2.1. Oligosaccharides Produced from Cellulose Derivative
The acid hydrolysis of CMC was analyzed to characterize a non-enzymatic profile. As a random depolymerization was expected, it was also carried out to evaluate the ionization efficiency of the hydrolysis products because no substituted standard compounds were available. In the case of derivative cellulose substrate, a discrimination against molecules with few substituents was reported for MALDI-TOF MS analysis of CMC [
21]. The acid hydrolysis profile of CMC (
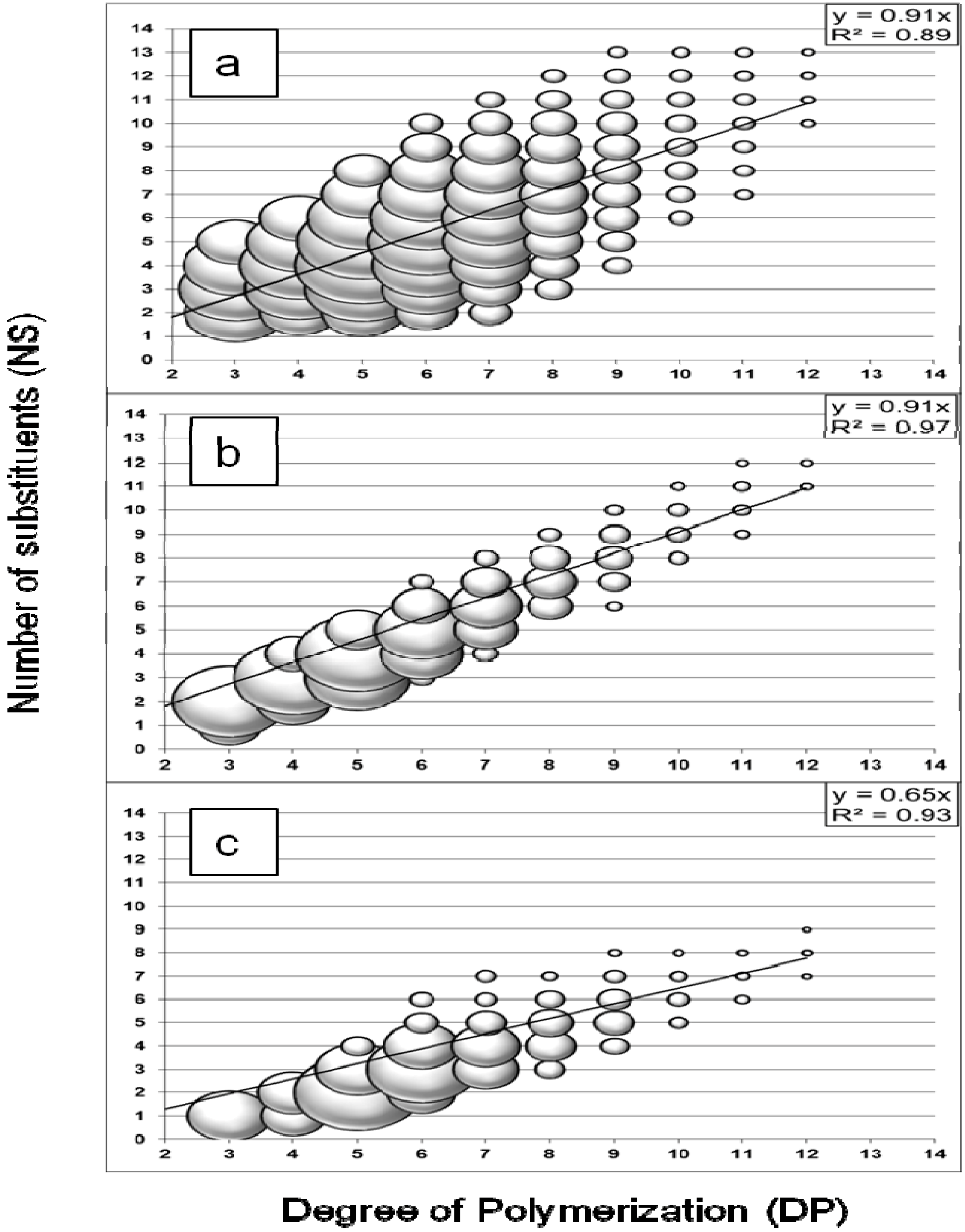
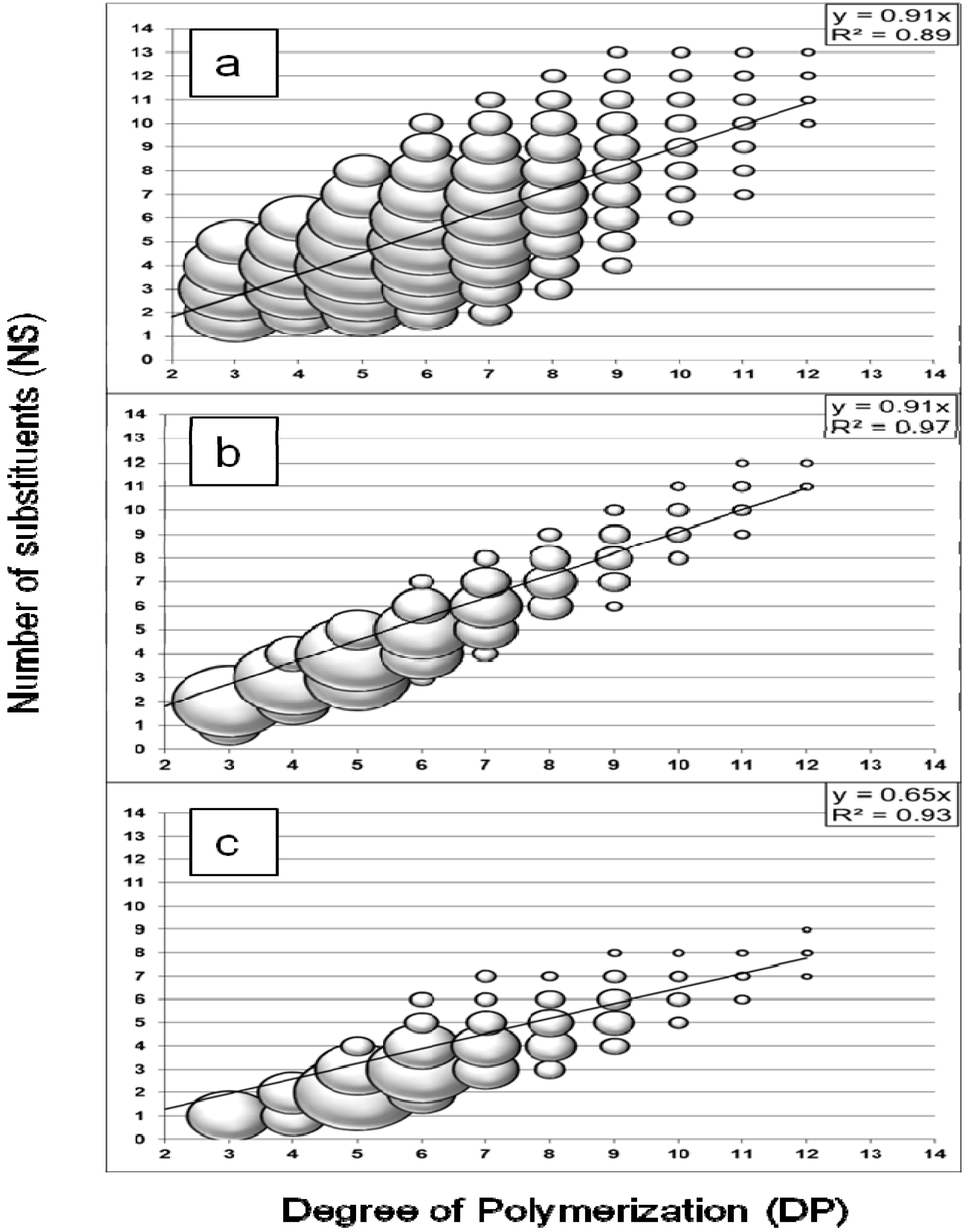
Figure 1a) showed a large distribution of oligosaccharides with DPs from three to 12 and an average number of substituents (NS) range, with DP-dependency, from NS = DP − 5 to NS = DP + 3. The results demonstrated no significant discrimination against oligomers with few substituents as they were detected with relative intensity (represented by the spheric volume of the markers) similar to oligomers with the highest number of substituents. However, the intensity decreased for oligomers with DP higher than eight because of the discrimination against larger oligomers for MALDI analysis in positive mode [
21]. It was suggested that the cationisation process from adduct formation favors the smaller molecules, which are likely more soluble and able to migrate to the surface of the sample spot and generate a signal. The relationship between DP and NS values was demonstrated with a linear regression model (least squares method), with R
2 = 0.89 and a slope value of 0.91. In the case of acid hydrolysis, this provided an interesting method to calculate the degree of substitution (DS) of CMC
i.e. the average number of carboxymethyl substituent per unit of glucose. An average similar DS value (0.88 ± 0.02, RSD 2.5%, n = 4) was calculated with the method (DS
Level Off) proposed by Enebro and Karlsson [
25]. This method (measuring DS where the curve levels off) was based on the fact that the difference between the measured and the true DS should decrease with increasing DP, considering the lower ionization efficiency for oligomers with few substituents discussed by Momcilovic
et al. [
21]. However, the estimated values calculated with both methods remained higher than the DS value of 0.7 specified by the supplier of the CMC used in this study. Overestimation when measuring the DS in CMC by MALDI-TOF MS was also discussed by Enebro and Karlsson [
