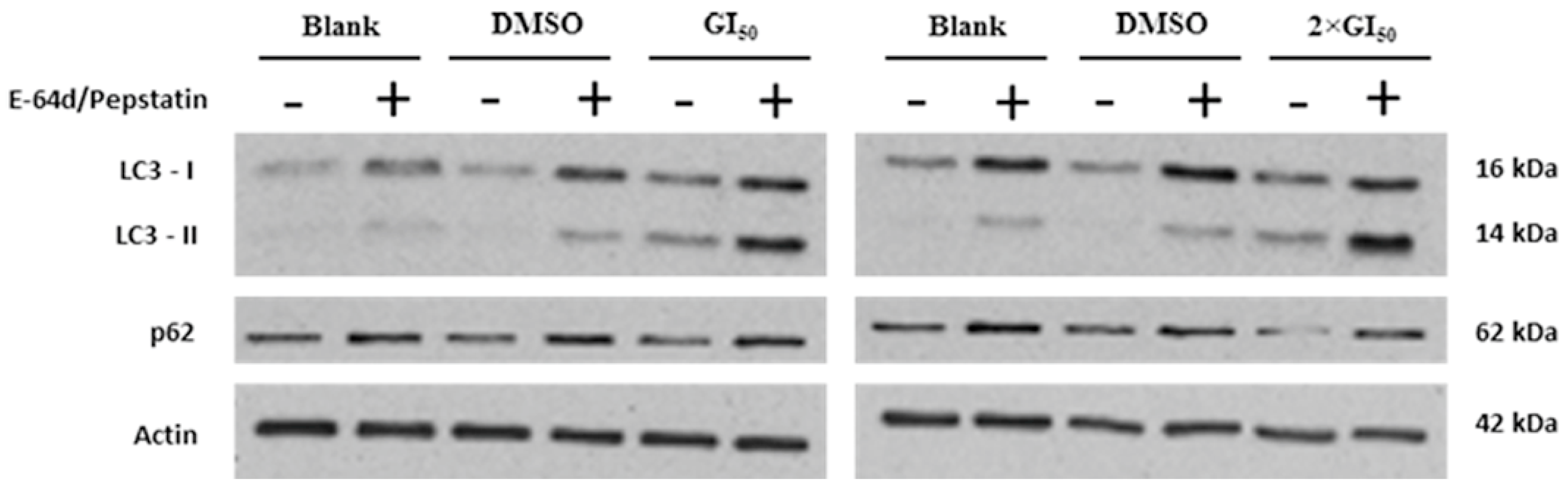
2.3. Effect of G. lucidum Methanolic Extract in AGS Cellular Autophagy

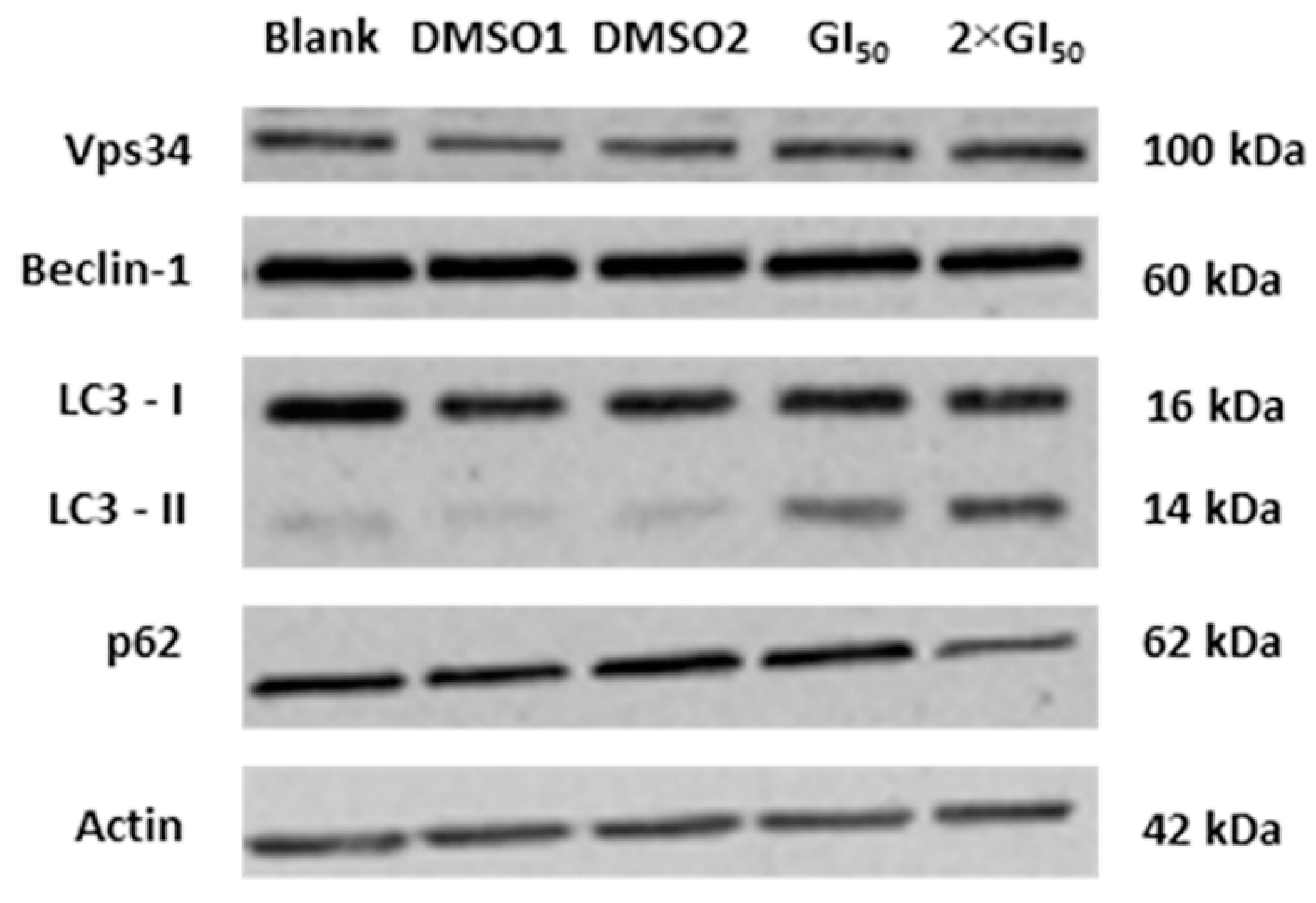
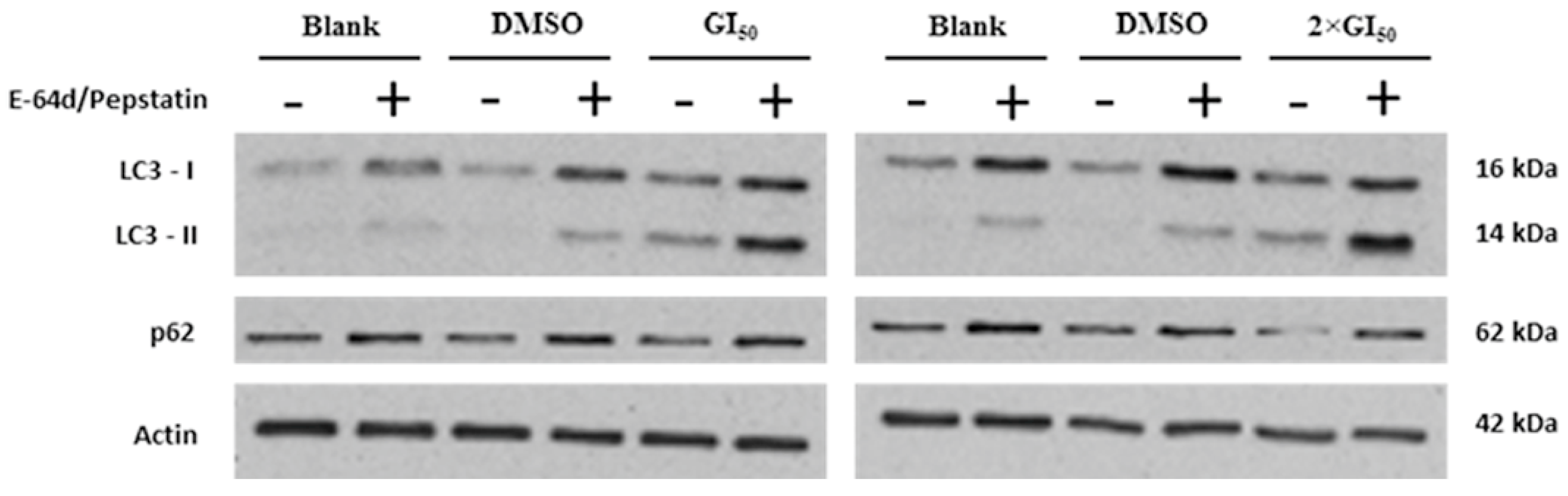

3. Experimental Section
3.1. Collection and Sample Preparation
3.2. Preparation and Chemical Characterization of Ganoderma lucidum Methanolic Extract
3.3. Screening of the Growth Inhibitory Activity in Human Tumor Cell Lines
3.4. Transfection with LC3-mCherry Expression Vector
3.5. Expression of Autophagy-Related Proteins
3.6. Treatment with Autophagy Inhibitors E-64d/Pepstatin
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.L.; Ooi, J.P.; Ismail, N.; Moad, A.I.H.; Muhammad, T.S.T. Programmed cell death pathways and current antitumor targets. Pharm. Res. 2009, 26, 1547–1560. [Google Scholar] [CrossRef]
- Yang, Z.J.; Chee, C.E.; Huang, S.; Sinicrop, F.A. The role of autophagy in cancer: Therapeutic implications. Mol. Cancer Ther. 2011, 10, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.C.F.R.; Vaz, J.A.; Vasconcelos, M.H.; Martins, A. Compounds from wild mushrooms with antitumor potential. Anticancer Agents Med. Chem. 2010, 10, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; Goyal, A. Recent developments in mushrooms as anti-cancer therapeutics: A review. 3 Biotech 2012, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.S.; Morales, P.; Ferreira, I.C.F.R.; Vasconcelos, M.H. Mushrooms as a source of compounds that induce programmed cell death in tumor cells. In Recent Progress in Medicinal Plants; Govil, J.N., Ed.; Studium Press LLC: Houston, TX, USA, 2015; Volume 43—Phytotherapeutics II, in press. [Google Scholar]