25]. Additionally to the influence of the cationisation process dependent on the total number of substituents, they suggested an influence of the homogenization of the sample spot and the crystal size as they obtained better estimation with spectra acquisition centered in the target spot. In this study, part of the overestimation of DS could be explained because of the automated random shooting of the entire spot applied for all analyses.
Figure 1.
Comparison of carboxymethyl cellulose (CMC) hydrolysis profiles obtained with (a) trifluoroacetic acid (TFA) 2M, (b) a commercial cellulase from Trichoderma reesei, (c) termite hindgut homogenates (Reticulitermes santonensis). The oligosaccharides produced were analyzed by MALDI-TOF MS and displayed with markers according to their degree of polymerization (DP) and total number of substituents (NS), identified from m/z values. The relative intensity of each compound was represented by the spherical volume of the marker.
Figure 1.
Comparison of carboxymethyl cellulose (CMC) hydrolysis profiles obtained with (a) trifluoroacetic acid (TFA) 2M, (b) a commercial cellulase from Trichoderma reesei, (c) termite hindgut homogenates (Reticulitermes santonensis). The oligosaccharides produced were analyzed by MALDI-TOF MS and displayed with markers according to their degree of polymerization (DP) and total number of substituents (NS), identified from m/z values. The relative intensity of each compound was represented by the spherical volume of the marker.
The method was validated for the analysis of enzymatic hydrolysis using a commercial
T. reesei cellulase (
Figure 1b). The DPs of the measured oligosaccharides ranged from three to twelve and the NS values were also DP-dependent, but within a limited range from NS = DP − 2 to NS = DP + 1, on average. Additionally, a higher relative intensity was obtained for some compounds. This preferential formation of hydrolysis products reflected the selectivity of an enzymatic system, for which the hydrolysis reaction was limited by the presence of substituents on the substrate, probably from steric hindrance at the enzyme’s active site. A linear correlation was also demonstrated with a higher coefficient of determination (R
2 = 0.97). The slope value was similar to the acid hydrolysis result (0.91).
The hydrolysis profile obtained with the termite hindgut homogenates and CMC substrate is shown in
Figure 1c. The profile presented the characteristics of an enzymatic hydrolysis profile: the NS range for similar DP was limited to a few values and some compounds presented higher relative intensity. The measured oligosaccharides had DPs from three to twelve with a NS range from NS = DP − 5 to NS = DP − 2, on average. The relationship between DP and NS values was linear (R
2 = 0.93) but the slope value (0.65) was lower than for the other profiles.
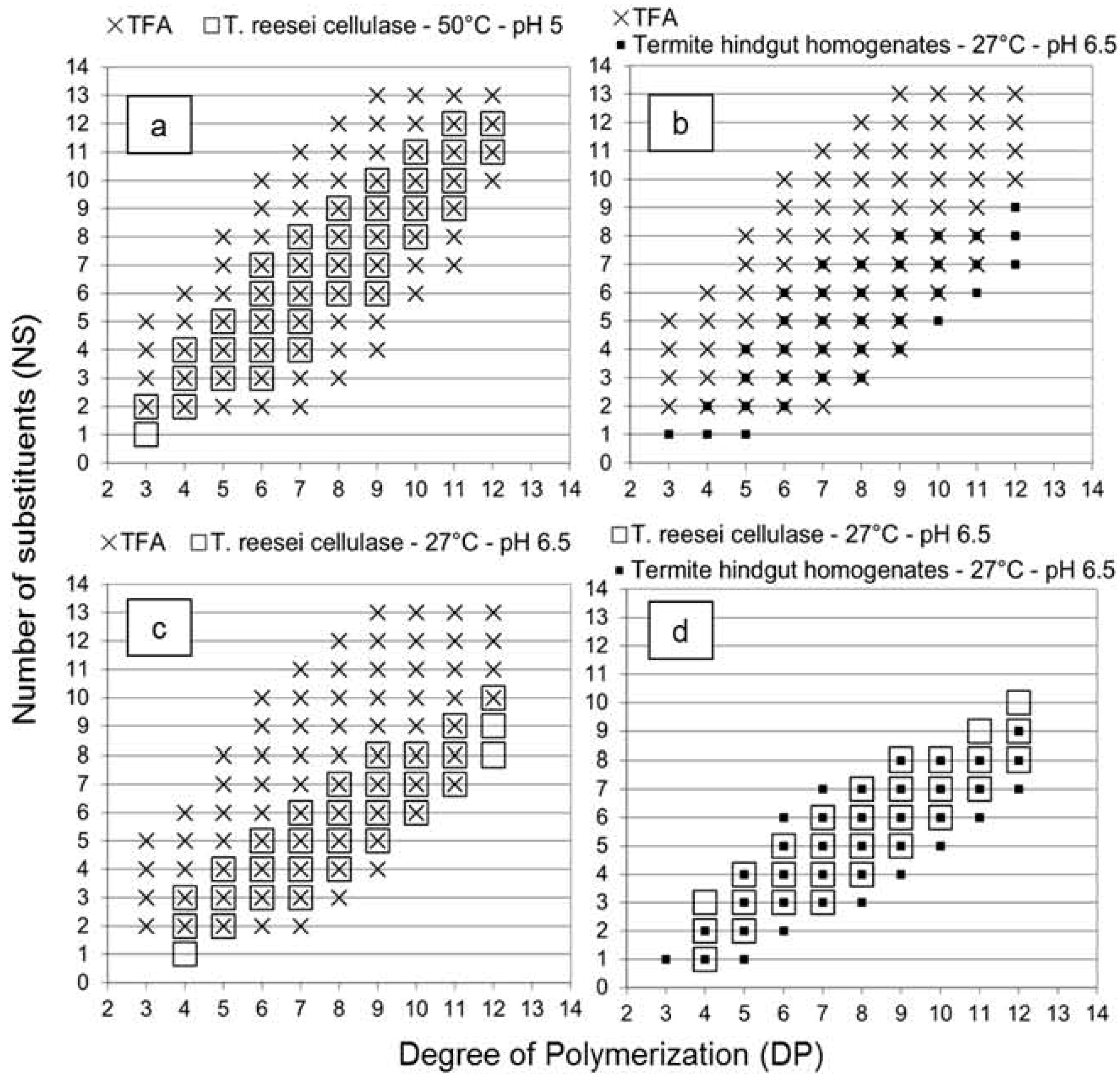
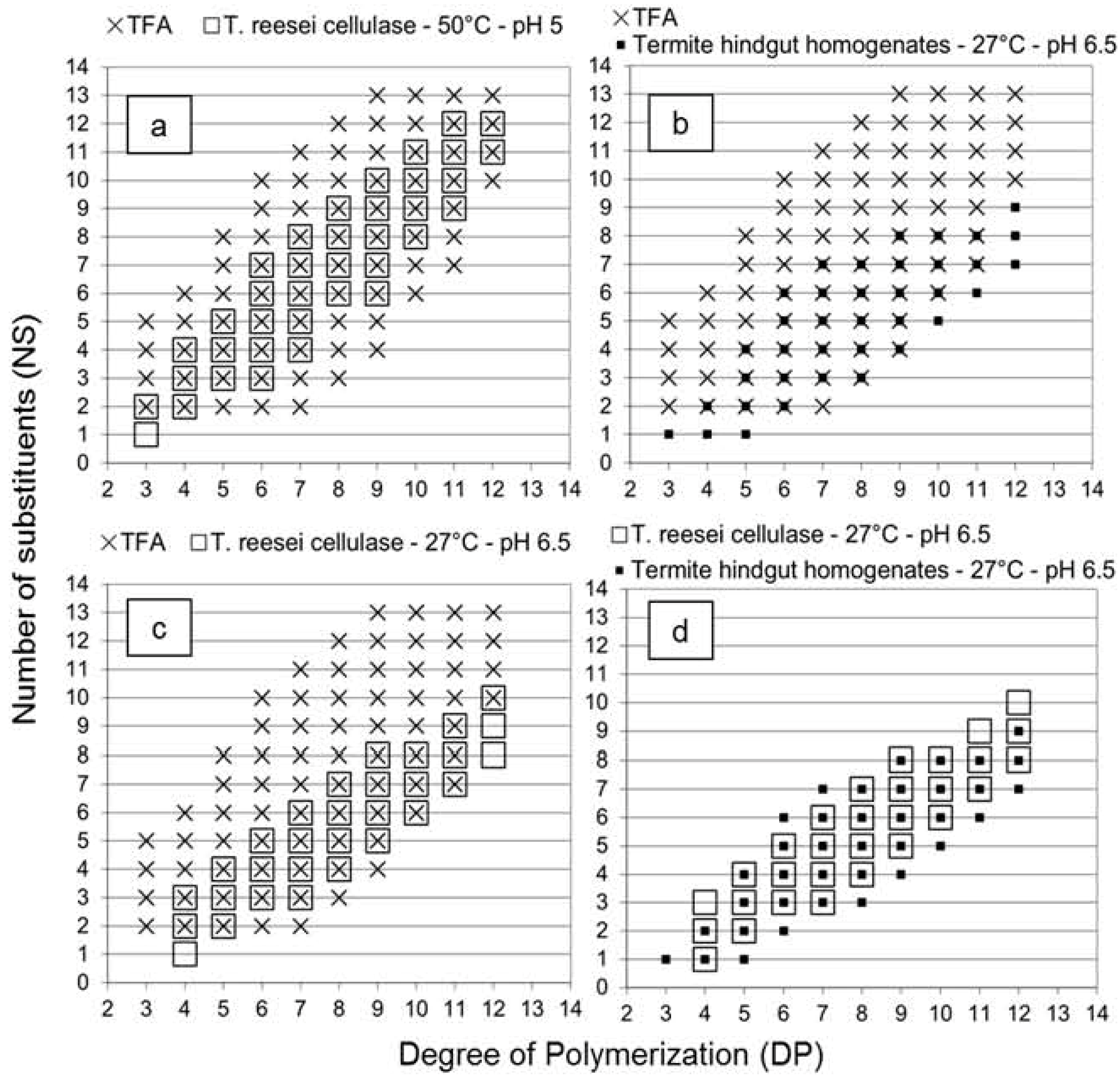
It is interesting to note that similar slope values were obtained for acid hydrolysis and
T. reesei cellulase profiles, despite the enzymatic selectivity of the
T. reesei cellulase. The explanation was clearly demonstrated in
Figure 2 with the overlay of the different graphs. Oligomers with fewer substituents were not detected in the
T. reesei cellulase profile in comparison with the acid hydrolysis profile (
Figure 2a). In the termite profile, oligomers with fewer NS values similar to the acid hydrolysis profile were detected (
Figure 2b), and as the enzymatic selectivity inhibited production of oligomers with high NS value, the slope value of the termite profile was lower (0.65), as expected.
Figure 2.
The enzyme selectivity of the Trichoderma reesei cellulase (empty squares) and the termite hindgut homogenates (black squares) can be evaluated based on the number of substituents (NS) obtained for each degree of polymerization (DP) of the oligosaccharides produced from carboxymethyl cellulose (CMC) hydrolysis. The T. reesei cellulase was tested with different conditions of pH and temperature and trifluoroacetic acid (TFA) 2 M was used for comparison with non-enzymatic hydrolysis reference profile (a–d). The oligosaccharides were analyzed by MALDI-TOF MS.
Figure 2.
The enzyme selectivity of the Trichoderma reesei cellulase (empty squares) and the termite hindgut homogenates (black squares) can be evaluated based on the number of substituents (NS) obtained for each degree of polymerization (DP) of the oligosaccharides produced from carboxymethyl cellulose (CMC) hydrolysis. The T. reesei cellulase was tested with different conditions of pH and temperature and trifluoroacetic acid (TFA) 2 M was used for comparison with non-enzymatic hydrolysis reference profile (a–d). The oligosaccharides were analyzed by MALDI-TOF MS.