- Harhaji Trajković, L.M.; Mijatović, S.A.; Maksimović-Ivanić, D.D.; Stojanović, I.D.; Momčilović, M.B.; Tufegdžić, S.J.; Maksimović, V.M.; Marjanovi, Ž.S.; Stošić-Grujičić, S.D. Anticancer properties of Ganoderma lucidum methanol extracts in vitro and in vivo. Nutr. Cancer 2009, 61, 696–707. [Google Scholar]
- Calviño, E.; Manjón, J.L.; Sancho, P.; Tejedor, M.C.; Herráez, A.; Diez, J.C. Ganoderma lucidum induced apoptosis in NB4 human leukemia cells: Involvement of Akt and Erk. J. Ethnopharmacol. 2010, 128, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.C.F.R.; Heleno, S.A.; Reis, F.S.; Stojković, D.; Queiroz, M.J.R.P.; Vasconcelos, M.H.; Soković, M. Chemical features of Ganoderma polysaccharides with antioxidant, antitumor and antimicrobial activities. Phytochemistry 2015, 114, 38–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thyagarajan, A.; Jedinak, A.; Nguyen, H.; Terry, C.; Baldridge, L.A.; Jiang, J.; Sliva, D. Triterpenes from Ganoderma lucidum induce autophagy in colon cancer through the inhibition of p38 mitogen-activated kinase (p38 MAPK). Nutr. Cancer 2010, 62, 630–640. [Google Scholar] [PubMed]
- Hossain, A.; Radwan, F.F.Y.; Doonan, B.P.; God, J.M.; Zhang, L.; Bell, P.D.; Haque, A. A possible cross-talk between autophagy and apoptosis in generating an immune response in melanoma. Apoptosis 2012, 17, 1066–1078. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Li, H.; Zhou, H.; Zhang, S.; Liu, Z.; Zhou, Q.; Sun, F. Recombinant Lz-8 from Ganoderma lucidum induces endoplasmic reticulum stress-mediated autophagic cell death in SGC-7901 human gastric cancer cells. Oncol. Rep. 2012, 27, 1079–1089. [Google Scholar] [PubMed]
- Oliveira, M.; Reis, F.S.; Sousa, D.; Tavares, C.; Lima, R.T.; Ferreira, I.C.F.R.; dos Santos, T.; Vasconcelos, M.H. A methanolic extract of Ganoderma lucidum fruiting body inhibits the growth of a gastric cancer cell line and affects cellular autophagy and cell cycle. Food Funct. 2014, 5, 1389–1394. [Google Scholar] [CrossRef] [PubMed]
- Funderburk, S.F.; Wang, Q.J.; Yue, Z. The Beclin 1-VPS34 complex—At the crossroads of autophagy and beyond. Trends Cell. Biol. 2010, 20, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Yoshimori, T.; Levine, B. Methods in mammalian autophagy research. Cell 2010, 140, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Seca, H.; Lima, R.T.; Lopes-Rodrigues, V.; Guimarães, J.E.; Almeida, G.M.; Vasconcelos, M.H. Targeting miR-21 induces autophagy and chemosensitivity of leukemia cells. Curr. Cancer Drug Targets 2013, 14, 1135–1143. [Google Scholar] [CrossRef]
- Ichimura, Y.; Komatsu, M. Selective degradation of p62 by autophagy. Semin. Immunopathol. 2010, 32, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Cerniglia, G.J.; Karar, J.; Tyagi, S.; Christofidou-Solomidou, M.; Rengan, R.; Koumenis, C.; Maity, A. Inhibition of autophagy as a strategy to augment radiosensitization by the dual phosphatidylinositol 3-kinase/mammalian target of rapamycin inhibitor NVP-BEZ235. Mol. Pharmacol. 2012, 82, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
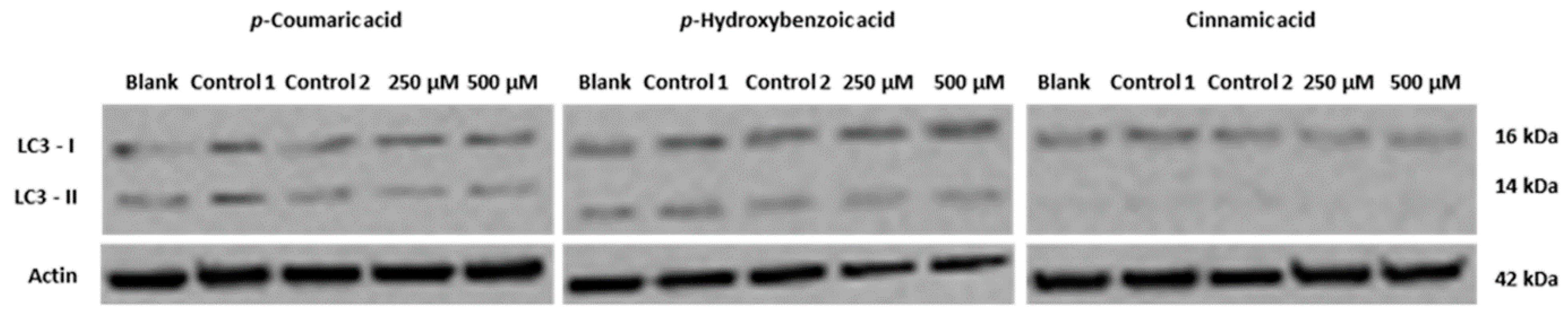
- Heleno, S.A.; Ferreira, I.C.F.R.; Esteves, A.P.; Ćirić, A.; Glamočlija, J.; Martins, A.; Soković, M.; Queiroz, M.J.R.P. Antimicrobial and demelanizing activity of Ganoderma lucidum extract, p-hydroxybenzoic and cinnamic acids and their synthetic acetylated glucuronide methyl esters. Food Chem. Toxicol. 2013, 58, 95–100. [Google Scholar] [CrossRef]