The high slope value of the
T. reesei cellulase profile suggested an exoglucanase or β-glucosidase activity on oligomers produced by the endoglucanase activity (see
Table 1). Enzymes were likely able to find and hydrolyze one or two unsubstituted glucose units at the extremity of the produced oligomers, resulting from the selective endoglucanase activity. Consequently, the DP value of each oligomer decreased of one or two units while the NS value remained constant, increasing the slope of the linear relationship between DP and NS values. This additional activity influenced the distribution of the oligomers produced and compensated the decrease of the slope value due to the endoglucanase selectivity. Results obtained with the
T. reesei cellulase at 27 °C and pH 6.5 (
Figure 2c) supported this hypothesis, as oligomers with fewer NS values similar to the acid hydrolysis profile were detected and a lower slope was obtained. This suggested that a temperature of 27 °C and a pH value of 6.5 were not optimal conditions for the exoglucanase or β-glucosidase activity observed. Therefore, without the influence of these activities on the
T. reesei profile,
Figure 2d demonstrates that the
T. reesei cellulase and the termite hindgut homogenates presented similar enzymatic selectivity at a temperature of 27 °C and a pH value of 6.5, although the comparison should be evaluated carefully as enzyme powder and a complex microbiota system were compared. However, these results opened several perspectives to study the parameters influencing the enzymatic selectivity, to evaluate predominant activities in termites from comparison with isolated symbiont profiles, as well as to evaluate endoglucanase activity and potential enzymes for cellodextrins production in cellulolytic systems.
2.2. Oligosaccharides Produced from Crystalline Cellulose
The analytical method was evaluated with standard compounds available for glucose and different cellodextrins: cellobiose, cellotriose and cellotetraose (Sigma-Aldrich). Glucose and cellobiose were not detected below mM levels, due to matrix interferences and limited ionization efficiency. The instrumental limits of detection (iLODs) for cellotriose and cellotetraose were estimated to be 3.5 µM and 1 µM, respectively, based on a signal-to-noise ratio of 3. Analysis of a mixture of cellotriose and cellotetraose at similar levels demonstrated the higher ionization efficiency for cellotetraose, with a signal 3 times higher than cellotriose. These results were important for the analysis of enzymatic profiles, to properly evaluate higher intensity obtained for some DP oligomers.
Avicel and α-cellulose substrates were both tested to compare the influence of the crystallinity index on the hydrolysis reaction. A previous study reported crystallinity indexes for the commercial substrates used in this project, with values 15 to 30% lower for α-cellulose, depending on the method used [
26]. Acid hydrolysis as well as enzymatic hydrolysis with the
T. reesei cellulase were analyzed to validate the method. Fewer cellodextrins (DPs from three to six) were detected using the Avicel substrate in both acid and
T. reesei cellulase profiles, demonstrating that the crystalline structure reduced hydrolysis reaction. Cellodextrins with DPs from three to 10 (with one sodium adduct) were significantly detected (S/N > 10) in acid or
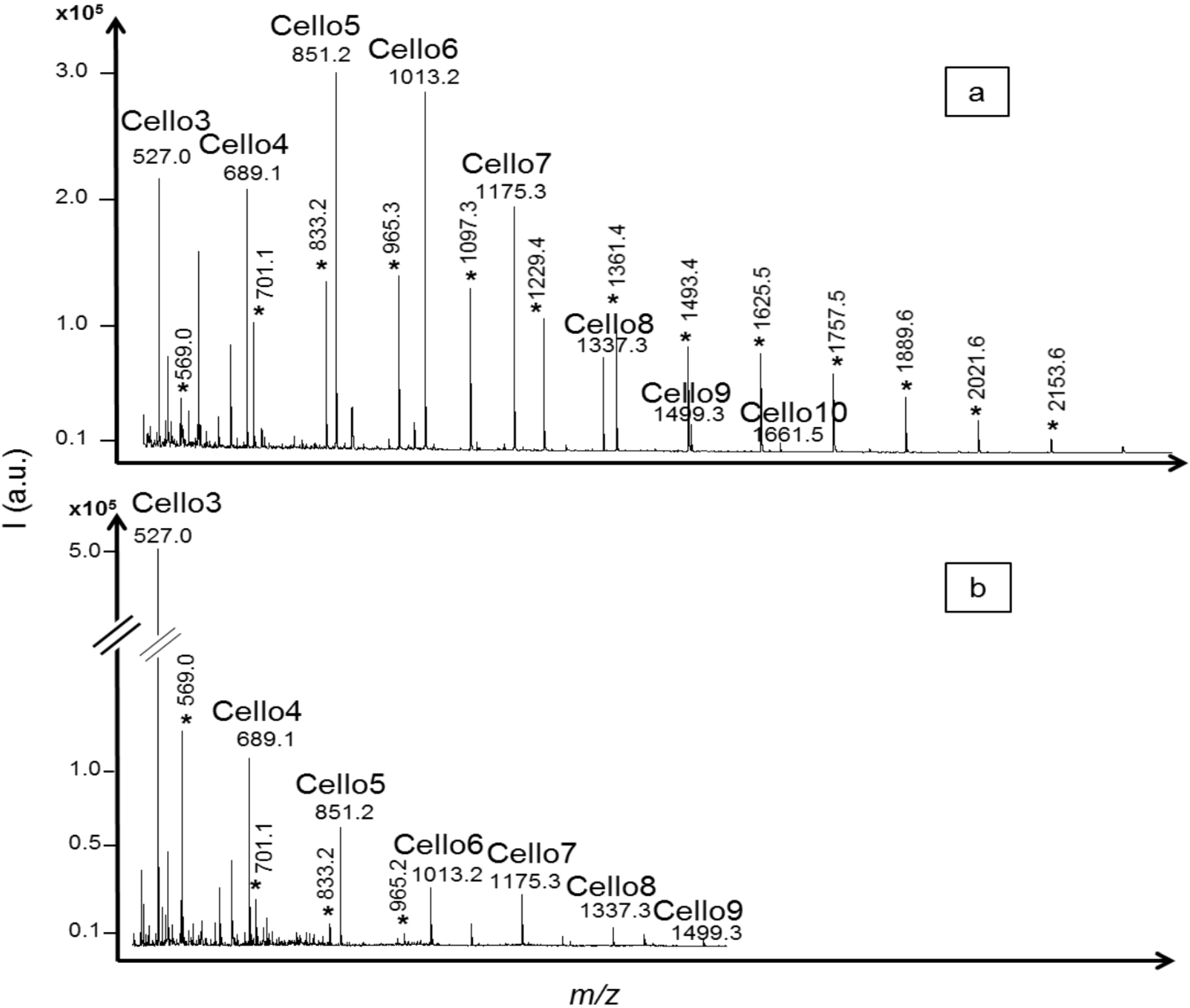
T. reesei cellulase profiles using α-cellulose substrate (
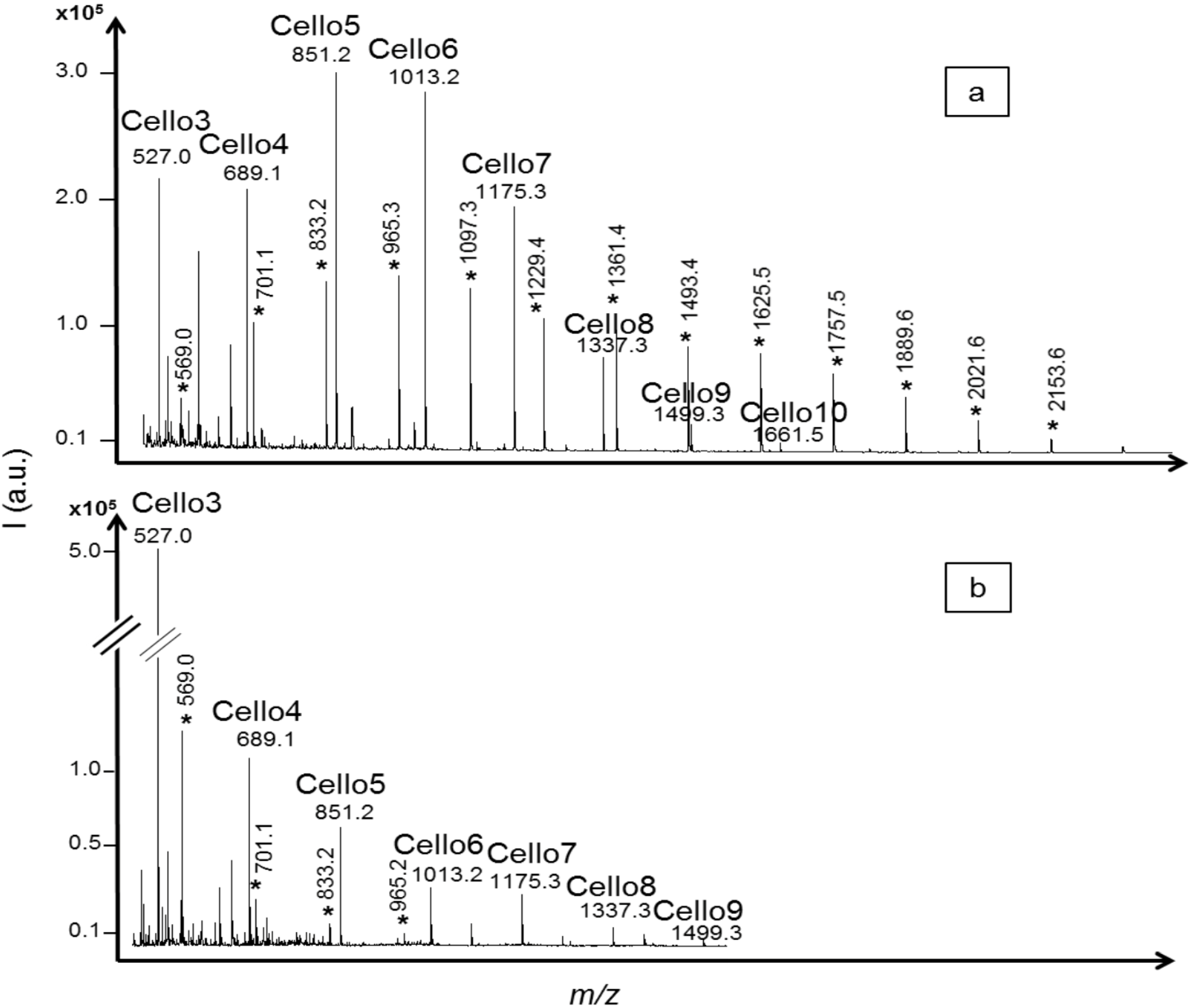
Figure 3). In the acid hydrolysis profile, the intensities of cellodextrins with DPs from three to seven were similar while the intensity of cellodextrins with DP higher than eight decreased, reflecting the discrimination against larger oligomers due to lower solubility. In the
T. reesei cellulase profile, the highest signal intensity was obtained for cellotriose, around five times higher than cellotetraose, despite lower ionization efficiency demonstrated with standards. Cellotriose has already been reported as an abundant and important intermediate product in enzymatic hydrolysis mechanisms [
27]. It is one of the most soluble cellodextrins [
28] and its higher accumulation level, compared to cellotetraose, could be explained by its number of glucose units and potentially a lack of completely filling the number of active sites of endoglucanases for further cleavage [