- Cai, S.; Wang, O.; Wang, M.; He, J.; Wang, Y.; Zhang, D.; Zhou, F.; Ji, B. In vitro inhibitory effect on pancreatic lipase activity of subfractions from ethanol extracts of fermented oats (Avena sativa L.) and synergistic effect of three phenolic acids. J. Agric. Food Chem. 2012, 60, 7245–7251. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-J.; Lee, C.-C.; Shih, Y.-L.; Lin, T.-Y.; Wang, S.-H.; Lin, Y.-F.; Shih, C.-M. Resveratrol enhances the therapeutic effect of temozolomide against malignant glioma in vitro and in vivo by inhibiting autophagy. Free Radic. Biol. Med. 2012, 52, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Vaz, J.A.; Almeida, G.M.; Ferreira, I.C.F.R.; Martins, A.; Vasconcelos, M.H. Clitocybe alexandri extract induces cell cycle arrest and apoptosis in a lung cancer cell line: Identification of phenolic acids with cytotoxic potential. Food Chem. 2012, 132, 482–486. [Google Scholar] [CrossRef]
- Weng, C.-J.; Yen, G.-C. Chemopreventive effects of dietary phytochemicals against cancer invasion and metastasis: Phenolic acids, monophenol, polyphenol, and their derivatives. Cancer Treat. Rev. 2012, 38, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Gonzáles-Vallinas, M.; Gonzáles-Catejón, M.; Rodríguez-Casado, A.; de Molina, A.R. Dietary phytochemicals in cancer prevention and therapy: A complementary approach with promising perspectives. Nutr Rev. 2013, 71, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Barros, L.; Martins, A.; Queiroz, M.J.R.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Fruiting body, spores and in vitro produced mycelium of Ganoderma lucidum from Northeast Portugal: A comparative study of the antioxidant potential of phenolic and polysaccharidic extracts. Food Res. Int. 2012, 46, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Monks, A.; Scudiero, D.; Skehan, P.; Shoemaker, R.; Paull, K.; Vistica, D.; Hose, C.; Langley, J.; Cronise, P.; Vaigro-Wolff, A.; et al. Feasibility of a high-flux anticancer drug screen using a diverse panel of cultured human tumor cell lines. J. Natl. Cancer Inst. 1991, 83, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.A.; Outzen, H.; Overvatn, A.; Bjorkoy, G.; Johansen, T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–24145. [Google Scholar] [CrossRef] [PubMed]