29]. A similar hydrolysis profile was obtained at a temperature of 27 °C and a pH value of 6.5. These conditions were characterized as not optimal for exoglucanase activity according to results obtained with CMC substrate. The similar profiles suggested that the results could be related mainly to endoglucanase activity.
Hydrolysis products with
m/z corresponding to oligosaccharides of pentose units were also detected in the α-cellulose hydrolysis profiles. Extracted α-cellulose was reported to contain a percentage of xylan [
30,
31]. Xylo-oligosaccharides (XOS) with similar intensities were detected in the acid hydrolysis profile (
Figure 3a, marked with *), from
m/z = 569.0 to 2153.6, corresponding to four to 16 xylose units with one sodium adduct. XOS with DPs from four to seven were detected in the
T. reesei cellulase profile, indicated as
m/z = 569.0 to 965.2 (
Figure 3b, marked with *). This demonstrated the advantages of the method in differentiating the products formed, in comparison with non-specific analysis like reducing ends analysis that would have normalized all the compounds as “glucose production”.
Figure 3.
MALDI mass spectra of cellodextrins produced from α-cellulose hydrolysis by (a) trifluoroacetic acid (TFA) 2M; and (b) a commercial cellulase from Trichoderma reesei. Degradation products of interfering xylan included in the α-cellulose substrate were also detected and identified as xylo-oligosaccharides based on their m/z values (from 569.0 * to 2153.6 * corresponding to four to 16 xylose units including one sodium adduct).
Figure 3.
MALDI mass spectra of cellodextrins produced from α-cellulose hydrolysis by (a) trifluoroacetic acid (TFA) 2M; and (b) a commercial cellulase from Trichoderma reesei. Degradation products of interfering xylan included in the α-cellulose substrate were also detected and identified as xylo-oligosaccharides based on their m/z values (from 569.0 * to 2153.6 * corresponding to four to 16 xylose units including one sodium adduct).
Hydrolyses of crystalline cellulose substrates were tested with the termite hindgut homogenates. Although hydrolysis profiles were obtained with CMC, no cellodextrins were detected by MALDI-TOF MS, even if the partially amorphous alpha-cellulose substrate was used or incubated with freshly prepared hindgut homogenates. The results obtained with the
T. reesei cellulase (
Figure 3b) clearly demonstrated the possibility of specifically evaluating endo-1,4-β-glucanase activity against crystalline cellulose with the method. It is possible that the endoglucanase activity from the hindgut homogenates was combined with higher activity of cellodextrinases and β-1,4-glucosidases, resulting in a limited accumulation of cellodextrins. However, no intermediate compounds were detected when experiments were performed with 1, 2, 4 or 6 h of incubation time with substrates. Levels were probably below detection limits. The cellulolytic system in lower termites like
R. santonensis is described with endogenous secretion of endo-1,4-β-glucanases and β-glucosidases in the salivary glands [
4]. These enzymes are therefore secreted physically earlier in the digestive tube, and they probably act primarily against the amorphous cellulose of the wood particles ingested by termites. However, amorphous cellulose represents a small part of the total amount of cellulose in wood and further digestion of the abundant crystalline cellulose is achieved in the symbiontic section of the gut, essentially by protistan cellobiohydrolases. In this study, the results suggested no significant endo-1,4-β-glucanase activity against crystalline cellulose in the symbiotic microbiota gut part of the termite
R. santonensis. This was demonstrated with crystalline or partially amorphous non derivative cellulose substrates, more representative of naturally occurring cellulose than the conventional CMC substrate. The discovery of a mechanism of depolymerization of crystalline cellulose through endoglucanase activity would have been be interesting to improve lignocellulose valorization with cellodextrins production. More studies on cellulolytic organisms should be performed in that direction, according to the abundance of crystalline cellulose in lignocellulose and the need to improve enzymatic production of cellodextrins.
2.3. Oligosaccharides Produced from Xylan
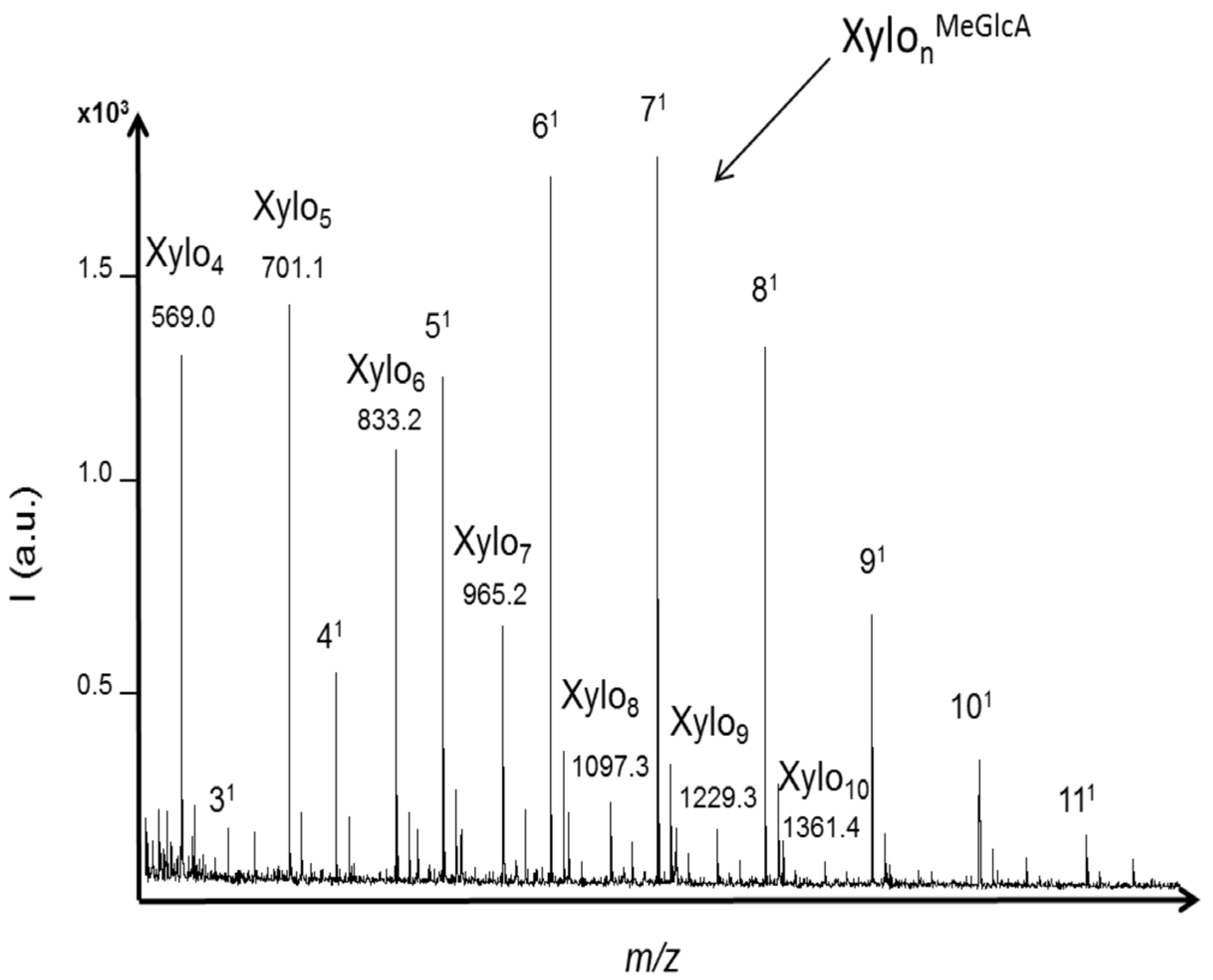
Xylanase activity was evaluated using xylan substrate from birchwood. In hardwood, the principal hemicellulose is a backbone of xylose residues with 4-
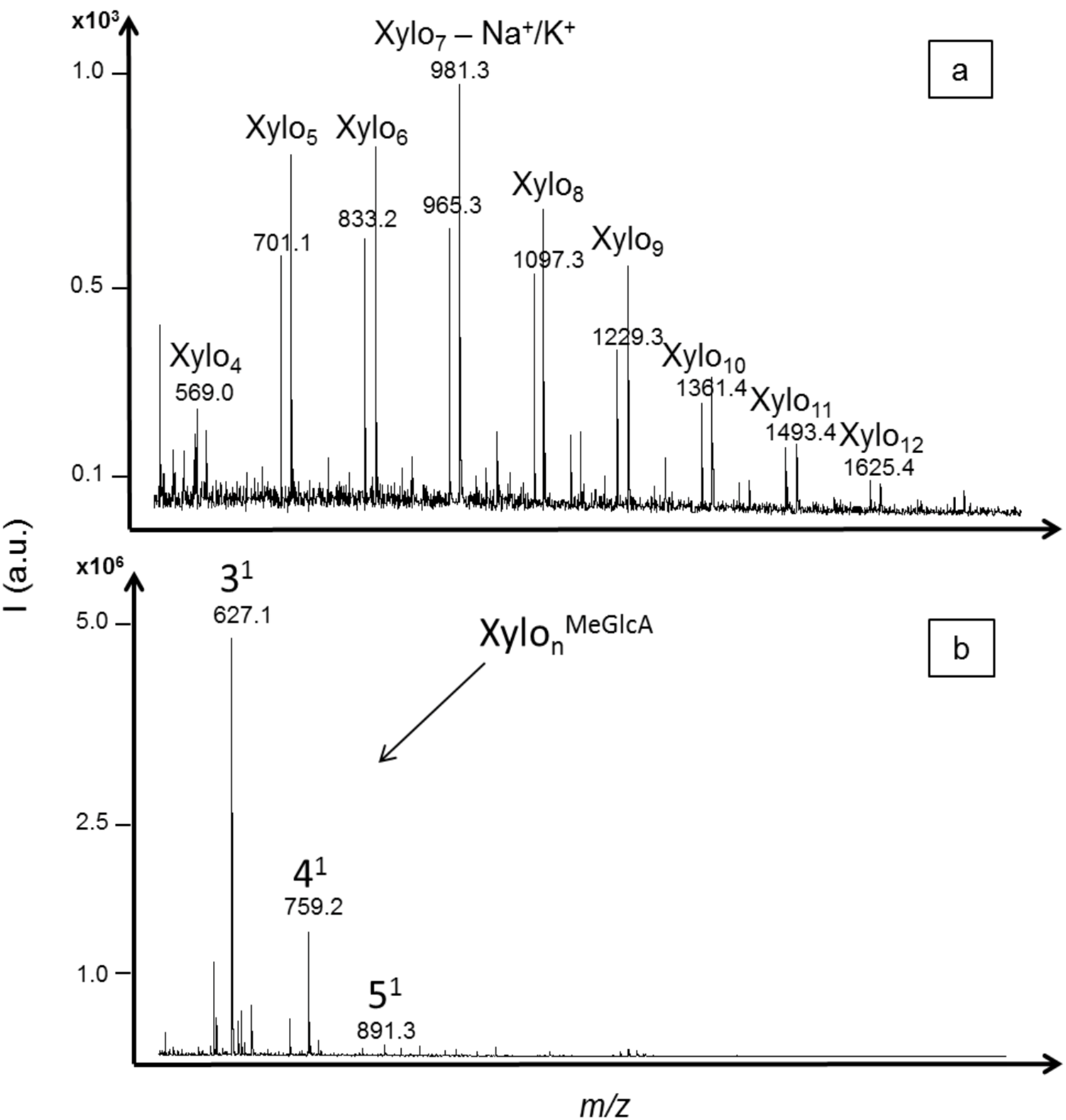
O-methylglucuronic acid substituent attached at various positions along the polysaccharide chain. Only neutral xylo-oligosaccharides with DPs ranging from four to twelve (with one sodium or one potassium adduct) were significantly detected (S/N > 10) with the acid hydrolysis (
Figure 4a). These results showed that methylglucuronic side chains were hydrolyzed with acid treatment. Similar intensities were obtained for XOS with DPs from five to nine while the intensities of XOS with DP higher than 10 decreased. Also, a lower intensity was obtained for xylotetraose.