- Birame, B.M.; Jigui, W.; Fuxian, Y.; Shuang, W.; Li, Y.D.; Jiazeng, S.; Zhili, L.; Bao, Y.; Weiquan, L. Co-expression of apoptin (VP3) and antibacterial peptide cecropin B mutant (ABPS1) genes induce higher rate of apoptosis in HepG2 and A375 cell lines. Afr. J. Biotechnol. 2012, 11, 10981–10988. [Google Scholar]
- Lima, R.T.; Barron, G.A.; Grabowska, J.A.; Bermano, G.; Kaur, S.; Roy, N.; Vasconcelos, M.H.; Lin, P.K.T. Cytotoxicity and cell death mechanisms induced by a novel bisnaphthalimidopropyl derivative against the NCI-H460 non-small lung cancer cell line. Anticancer Agents Med. Chem. 2013, 13, 414–421. [Google Scholar] [PubMed]
- Sample Availability: Mushroom voucher specimens are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reis, F.S.; Lima, R.T.; Morales, P.; Ferreira, I.C.F.R.; Vasconcelos, M.H. Methanolic Extract of Ganoderma lucidum Induces Autophagy of AGS Human Gastric Tumor Cells. Molecules 2015, 20, 17872-17882. https://doi.org/10.3390/molecules201017872
Reis FS, Lima RT, Morales P, Ferreira ICFR, Vasconcelos MH. Methanolic Extract of Ganoderma lucidum Induces Autophagy of AGS Human Gastric Tumor Cells. Molecules. 2015; 20(10):17872-17882. https://doi.org/10.3390/molecules201017872
Chicago/Turabian StyleReis, Filipa S., Raquel T. Lima, Patricia Morales, Isabel C. F. R. Ferreira, and M. Helena Vasconcelos. 2015. "Methanolic Extract of Ganoderma lucidum Induces Autophagy of AGS Human Gastric Tumor Cells" Molecules 20, no. 10: 17872-17882. https://doi.org/10.3390/molecules201017872