The enzymatic hydrolysis analysis was tested with an endo-1,4-beta-xylanase of
Trichoderma longibrachiatum. Only acidic XOS were detected, with
m/z values corresponding to oligomers of three to five xylose units carrying one methylglucuronic acid as side chain (including one sodium adduct) (
Figure 4b). The aldotetrauronic acid (4-
O-methylglucuronic acid linked to xylotriose) presented the highest intensity, five times higher than the aldopentauronic acid (4-
O-methylglucuronic acid linked to xylotetraose). In this enzymatic hydrolysis profile, neutral XOS were not detected, probably because they were further hydrolyzed by the endoxylanase activity or by other additional enzymatic activities also reported for this commercial enzyme. The mode of action of endoxylanases belonging to the glycosyl hydrolase families (GHFs) 10 and 11, the two main families of endoxylanases, were characterized on acidic XOS [
32]. It was demonstrated that neither aldotetrauronic acid nor aldopentauronic acid were substrates for endoxylanases of these families. The higher intensity obtained for the aldotetrauronic acid compound in the enzymatic profile supported the hypothesis of additional xylanase activity like β-xylosidase [
33].
Figure 4.
MALDI mass spectra of xylo-oligosaccharides (XOS) produced from xylan hydrolysis by (a) trifluoroacetic acid (TFA) 2 M; and (b) a commercial endo-1,4-xylanase from Trichoderma longibrachiatum. Neutral XOS were identified from acid hydrolysis based on their m/z values including one sodium or one potassium adduct. Acidic XOS corresponding to xylose units branched with one methylglucuronic acid were identified from enzymatic hydrolysis based on their m/z values including one sodium adduct.
Figure 4.
MALDI mass spectra of xylo-oligosaccharides (XOS) produced from xylan hydrolysis by (a) trifluoroacetic acid (TFA) 2 M; and (b) a commercial endo-1,4-xylanase from Trichoderma longibrachiatum. Neutral XOS were identified from acid hydrolysis based on their m/z values including one sodium or one potassium adduct. Acidic XOS corresponding to xylose units branched with one methylglucuronic acid were identified from enzymatic hydrolysis based on their m/z values including one sodium adduct.
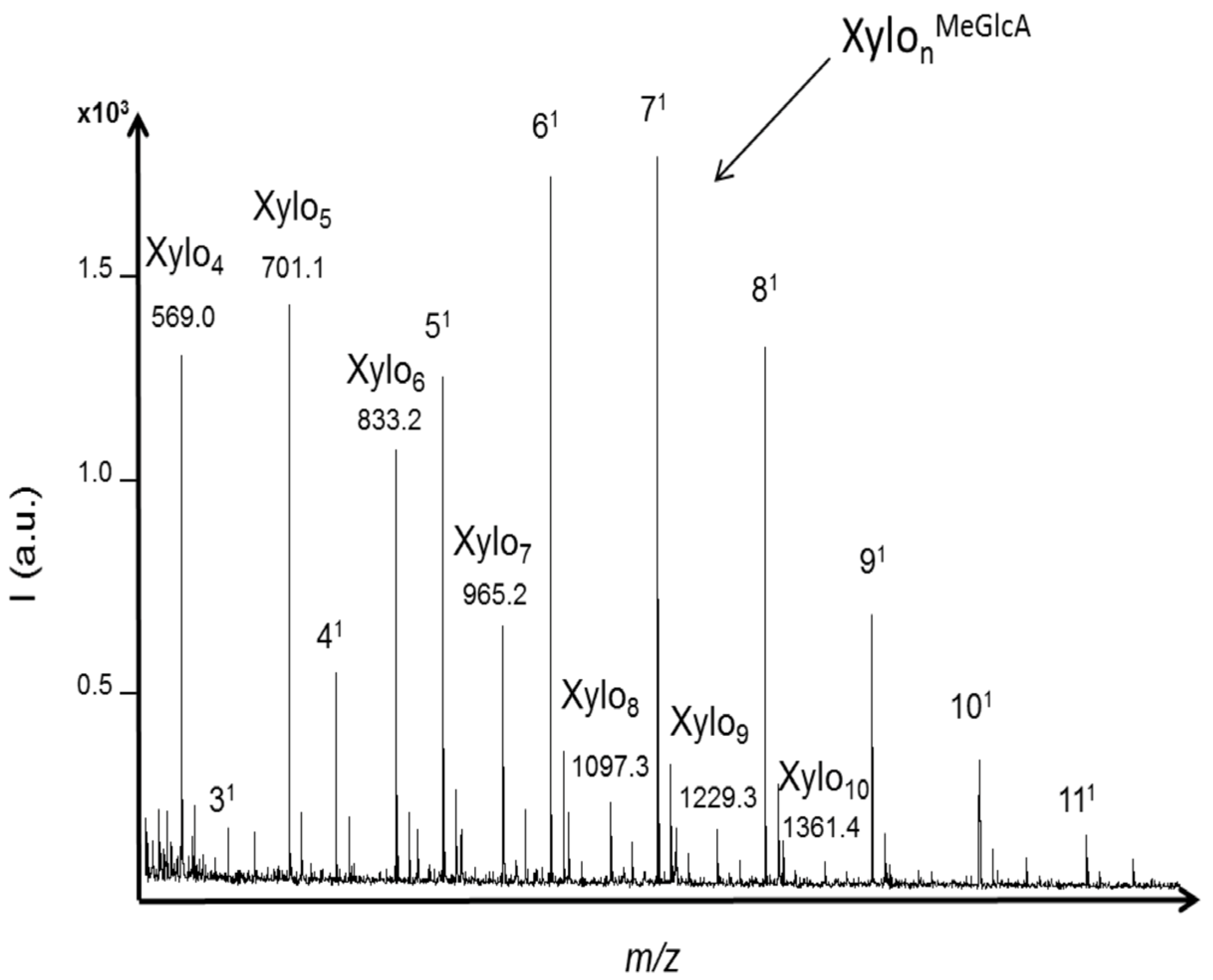
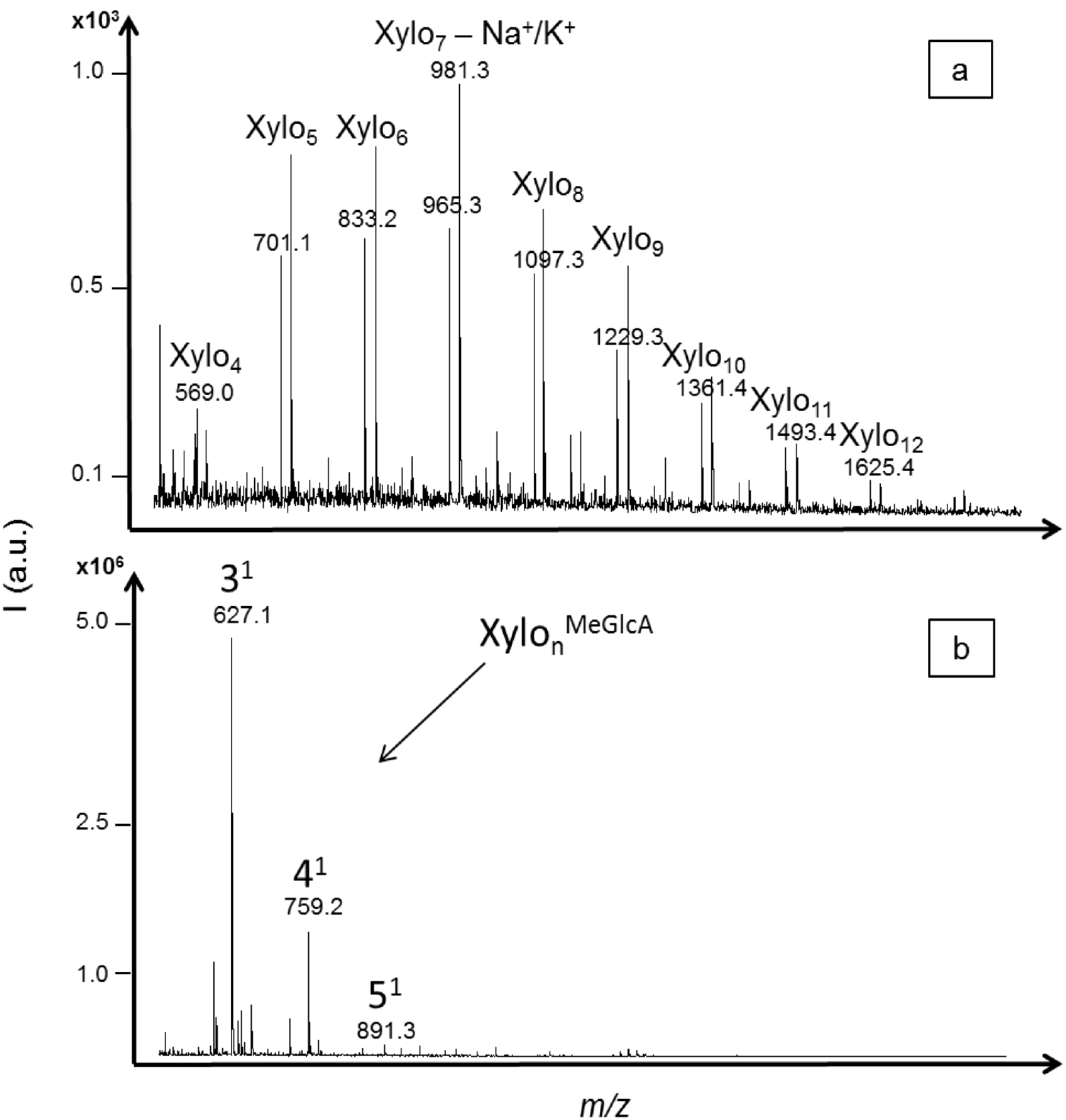
The enzymatic activity against xylan was evaluated with the termite hindgut homogenates. Both neutral and acidic XOS were detected in the profile (
Figure 5). XOS with DPs ranging from four to 10 (with one sodium adduct) were significantly detected (S/N > 10) and the highest intensity was obtained for XOS with DPs from four to six. Acidic oligomers of three to 11 xylose units carrying one methylglucuronic acid as side chain were detected within the
m/z range from 627.1 to 1683.5 (including one sodium adduct). The highest intensity was obtained for the acidic XOS with DPs from five to eight. The profile that was obtained was in accordance with the distribution of 4-
O-methylglucuronic acid in xylan from birchwood that have been characterized as heterogenic and every 10 to 15 xylose residues on average [
34,
35]. The hydrolysis was likely achieved preferentially in regions with no substituted xylose residues, as 4-
O-methylglucuronic acid substituents limited the action of endoxylanases. Then, further hydrolysis was probably achieved between glycosidic linkages closed to substituents, producing both neutral and acidic XOS. It was reported that endoxylanases of GHF-10 are able to cleave linkages in the xylan backbone closer to the substituents. Also, in the case of glucuronoxylan, the shortest acidic fragment released by GHF-10 endoxylanases was aldotetrauronic acid while GHF-11 released aldopentauronic acid. The detection of aldotetrauronic acid in the termite profile suggested that the result of GHF-10 endoxylanase activity could be detected in the profile, although GHF-10 xylanases were reported as much less abundant than GHF-11 in termites from the same family of
R. santonensis,
i.e. the
Rhinotermitidae [
4].
Figure 5.
MALDI mass spectra of xylo-oligosaccharides (XOS) produced from xylan hydrolysis by termite hindgut homogenates. Both neutral and acidic XOS were identified based on their m/z values including one sodium adduct. Acidic XOS from three to eleven xylose units carrying one methylglucuronic acid as side chain were detected within the m/z range from 627.1 to 1683.5.
Figure 5.
MALDI mass spectra of xylo-oligosaccharides (XOS) produced from xylan hydrolysis by termite hindgut homogenates. Both neutral and acidic XOS were identified based on their m/z values including one sodium adduct. Acidic XOS from three to eleven xylose units carrying one methylglucuronic acid as side chain were detected within the m/z range from 627.1 to 1683.5.
Most of the studies on xylanases in termites available to date are based on molecular genetic analysis or xylanolytic activity of isolated and purified enzymes evaluated with reducing ends analysis. Investigations with isolated xylanolytic bacteria and yeasts were also carried out [
36]. It was interesting to evaluate the xylanase activity of a termite gut homogenate as the evaluation of purified enzymes or cultures could largely differ from the activities under natural conditions in the gut [
2]. Moreover, the analysis of intermediate hydrolysis products is important in the case of xylan degradation, as xylan is the main source of oligosaccharides production during lignocellulosic biomass pretreatment. In our study, the MALDI-TOF MS method used was quite sensitive to allow the evaluation of XOS produced from xylan substrate by the equivalent of one termite hindgut. The results demonstrated the mechanism of depolymerization of xylan through endoxylanase, with production of intermediate XOS with DP up to 10.
Xylanolytic activity might play an important role in the efficient lignocellulose degradation achieved by the symbiont-termite association. It can probably be seen as a complementary activity to the mechanical pre-treatment of the wood achieved by the termite host, allowing the access to crystalline cellulose. More studies are needed on the addition of xylanases to cellulase mixtures to enhance enzyme accessibility and improve sugar recovery [
14]. Inhibition mechanisms also need to be further investigated as well as the XOS responsible as different inhibitory effects on cellobiohydrolases were observed. It is possible that cellobiohydrolases produced by termites or their symbiotic microbiota are less inhibited by XOS or not inhibited by XOS with higher DP. In such a case, a better control of the DP of the produced oligosaccharides would be required, according to specific endoxylanases, amounts and conditions.

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}